
Download Supporting Information (PDF)
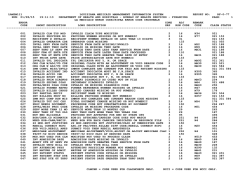
Supporting Information Dabertrand et al. 10.1073/pnas.1420765112 SI Materials and Methods Diameter Measurements. After euthanasia, the brain was removed and placed into chilled (4 °C) Mops-buffered saline. Intraluminal pressures of 60 mmHg and 40 mmHg were used for PCAs and PAs, respectively, unless stated otherwise. Both PCAs and PAs were superfused (4 mL/min) with prewarmed (36 °C ± 1 °C), gassed (gas composition, vol/vol: 5% CO2, 20% O2, 75% N2) artificial cerebrospinal fluid (aCSF) for at least 45 min. Only viable PCAs and PAs, defined as those that developed pressureinduced myogenic tone greater than 15%, were used in subsequent experiments. Endothelial function was tested by assessing the vasodilator response to NS309 (1 μM), an activator of endothelial SK and IK channels. Drugs were applied by addition to the superfusate. Changes in arterial diameter were calculated as the percentage of change from baseline (initial diameter). Maximal dilation was obtained in nominally Ca2+-free aCSF (0 mM [Ca2+]o, 5 mM EGTA, 100 μM papaverine). Third-order mesenteric arteries (∼100 μm internal diameter) were isolated into Hepes–physiological saline solution (HepesPSS), mounted on similar-sized glass pipettes in an organ chamber, and pressurized as described for PCAs. Arteries were pressurized to 80 mmHg for at least 45 min in prewarmed (36 °C ± 1 °C), gassed (gas composition, vol/vol: 5% CO2, 20% O2, 75% N2) PSS. Only arteries that exhibited myogenic constriction to pressure were used in experiments. Changes in arterial diameter were calculated as the percentage of change from baseline (initial diameter). Maximal dilation was obtained in nominally Ca2+-free PSS (0 mM [Ca2+]o, 5 mM EGTA, 100 μM papaverine). Arteriolar Smooth Muscle Membrane Potential Recordings. PAs were isolated, mounted, and pressurized (10 mmHg or 40 mmHg) as described above. Myocytes were then impaled with glass microelectrodes filled with 0.5 M KCl (tip resistance, 150–250 MΩ). A WPI Intra 767 amplifier was used for recording membrane potential. Analog output from the amplifier was obtained using AxoScope (Molecular Devices) software (sample frequency, ∼400 Hz). Criteria for accepting recordings were (i) an abrupt negative deflection of potential as the microelectrode was advanced into the cell, (ii) stable membrane potential for at least 1 min, and (iii) an abrupt change in potential to ∼0 mV after retracting the electrode from the cell. For each PA, membrane potential was recorded in three different cells and averaged. SMC Isolation and Perforated Patch-Clamp Experiments. Cerebral arteries (anterior, middle, and posterior) and arterioles were cleaned of connective tissue and placed in cell isolation solution (see Solutions and Drugs for composition). Single SMCs were isolated from cerebral arteries by enzymatic digestion in papain (0.5 mg/mL) and dithioerythritol (1 mg/mL) for 12 min, followed by a second digestion in collagenase type 4 (1 mg/mL) without Ca2+ for 10 min. Digested tissue was washed three times and gently triturated with a fire-polished glass pipette to release the SMCs. The single-cell suspension of myocytes was refrigerated until use (typically 4–6 h). Third-order mesenteric arteries were enzymatically digested with papain (2.5 mg/mL), dithioerythritol (1 mg/mL), and BSA for 17 min, followed by a second digestion in collagenase type F (0.6 mg/mL) and CaCl2 (100 μM) for 6 min. Outward