
bonilla1993
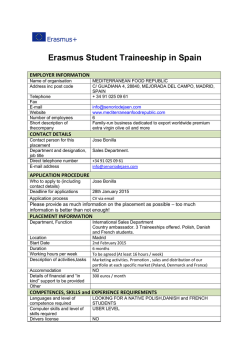
JOURNAL OF GEOPHYSICAL RESEARCH, VOL. 98, NO. C2, PAGES 2245-2257, FEBRUARY 15, 1993 Seasonal Distribution of Nutrients and Primary Productivity on the Eastern Continental Shelf of Venezuela as Influenced by the Orinoco River JAIME BONILLA,1 WILLIAM SENIOR,1 JOHN BUGDEN,2 OLIVER ZAFIRIOU,3 AND RONALD JONES2 Nitrogenous nutrients, dissolved silicate, and salinity were measured in surface waters and shallow hydrocastsalongsimilar cruisetracks duringthe spring(dry season)and fall (wet season)of 1988.Both cruises transected the eastern Caribbean, transited the Gulf of Paria, ran parallel to the Orinoco Delta and into the main channel of the Orinoco River. Trends in primary productivity were also measured by daily carbon 14 incubations. In both seasons,samplescovered the range from highly oligotrophic and transparentto highly productive and rich in biogenicand abiogenicparticulate matter. Most of the I•rinnr, n n,,tflnw •ppo•re tn t,,m 1%/tn l•I•u•7•nrl r•molne in eho!!n• ,x;o•,•.o ngle ,1....... 1• •nA surroundingTrinidad, permitting benthic regenerationof river-borne nutrients. However, the role of the Orinoco and associated low-salinity coastal waters in fertilizing large areas of the eastern Caribbean basin, as suggestedby satellite imagery, can be approximated crudely from the nutrient compositionat Boca de Dragon, which is representativeof the nutrient statusof these waters as they flow into deeper Caribbean waters. Additional nutrients may be supplied to the area primarily from Amazon-derived water entering the Caribbean Basin further north, with some coastal upwelling along the continental shelf in the dry season. of 33.95x 103 m3 s-1 [Salazar,1989],second onlyto thatof INTRODUCTION Coastal oceanic water masses,especially those located in the tropics, are among the most fertile and productive. This biological and organic richnessmay result from high loads of nitrogen, phosphorusand silicon suppliedby hydrodynamic processes resulting from continental drainage, or from coastal upwelling processesduring seasonsof winds favoring this process. Within the context of coastal bioecosys- the Amazon. The flow regime is characterized by precipitation throughout the year, with maximum flow in August and minimum flow in March. The estuaries formed in the delta are known as Boca Ajies and Boca Bagre in the Gulf of Paria, and Boca Grande or Boca de Navfos, where the navigation channel is located (Figure 1). Knowledge of the estuarine-deltaic system of the Orinoco and the Gulf of Paria dates from 1954, when Van rems, estuarine-deltaic zones are notable as transition enviAndel and Postma describedthe hydrographic, climatologironments between the ocean and the continent. The Orinoco cal and sedimentary conditions of the area [Morelock, 1972; River systemin Venezuela provides one of the largest-scale Eisrna et al., 1978]. The formation of muddy plains in the examples of these interactions. It has a major influence on deltaic coast is due to depositionof sedimentstransported to the hydrographic and biological conditions of the estuarine the northwest by the Guyana current. Butenko et al. [1981] system of the Guff of Paria and the Caribbean Sea. reported the formation of a clay wedge in the delta area up to The Orinoco River begins near Brazil, at the junction of about 80 km into the adjoining Atlantic, with additional Sierra Pfirima and Sierra Unturan, and flows over an area of organic clays and inorganic input from the Amazon River. 2560km2, of which1670km2 are navigable. It irrigatesa This was confirmed by Milliman et al. [1982], who found floodplainof 981,000km2 in Venezuela,Colombia,and montmorillonite characteristic of the clay sediments of the Brazil, is the only South American river with a true delta, Amazon River in the Orinoco delta, also transported by the and has an ocean front of 300 km. Its principal affluent is the Guyana current. Herrera and Masciangioli [1984] deterCaroni River, with a brown-ochre color, which is mixed with minedthat the predominantcirculationin the coastalarea of the waters of the Orinoco, which are yellow. The Orinoco the delta consistsof surface currents generatedby the trade flow is turbulent and contains a large amount of suspended winds moving to the west and northwest. The hydrographic clay. These Orinoco-Caroni waters are rich in organic and and chemical