K+ currents were recorded from single cells in the presence of 1 μM paxilline (to block BK currents) at room temperature, using the perforated-cell configuration of the patchclamp technique. Recording electrodes with resistances of Dabertrand et al. www.pnas.org/cgi/content/short/1420765112 2–4 MΩ were pulled from borosilicate glass and backfilled with a pipette solution of appropriate composition (Solutions and Drugs). Currents were recorded from cells using an Axopatch 200B amplifier, filtered at 2 kHz using a low-pass Bessel filter, and digitized at 10 kHz (Digidata 1322A; Molecular Devices). pCLAMP-9 software (Molecular Devices) was used for data recording and analysis. NOTCH3ECD Immunohistochemistry. Mice (TgNotch3R169C, TgNotch3WT, and Non-Tg) were overdosed with isoflurane. Second- and thirdorder mesenteric arteries were dissected under a microscope, flash frozen in liquid nitrogen, and stored at −80 °C until use. For immunohistochemistry, arteries were transferred to Netwell inserts (VWR International) with an 80-μm mesh size polyester membrane, briefly rinsed in PBS, and fixed in 100% ethanol for 20 min. After rinsing in PBS, arteries were blocked and permeabilized by incubating in a PBS solution containing 5% BSA and 0.4% Triton X-100 for 2 h at room temperature. Arteries were then incubated overnight at 4 °C with primary antibodies against NOTCH3ECD (mouse monoclonal, clone 5E1) and smooth muscle myosin heavy chain (rabbit polyclonal; Biomedical Technologies), diluted 1:2 and 1:100, respectively, in PBS/0.5% BSA. The next day, arteries were washed with PBS and then incubated for 2 h at room temperature with goat anti-mouse Alexa 488 and goat anti-rabbit Alexa 594 secondary antibodies (Life Technologies) in PBS/0.5% BSA. All steps in the immunochemistry protocol were performed with gentle shaking. Nuclei were counterstained with DAPI (4′,6-diamidino-2-phenylindole), and arteries were coverslip mounted on glass slides with fluorescent mounting medium (Dako). Arteries were imaged on a Nikon eclipse 80i microscope (40× oil objective, NIS elements AR Software, v4.2; Nikon), and images were deconvolved using the AutoQuant Blind Deconvolution plug-in for NIS elements. Solutions and Drugs. The composition of Mops-buffered saline was 135 mM NaCl, 5 mM KCl, 1 mM KH2PO4, 1 mM MgSO4, 2.5 mM CaCl2, 5 mM glucose, 3 mM Mops, 0.02 mM EDTA, 2 mM pyruvate, 10 mg/mL BSA, pH 7.3 (at 4 °C). The composition of aCSF was 125 mM NaCl, 3 mM KCl, 26 mM NaHCO3, 1.25 mM NaH2PO4, 1 mM MgCl2, 4 mM glucose, 2 mM CaCl2, pH 7.3 [with aeration with 5% (vol/vol) CO2]. The composition of Hepes-PSS was 134 mM NaCl, 6 mM KCl, 1 mM MgCl2, 10 mM Hepes, 7 mM glucose, 2 mM CaCl2, pH 7.4 (at 4 °C). The composition of PSS was 119 mM NaCl, 4.7 mM KCl, 24 mM NaHCO3, 1.2 mM KH2PO4, 0.026 mM EDTA, 1.2 mM MgCl2, 7 mM glucose, 2 mM CaCl2, pH 7.4 [with aeration with 5% (vol/vol) CO2]. The composition of cell isolation solution was 60 mM NaCl, 85 mM Na-glutamate, 3 mM KCl, 2 mM MgCl2, 10 mM Hepes, 10 mM glucose, 7 mM mannitol, pH 7.4. For patch-clamp experiments using cerebral myocytes, the bath solution composition was 137 mM NaCl, 3 mM KCl, 0.1 mM CaCl2, 4 mM glucose, 10 mM Hepes (pH 7.3) and contained paxilline (1 μM); the pipette solution was 10 mM NaCl, 30 mM KCl, 110 mM K-aspartate, 1 mM MgCl2, 10 mM Hepes (pH 7.2) and contained 250 μg/mL amphotericin B. Paxilline was purchased from A.G. Scientific. Apamin and charybdotoxin were purchased from Enzo Life Sciences. Papain and