conditions of the sediments in the Gulf of Paria inorganicmatter which is transportedto the Atlantic Ocean were studied by Benitez and Okuda [1976], Bonilla [1977], and the Caribbean Sea through the estuarine-deltaicsystem. and Bonilla and Lin [1979]. The geochemicalcharacteristics The Orinoco River contributesa large amount of suspended of the sedimentsin parts of the river and delta were studied sediments (86.3x 106tonsyr-•), withanaverage waterflow by Bonilla et al. [1985]. Miiller-Karger and Varela [1988] 1Instituto Oceanogrfifico deVenezuela, Universidad de Oriente, Cumanfi. determined that the nutrient influence of the Orinoco River on the Caribbean Sea reaches Puerto Rico, based on the 2Department ofBiological Sciences andDrinking WaterResearch concentrationof pigmentsduring the high flow seasonfor the Center, Florida International University, Miami. Orinoco (August-October). Salazar [1989] studied the hy- 3WoodsHole Oceanographic Institution,WoodsHole, Massa- drogeochemicalconditionsof the estuarine-deltaicregion of chusetts. the Orinoco, from the island of Curiapo inside the fiver to Copyright 1993 by the American Geophysical Union. 180 km into the Atlantic Ocean. In these tropical bioecosystemsthere are two seasons:a dry seasonfrom December to April, and a rainy seasonfrom Paper number 92JC02761. 0148-0227/93/92JC-02761 $05.00 2245 2246 BONILLA ET AL ' NUTRIENTS AND PRIMARY PRODUCTIVITYOF THE ORINOCO 64.0 $ :•o ! I .... (50 ø •1 I ! -- 15" CARIBBEAN S E A -- 13 ø ATLANTIC M OCEAN eN ,..:•. =:eQ ß0 '" ' .,...,.•" ';.• i""?';":'" • P5 GulfofPari. ,P2 p• '•:. • -- • ,I R1 9 o -- -• 64 ø VENE Z UELA I , '•'•' '•1•;'" ".• ."•1• R5,' OR •NOCO I R. 62 ø ß I ':.• , , I ...."• G0• 58 ø Fig. 1. Locationof the sampling sitesfromColombus Iselincruises CI-8805andCI-8816,springandfall 1988(solid line, E-W transect;dashedline, NE-SW transect). May to November. Evaporation is highestin February (129 mm) and lowest in November (76 mm), while precipitationis highestin August (127 mm) and lowest in April (19 mm). Thus the timing of the two cruises reported here nearly coincided with the two extremes. Westerly winds are prev- the hydrochemicalmechanismsof the delta, conservative and nonconservativeprocessesresultingfrom the runoff of the Orinoco during high and low flow are considered. METHODS alent, with an easterncomponentduring most of the year. A semi-diurnaltide regime dominatesin the estuarine-deltaic Source of Samples region [Salazar, 1989]. Water sampleswere collected in the estuarine-deltaic Here we summarize the saline stratification, nutrient distributions,and primary productivitymeasurements from the systemof the OrinocoRiver, Gulf of Paria, and adjacent spring (April 1988) and fall (September 1988) cruises. We areas(Figure1) duringresearchcruisesCI-8805in the spring focus on longitudinaltransectsNE-SW and E-W from the (April 1988)and fall CI-8816 (September1988)on boardthe mouth of the Orinoco. This paper focuseson the near-shore R/V ColumbusIselin. Seawater sampleswere taken using a data (all data S of • 11øN), as studiesin that area were the rosettesampler(GeneralOceanics,Miami, Florida)consistmost intensive and the rivefine impact is strongest. For ing of twelve 5-L teflon-linedpolyvinyl chloride Niskin effects in the Caribbean N of 1IøN, see the accompanying bottles. The rosette was fitted with a Neil Brown conductiv- paper [Zika et al., this issue].In order to betterunderstand ity, temperature,anddepth(CTD) probe.Additionalsurface BONILLAET AL.' NUTRIENTS ANDPRIMARY PRODUCTIVITY OFTHEORINOCO 2247 (2 m) samples werecollectedfromthethrough-the-hull teflon limitedprimarilyto a shallowsurfacelayer and are hardly detectable evenat 50 m. Thisfactorcanbe seenclearlyby comparing salt,silica,andnitrateseasonaldifferences along Nutrient Analyses bothtransects'NE-SW (Figure2, spring;Figure3, fall) and Water sampleswere collectedin 60-mL Nalgenebottles E-W (Figure 4, spring;Figure 5, fall). The tendencyto and refrigerateduntil analyzed.Sampleswere analyzed upwelling in the spring and its relaxation in the fall are within24 hoursaftercollection.Turbidsamples werefiltered clearlyindicatedin the NE-SW (onshore-offshore) sections using0.45 tampore-sizeAcrodiscdisposablefilter assem- by the slopingand horizontal(respectively)isolinesfor S = blies(GelmanSciences,Ann Arbor, Michigan). 36 ppt, and silicate or nitrate 1 taM. The E-W transect Nutrientdeterminations (NO•-, NO•-, Si(OH)4)wereper- comparisonmost clearly showsthe much larger extent of formedon an AlpkemRFA-300nutrientanalyzer(Alpkem freshwaterinfluence'the springtop-bottomcontrastis •31Corp., Clackamas, Oregon). All nutrient determinations 36.5 ppt, while in the fall the rangeis •13-36.6 ppt. were measuredagainstinternalstandardsaccordingto the The slopingisolinesindicatethe dry seasonupwellingof RFA methodologyhandbook(Alpkem). intermediate water from the southeastern Atlantic in the bow pumping system. •JIGLLM$1111 GL.I.)•/•,,,,CL %JICllI•.I•5. ¾¾ •tL•l Ol bi:tlllllty ..•2.2 pp[ at Primary Productivity Measurements Surfacewater (•0-2 m) sampleswere collectedin BOD (R1) ascendsto the surfacelayers (2-10 m), pushingthe freshwater massintothefiver (Figure2) fromthe surface(R3) with changes in salinity,delineating thisareaof the estuary from the salt wedgetowardsthe bottomat R5, where the bottlesto which0.95mL of NaH•4CO3(2.5 taCimL-• 42.37nmolmL-• Amersham Corp, Arlington Heights, wedge'sapexis formed.The mixingzonewasbetweenR5 and , Illinois) was added. The sampleswere irradiated in full R6 nearthe platformat BocaGrande.At a depthof 90 to 140 sunlightin a flowingseawaterbathfor 3 hours(darkcontrols were covered with aluminum foil). After irradiation the sampleswerefilteredthroughGF/F glassfiberfilters(WhatmanInternationalLtd., Maidstone,England),rinsedwith 50 m there was a cold, deep-watermass from the southeastern Atlanticwhichwashigherin salinity(36.8-37.15ppt)between R1 and R2, rich in nitrogenouscompounds(nitrite, 0.18-0.16 taM;nitrate,0.47-6.2taM)andlow in silicate(0.12-0.35taM). mL of 1 N HC1 and storedfor furtheranalysis.Integrated Thus, duringthe dry season,when the Orinoco has its lowest photosyntheticallyactive radiation (PAR) was measured flow,itsinfluence islimitedto a narrowfringestretching along with a LI-COR (model 1800)calibratedspectroradiometer. Radioactivity was determined with a Beckman LS 3801 the estuarine-deltaic area (Figures2 and 3). liquid scintillationcounter(BeckmanInstruments,Fullerton, California).Primaryproductivityvalueswere calculated Nutrients and Primary Productivity as tagCfixedper m3 h- •. The verticalnutrientdistributions are alsoshownin Figures2-5, while the surfacevaluesandprimaryproductivity RESULTS dataare givenin Figures6 and7. Table 1 presentsa highly Spring and Fall Salinity Distributions condensed summary of the data for both seasons in the vicinity of the Orinoco River. Giventhe restfictionsimposedin the springby navigation The spring-fallsalinity distributions(Figures 2-5) are dramaticallydifferent.In springtherewasa sharpfreshwa- requirementsand the strongtendencyof the fiver water to sheet ter-seawater interface within the navigation channel and spreadover the saltwater in a thin difficult-to-sample in both seasons, these data are obviously too sparse and upstreamof Pt. Bafima,sothatthe NE-SW transect(Figure 2) neverencountered purefiver water(Smin ----8 ppt at R5 at unrepresentativeof the fiver-delta-coastalsystemas a whole ebb tide). The E-W transect(Figure4), as closeinshoreas to permit a detailedcharacterizationof the regionin either comparison. The purpose feasiblenavigationally, alsoencountered onlyrelativelysa- season,or a detailedinterseasonal line waters (S > 30 ppt) becausethe saltwater-freshwater of gatheringthese data was rather to provide ancillary gradientwas centeredcloseinshorein the nonnavigable informationfor the more detailedprocess-ofientedstudies watersof the Orinoco'sextensivedeltaicsystemanddistrib- reportedin other papersin this Special Section. utaries in spring. In fall, both sections were within the core of the near-field freshwater-saltwater mixingzone (Figures3, 5). The E-W transectand the Gulf of Paria was roughly50% saltwaterat thattime,whiletheNE-SW transectencountered S = 0 ppt Nevertheless, a few issues of general interest to the biogeochemistry of this fascinatingbut relatively littlestudiedregioncanbe addressed with thisdataset. They are discussed below. Some of these same issues are also ad- dressed fromdifferentperspectives in someof the accompanyingpapers(e.g., the Si budgetand Ra-Si relationships [Moore and Todd, this issue];primaryproductivity,nutrigivenby Rodriguez[1975]for April, but in September the ents,and specificpigmentdistributions[Bidigareet al., this E-W transectandGulf of Pariawaterswere alreadyalmost issue]). twiceassalineasthosereportedfor August,suggesting that DISCUSSION the peakflow was alreadywell pastandthe gradualadvection and mixing of this large input with seawaterwere underway.Wheretherewas sufficient waterfor navigation Silicateand (Nitrate Plus Nitrite) in Springand Fall by the R/V Iselin, the delta region of the Orinoco River Coastalnitrogencyclingis intenseandcomplex[Carpenexhibiteda typical salt wedgecirculationwhich governs ter and Capone, 1983].The availabledata provide strong hydrographicand chemicalconditionsin deeperwaters. qualitativeconfirmationof the expectedintensebiological Thus the springversus fall effects of the rivefine inflow are cyclingin this region, as discussed previously[D'Armas, water from surface to bottom at R5. The surface distribution of salt (not shown) on these two cruisesthus resembledthat 2248 BONILLA ETAL.' NUTRIENTS ANDPRIMARY PRODUCTIVITY OFTHEORINOCO R1 $w NE $w NE n2 3O 4O 5O 6O 8O •loo Salinity IO0 (%ø) 120 120 Nitrate ß (•LM) 140 140 160 160 180 180 2OO 2O0 3OO $W NE $W NE o lO 2O 20 3o >0.05 4o 5o 0.1 Nitrite 120 140 160 180 (I•M) Silicate 12o (•LM) 14o 16o 18o NE-SW Spring Fig. 2. Depthprofileof salinity, nitrite,nitrate,andsilicate ona NE-SWtransect, spring 1988. BONILLAET AL ßNUTRIENTSANDPRIMARYPRODUCTIVITY OF THE ORINOCO NE W NE ß 2O 2249 20 3O E 1 2 Nitrate o) 100 lOO --6 , 120 • • --:36.8 -.... ß.._• • 120 ß 140 ,. 140 160 ß ß 160 180 o ß 180 2O0 NE SW R1 o , 4/ lO 20 •2 \ E A 50 5o -I /d •. so C] 8o Silicate Nitrite 100 lOO (IzM) 12o 120 140 140 160 ß 180 ß 200 ß NE-SW Fall Fig. 3. Depthprofileof salinity,nitrite, nitrate,and silicateon a NE-SW transect,fall 1988. 1986; Salazar, 1989] for November conditions in the water of Paria have been describedby Bonilla and co-workers, as column of the delta region. Sedimentaryconditionsthere have been discussedby Milliman et al. [1982], Eisma et al. [1978], and Kennicut et al. [1987] and conditionsin the Gulf referenced in the introduction. In addition to salt, other semiconservative tracers are usefulin estuarinestudies,mostclassicallydissolvedsilicate 2250 BONILLA ETAL.:NUTRIENTS ANDPRIMARY PRODUCTIVITY OFTHEORINOCO (us)qtdea (us)qtdoG BONILLA ETAL.' NUTRIENTS ANDPRIMARY PRODUCTIVITY OFTHEORINOCO (w) qld•a I I (w) qld•a I (LU)•dea (m) qldea 2251 2252 BONILLAET AL.' NUTRIENTS ANDPRIMARYPRODUCTIVITY OFTHEORINOCO i iß lOB ß te lO' SEA Spring Nitrite (•u) OCEAN . Spring ß-' Silicate ./' Spring / t/,ø' o---- (•U) .•• Primary Productivity (•gc/re') AT•ANTIC • / OCEAN / ,/ / / , / ; / ( ,/ : / / . . / ß ß \ io .. , t 'd • • :f r••, •1 • •.",," 5'0'..' ' • 0ß Fig. 6. Spatial distribution of nitrite,nitrate,silicate, andprimary productivity inthesurface waterduring thespring 1988. Primary productivity values areexpressed as/xgC m-3 h-] . behavioron spatialand [Burtonet al., 1970;Liss, 1976],as exemplifiedrecentlyfor lead to seriouslynonconservative the Mackenzie Shelf Estuary by MacDonald et al. [1989]. temporalscalesthat needto be definedempiricallyin each However, silicatedistributionsmustbe interpretedwith care system[e.g., Edmond et al., 1981]. However, use of Si as a tracer has a history in the greater becausebiologicaluptakeand subsequent sinkingandredissolutionof biogenicopal in the water and on the bottomcan easternCaribbeanbasin with respectto understandingthe BONILLA ET AL.' NUTRIENTSAND PRIMARYPRODUCTIVITYOF THE ORINOCO ii- i1,' - C&l•1111 Fall Nitrate All , , ß Fall /•f Nitrite • (izM) 2253 ' ii I ii ß OCEAII / -'' io . CAlllie ,/' / / /, '"' 4111 I ß Fall Silicate •M) Fall Primary Productivity c/re') IULLOF PAllIA ,\ Fig. 7. Spatialdistribution of nitrite,nitrate,silicate,andprimaryproductivity in the surfacewaterduringthe fall 1988.Primaryproductivity valuesareexpressed as/xgCm-3 h-] . influenceand seasonalvariabilityof the greatriver systems to the southeast(Amazonand Orinoco)versusrain inputs. Froehlichet al. [1978]first usedSi sectionsto suggestthat rivers, not rainfall, were the principalcauseof the seasonal ine inputsto this region.The radiochemistryapproachwas alsoappliedon CI-8805 and 8816;the combineduse of Si and surface fresheningas far away as Puerto Rico. More re- radiochemical data to determine the "estuarine cently, Moore et al. [1986] used Ra-Si relationships to construct quantitative, sensitive tracers of the remote river- end mem- 2254 BONILLA ET AL.: NUTRIENTS AND PRIMARY PRODUCTIVITY OF THE ORINOCO TABLE 1. Nutrient Content and Primary Productivity in Different Offshore Regions of Estuarine-Deltaic Systems of the Orinoco Spring Fall Parameter Region Measured Dragon's Mouth NO/NO•Si(OH) 4 31.0 NO•NO•Si(OH) 4 0.16 0 4.90 PP Serpent's Mouth 14.0 NO•NO•Si(OH) 4 Shelf River (surface transect DM) Ocean Maximum 0.27 0.55 3.21 PP Gulf of Paria Minimum 0.12 0 0.93 PP ...... NO•NO•Si(OH)4 0.20 0 2.05 Average Minimum Maximum 0.45 5.34 4.76 m • m m • • 12.9 12.9 12.9' 0.06 0.26 10.53 1.01 7.03 68.64 0.41 3.25 40.75 0.56 8.12 6.42 31.0 31.0' 0.60 1.00 7.43 0.34 0.41 6.33 14.0 14.0' 26.0 52.8 38.0t 0.42 0.98 6.01 • • • • • m m • • 0.97 9.30 17.92 9.55 3.75 6.83 0.15 0 4.61 35.0 35.0 35.0* NO•NO•Si(OH)4 0.36 0.13 10.84 2.09 4.52 98.26 1.38 3.24 41.46 0.02 0 12.20 PP 50.0 50.0 50.0* 21.6 0.02 0 0 PP 1.0 • • • 0.92 3.37 11.86 PP NO•NO•Si(OH) 4 Average 0.36 22.75 12.00 1.0 0.71 3.89 28.00 2.9 0.12 4.12 1.50 0 0 0 1.0' • 0.46 1.45 9.41 2.9 2.9* 1.01 6.55 149.80 0.30 5.00 110.46 31.2 26.45 0.86 16.48 9.86 0.08 3.58 3.52 • • PP,primaryproductivity (/zgCm-3 h-l), NO•-, NO•-, andSi(OH)4(/aM). *One value only, no average, depth 5 m. ?Average of four values, depth 5 m. SAverage of two values, depth 0 m. gles), often with NO/-/NO/- approachingunity. This com- ber" by this approach is discussedin their paper [Moore and Todd, this issue]. Hence it is of interest to examine its behavior also in the estuarine region which may alter this bination suggeststhat very active nitrification was convert- water before Saunders [1989] report •2.5 it reaches the eastern Caribbean. ing ammoniaand possiblyDON to NO/- + NO/-. Lewis and Figure 8 shows silicate versus nitrate plus nitrite plots for both seasonsalong with the mixing lines expected for spring and fall Orinoco fiver water plus oligotrophic seawater (nutrient-salinity plots cannot be used in spring, when all the low-salinity sampleswere acquired in rapid successionfrom the pumping system and could not be accurately matched with salinities). In spring (solid symbols) the data are from the vicinity of R4-R5, whereas in fall the same [Si] range was covered over much of the E-W transect O Fall (transect) ß Spring(R4-R5) ß [NO2']>21aM ß CH4>0.1gM / (•) Orinoco River Means 7 (8) Spring // // ß / outside of the Gulf of Paria. As noted above, the highest Si values in both seasons closely approximate those expected from the riverine data of Lewis and Saunders [1989]. In contrast, in both seasons NO/- + NO/- clearly cycles dynamically in waters of lower salinity (higher Si). The dominant cycling mechanisms appear to differ seasonally, as described below. Proceeding from high to low Si values in spring, we /xM ammonia and >11 ß / // / •, 6 ßßß ////Slope-0.075 O initially encounter waters with very low NO/- + NO/-/Si compared to that expected from the river-seawater mixing (Figure 8). These samples were from within the turbidity maximum, where photosynthetic uptake cannot be the nitrate sink. Rather, the associated high methane values in some samples (square symbols) strongly suggestthat interactions with anoxic bottom and pore water environments were intense. Loss of N0•- + N0•- by denitrification(and possibly Si injection by redissolution) are likely the domi- nant processesredefiningthe NO/- + N0•-/Si ratios here. At intermediate [Si] the data fall on the high N0•- + N0/-/Si side of the mixing line and the highestvalues are associatedwith unusually high nitrite concentrations(trian- /• IY II//./ 0 // / I/' / . 20 o .o 40 . 60 ß 80 100 120 140 Silicate•uM) Fig. 8. Plot of nitrite and nitrate versus silicate in surface water for spring and fall 1988. BONILLA ET AL.' NUTRIENTS AND PRIMARY PRODUCTIVITY OF THE ORINOCO Surface E-W Fall Transect - 140 2255 Boca Grande (R4-R5) may not typify the myriad other unsampleddistributariesto the NE and enteringthe Gulf of Paria. - 120 ß . [] - 100 80 • - 6O - 40 - 20 I I I ß I I 0 ,•0 Salinity(%o) Gulf of Paria and Inputs to the Caribbean Although some Orinoco and coastally trapped Amazon water enters the Caribbean by way of Galleons PassageN of Trinidad, etllux out of the Boca de Dragon is thought to represent a major component of the estuarine input to the Caribbean, as shown strikingly by satellite photographs [Miiller-Karger et al., 1990]. The gyre circulation in this Gulf [Bonilla, 1977; D'Armas, 1986]tendsto homogenizeits waters, includingthe inputs of the northernmost distributaries discharging into the Gulf itself. Hence it is especially interesting to examine the behavior of Si on this scale. Taking our fivefine end-member values and R1 as a seaward end-member for Atlantic water Fig.9. Plot Ofnitrite plus nitrate versus salinity andsilicate versusentering the Caribbean in the vicinity of the Grenada passalinity from the surface water of E-W transect, fall 1988. sage, conservative mixing of surface waters in spring predicts = 18 tam silicateat P1; <7 tam was present. Thus at low discharge, the residence time of water in the Gulf is apparently long enoughthat substantialbiological removal occurs. DON in the riverincend-member. Displacement of NHf/ In contrast, in fall Si appearsto be conservative along the DON bound to clays by seawater cations in this zone of intense mixing would provide a mechanism to free new coast and in the Gulf of Paria (Figure 10 inset), despite substrate for subsequent oxidation to nitrite by bacteria evidence for localized Si removal (Figure 9). Presumably, [Carpenter and Capone, 1983], while the high turbidity either this Si was regenerated and re-entrained in the flow would minimize light inhibition [Olson, 1981; Vanzella et al., (perhaps in the course of turbulent mixing in the Boca de 1989].Finally, at <20 tam Si this highNO•- + NO•-/Si water Serpente), or the fraction of the total flow thus affected was appears to mix conservatively with oligotrophic seawater, minor. This mixing line thus defines the Gulf of Paria source perhaps with some photosyntheticuptake of N at the very water to the Caribbean (at this time) as having an endlowest [Si]. member value of =120 tam Si representing fiver water Thus these data show that in spring, nonconservative (Orinoco plus coastally trapped Amazon flows) little modified by dilution by rainfall or biological removal. This differs behaviors indicating biogeochemicalcycling and input of N to coastal waters were intense in this very active zone, from Froehlich et al. [1978] estimate of average freshwater generatingboth relatively N-poor and N-rich waters (N/Si input to the eastern Caribbean (for the entire eastern Caribratios =0.1-5 of the riverinc ratio). However, our sampling bean, a significantamount presumably being Amazon water was far too sparseand unrepresentativeto permit estimating entering with the North Equatorial and Antilles currents the relative amounts of these two types of water input to the through more northern passages) had an apparent endcoastal zone over any significant area, and the absence of member compositionof =60 tam Si, correspondingmostly to ammonia and DON data prevents a comprehensive mass three-componentmixing of Amazon water (with nearly the same Si content as Orinoco water, = 130 taM), rainfall, and balance approach. In fall (Figure 8, open symbols)the much larger river flow oligotrophicsurface seawater. pushed high-Si water out well beyond the core of the Figure 10 is an Si-salt plot with our end-membermixing turbidity maximum; waters with stronglyenriched NO•- + line (dashed), that of Froehlich et al. [1978] (heavy solid), NO•-/Si ratio were absent. The most Si-rich waters were also higherin NO•- + NO•- than expectedfrom the data of Lewis and Saunders [1989] but in the expected ratio. The fall data are plotted against salinity in Figure 9, revealing estuarine Si consumption and strong NO•- + NO•- uptake, especially near =7 ppt salinity. This presumably reflects photosyntheticuptake by silicoflagellates(see Edmond et al. [1981] concerningthis same situation in the Amazon plume). At higher salinities, Si is more nearly conservative and N + N is exhausted. These trends suggestremineralization of Si and continued uptake of N + N by nonsiliceousprimary producersat higher salinity. However, in both seasonsthese interpretationsmust be tempered by the realization that the system was probably not at steady state, that significant vertical gradients which may have originated by nonconservativeprocessesare ignored, and that the "riverinc" (near Barrancas, at the head of the deltaic region) endmember data of Lewis and Saunders [1989] or our samplesat and the surface Si data for the entire east. Caribbean cruise track (Figure 1) in fall 1988. Much of the data lie near the line of Froehlich et al. [ 1978], especially if adjusted to assumean end-memberwith 0.5 tam Si at 36.2 ppt rather than the value of 1 at 36.4 they used (light line). However, to the north (up to about 13øN)many points lie in between these two or along our mixing lines. If Si is nearly conserved in the E Caribbean, our Si data suggest a modified view of the threecomponentmixing concept to account for the fact that the Gulf of Paria componentis apparentlylittle diluted by rain, whereas the "offshore Amazonian" inputs supply raindiluted fivefine Si. Thus our Si data more or less directly demonstrate the existence in fall of two freshwater/Si inputs and show that the sourceof high Si water is at least partly associatedwith the Orinoco input through the Gulf of Paria, up to = 13øN. Thesefindings thusreinforce theconclusion from228Ra data by Moore et al. [ 1986]that two discreteroutes of delivery for 2256 BONILLA ET AL.: NUTRIENTS AND PRIMARY PRODUCTIVITY OF THE ORINOCO 14- •• 12 - \\\xxx,'•'/ ß GulfLine ofParia ß Froehlich,-/•'• oeec',, (inset) 10- et al. (1978); Venezuela Basin • - • (67ow) • •, 'x ß •x• ' 6- • 4 '•'• GulfofPa•a -•• return toMiami Track) "Re-adjusted"/• • • I 0• No.•ibbean eo• • øo•a/ini• _..2o(•).... 4o o 27 28 (O =no•hern pa•, •1oo• I 29 30 (S>33.25) ' 31 32 33 34 O• -- • 35 • 36 Salinity (%ø) Fig. 10. Plot of silicate versus salinity for the surfacewaters of the East Carribean cruise track and the Gulf of Paria, fall 1988. the rivedfine waters exist [see also Moore and Todd, this issue]. Spring and Fall Distributions of Primary Productivity On a larger scale, Ryther et al. [1967] suggestedthat Amazon water actually depressedproductivity off Brazil by diluting N nutrients, whereas Mfiller-Karger et al. [1990] suggesta massive fertilization of the eastern Caribbean by Orinoco nutrients. As mentioned previously, the absenceof ammonia and DON data do not permit us to evaluate the nitrogenousinputs to the eastern Caribbean, though clearly in fall there is massive nitrate depletion (Figure 9) before waters exit the Gulf of Padfa. The absenceof small primary productivity values close inshore in either season shows a normally enhancedcoastal productivity, despitethe fact that light, turbidity, and salinity stresses must be strong. The data by Bidigare et al. [this issue] offer confirming evidence from pigment analyses. Thus the primary productivity data give an approximate indication that dfvedfnefertilization takes effect close inshore (Figures 6 and 7). If sufficient N nutrients survive passage through the Gulf of Padfa to fertilize the Caribbean Basin (as assumedby Mfiller-Karger et al. [ 1990]), this effect must be due to efficient biogeochemical recycling processesen route, not to an initially unproductive water mass. Acknowledgments. This work was supported by National Science Foundation grants OCE 8700576 and OCE 8620249, ONR contracts N0001-14-87-K-0007 and N00014-89-J-1258. We thank the Captain and crew of the R/V Columbus lselin for assistanceat sea. This work is partially the result of research sponsoredby NOAA National Sea Grant College Program Office, Department of Commerce, under grant NA86-AA-D-SG090, Woods Hole Oceanographic Institution Sea Grant Project R/O-15-PD. The U.S. Government is authorized to produce and distribute reprints for governmentalpurposesnotwithstandingany copyfight notation that may appear hereon. REFERENCES Benitez-Alvarez, J., and T. Okuda, Distribucitn de nitrtgeno org,•nico particulado en el Golfo de Paria, Bol. Inst. Oceanogr. Univ. Oriente, 15, 3-14, 1976. Bidigare, R. R., M. E. Ondrusek, and J. M. Brooks, Influence of the Orinoco River outflow on distributions of algal pigments in the Caribbean Sea, J. Geophys. Res., this issue. Bonilla, R. J., Condicioneshidroquimicasdel agua y caracteristicas quinicas de los sedimentosdel Golfo de Paria durante la expedicion oceanograficaLS-7302, Bol. Inst. Oceanogr. Univ. Oriente, 16, 99-14, 1977. Bonilla, R. J., and A. L. Lin, Materia org,•nicaen los sedimentosde los Golfos de Paria y Cariaco, Venezuela, Bol. Inst. Oceanogr. Univ. Oriente, 18, 37-52, 1979. Bonilla, R. J., Y. Poyer, and B. Gambra, Caracteristicasgeoqufmicas en nQcleos de sedimentos de la regitn Nororiental y rio Orinoco, Venezuela, Bol. Inst. Oceanogr. Univ. Oriente, 24, 43-61, 1985. Burton, J. D., P.S. Liss, and V. K. Venrigopalan, The behavior of dissolved silicon during estuarine mixing, I, Investigation in Southamptonwater, J. Cons. Perm. lnt. Explor. Mer, 33, 134140, 1970. Butenko, J., J.P. Barbot, and J. Daza, Singularidades geologicas exploradascon el minisubmarinodel Calypso en Venezuela, Rev. Tec. Intevep., 1, 27-35, 1981. Carpenter, E. J., and D. G. Capone (Eds.), Nitrogen in the Marine Environment, Academic, San Diego, Calif., 1983. D'Armas, H., Caracteristicas hidroqufmicas del Golfo de Paria como un sistemaestuarino, M.S. thesis, 158 pp., Inst. Oceanogr. de Venezuela, Univ. de Oriente, Cumanfi, 1986. Edmond, J. M., E. A. Boyle, B. Grant, and R. F. Stallard, The chemical mass balance in the Amazon plume, I, The nutrients, Deep Sea Res., 28A, 1339-1374, 1981. Eisma, D., J. V. Der Gaast, J. M. Martin, and A. J. Thomas, Suspended matter and bottom deposits of the Orinoco Delta: Turbidity,mineralogy andelementary. composition, Neth.J. Sea Res., 12, 224-251, 1978. Froehlich, P. N., Jr., D. K. Atwood, and G. S. Giese, Influence of Amazon River dischargeon surface salinity and dissolved silicate concentrationin the Caribbean Sea, Deep Sea Res., 25,735-744, 1978. Herrera, L. E., and P. Masciangioli, Caracteristicas de las corrientes frente al Delta del Orinoco, sector Occidental del Octano Atlfintico, Rev. Tec. lntevep., 4, 133-134, 1984. BONILLA ET AL ' NUTRIENTSAND PRIMARYPRODUCTIVITYOF THE ORINOCO Kennicutt,M. C., II, C. Barker,J. M. Brooks,D. A. DeFreitas,and G. H. Zhu, Selected organic matter source indicators in the Orinoco,Nile and Changjiangdeltas,Org. Geochem.,14, 41-51, 2257 ria: A possiblemechanismfor the formationof the primarynitrite maximum, J. Mar. Res., 39, 227-238, 1981. Lewis,W. M., andJ. F. Saunders, Concentration andtransportof dissolvedand suspended substances in the OrinocoRiver, Bio- Rodriguez,G., Someaspectsof the ecologyof tropicalestuaries,in TropicalEcologicalSystems:Trendsin Terrestrialand Aquatic Research,Ecol. Stud. Ser. 11, edited by F. B. Golley and E. Medina, pp. 313-333, Springer-Verlag,New York, 1975. geochemistry, 7, 203-240, 1989. Liss, P.S., Conservative and non-conservative behaviour of dis- Ryther, J. H., D. W. Menzel, and N. Corwin, Influence of the Amazon River outflow on the ecologyof the western tropical 1987. solvedconstituents duringestuarinemixing,in EstuarineChemistry, editedby J. D. Burtonand P.S. Liss, vol. 4, pp. 93-130, Atlantic, I, Hydrographyand nutrientchemistry,J. Mar. Res., 25, 69-83, 1967. Salazar,J. C., Condicioneshidrogeoqu/nicas de la regi6nestuarinaAcademic, San Diego, Calif., 1976. deltaicadel Orinocoduranteel mesde noviembrede 1985,M.S. MacDonald,R. W., E. C. Carmack,F. A. McLaughlin,K. Iseki, D. thesis, 128 pp., Inst. Oceanogr.Venezuela, Univ. de Oriente, M. MacDonald, and M. C. O'Brien, Compositionand modificaCuman•, 1989. tionof watermasses in theMackenzieShelfEstuary,J. Geophys. Vanzella, A., M. A. Guerrero, and R. D. Jones, Effect of CO and Res., 94(C12), 18,057-18,070, 1989. light on ammoniumand nitrite oxidationby chemolithotrophic Milliman,J. D., J. Butenko,J.P. Barbot,andJ. Hedberg,Deposibacteria, Mar. Ecol. Prog. Ser., 57, 69-76, 1989. tional patterns of modern Orinoco/Amazon muds on the northern Zika, R. G., P. J. Milne, and O. G. Zafiriou, Photochemicalstudies Venezuelan shelf, J. Mar. Res., 40, 643-657, 1982. of the easternCaribbean:An introductoryoverview,J. Geophys. Moore, W. S., and J. F. Todd, Radium isotopesin the Orinoco Estuaryand easternCaribbeanSea,j. Geophys.Res., this issue. Moore,W. S., J. L. Sarmiento,andR. M. Key, TracingtheAmazon Res., this issue. J. Bonilla and W. Senior, Instituto Oceanogrfificode Venezuela, component of surfaceAtlanticwaterusing228Ra,salinity,and Universidad de Oriente, Cuman,5,Venezuela. silica,J. Geophys.Res., 91, 2574-2580, 1986. Morelock, J., Guyana-Orinococontinentalshelf sediments,Bol. Inst. Oceanogr. Univ. Oriente, 11, 57-61, 1972. M/iller-Karger, F. E., and R. J. Varela, Variabilidad de la biomasa de fitoplanctonen aguas superficialesdel Mar Caribe con el CZCS, EDIMAR FLASA Contrib., 171, 1-23, 1988. J. Bugdenand R. Jones,Departmentof BiologicalSciencesand DrinkingWater ResearchCenter, FloridaInternationalUniversity, Miami, FL 33199. O. Zafiriou,WoodsHole Oceanographic Institution,WoodsHole, MA 02543. M•ller-Karger, F. E., C. R. McClain,T. R. Fisher,W. E. Esaias, and R. Varela, Pigmentdistributionin the Caribbean Sea: Observationsfrom space,Prog. Oceanogr.,23, 23-64, 1990. Olson,R. J., Differentialphotoinhibition of marinenitrifyingbacte- (Received May 10, 1991' revised February 14, 1992' accepted February 21, 1992.)






© Copyright 2026