collagenase type 4 were purchased from Worthington Biochemical Corporation. All other chemicals were purchased from Sigma-Aldrich. The vehicle for HB-EGF solutions was 0.2-μm– filtered PBS containing 0.1% BSA. 1 of 7 Fig. S1. Stromatoxin-induced vasoconstriction is not modified by the introduction of a CADASIL-causing mutation in pial arteries or parenchymal arterioles. (A and C) Representative inner diameters recorded as a function of time from pressurized (60 mmHg) posterior cerebral arteries (A) or pressurized (40 mmHg) parenchymal arterioles (C) before, during, and after exposure to stromatoxin (20 nM). (B and D) Bar plot showing the mean ± SEM constriction (given as a percentage) induced by stromatoxin; the number of animals is shown in parentheses. Dabertrand et al. www.pnas.org/cgi/content/short/1420765112 2 of 7 Fig. S2. The 4-AP–induced constriction is increased in pial arteries from CADASIL mice. (A, a–c) Representative traces showing constriction of pial arteries induced by the SK channel blocker apamin (300 nM), the BK channel blocker paxilline (1 μM), and the IK channel blocker charybdotoxin (100 nM). (B) Summary data (mean ± SEM). (C, a and b) Representative traces showing constriction of pressurized (60 mmHg) pial arteries induced by the Kv channel blocker 4-AP (1 mM) and the endothelial nitric oxide synthase (eNOS) inhibitor L-NAME (100 μM). (D) Summary data (mean ± SEM). The number of animals is shown in parentheses. *P < 0.05, one-way ANOVA. Dabertrand et al. www.pnas.org/cgi/content/short/1420765112 3 of 7 Fig. S3. The 4-AP–sensitive currents are significantly increased in cerebral artery SMCs from TgNotch3R169C mice. (A) Families of normalized, 4-AP (1 mM)– sensitive KV currents from isolated cerebral SMCs elicited by voltage pulses from −70 mV to +60 mV in the presence of 1 μM paxilline (to block BK channel currents). (B) Graph summarizing 4-AP–sensitive current densities. Currents were normalized to cell capacitance, and values recorded in the presence of 1 mM 4-AP were subtracted from those recorded in the absence of 4-AP (*P < 0.05, one-way ANOVA). Fig. S4. Notch3ECD accumulates in the mesenteric arteries of TgNotch3R169C mice at 6 mo of age. Shown are representative images of third-order mesenteric arteries of 6-mo-old TgNotch3WT (A and B) and TgNotch3R169C (C and D) mice stained with anti-smooth muscle myosin heavy chain (anti-MHC) (A and C) and anti-Notch3ECD (B and D) antibodies. The extracellular domain of Notch3 accumulates in TgNotch3R169C (D) but not TgNotch3WT (B) mesenteric arteries. (Scale bar, 50 μm.) Dabertrand et al. www.pnas.org/cgi/content/short/1420765112 4 of 7 Table S1. Mean values of passive diameter (measured in the absence of extracellular Ca2+), active diameter (after development of myogenic tone), and percentage of tone of the arteries and arterioles used in the study Vascular bed (pressure, mmHg) Parenchymal arterioles (40) Non-Tg TgNotch3WT TgNotch3R169C Pial arteries (60) Non-Tg TgNotch3WT TgNotch3R169C Mesenteric arteries (80) Non-Tg TgNotch3WT TgNotch3R169C Passive diameter, μm Active diameter, μm Tone, % 30.6 ± 2.7 31 ± 1.8 30.3 ± 1.3 18.4 ± 1.5 19.3 ± 1.3 23 ± 1.5 39.3 ± 1.8 37.9 ± 1.8 24.4 ± 2.5* 170.1 ± 2.3 163.7 ± 3.4 150.1 ± 4.4* 120.5 ± 4.7 114.8 ± 2.3 121.8 ± 2.9 28.8 ± 1.3 27.9 ± 1.9 17.9 ± 2.4* 122.1 ± 7.2 136 ± 6.5 136 ± 7.2 96.7 ± 7.4 107.7 ± 6.6 103.4 ± 7.6 21.5 ± 2 21.3 ± 1.2 24.3 ± 2.2 *P < 0.05; one-way ANOVA followed by Tukey’s post hoc test. Dabertrand et al. www.pnas.org/cgi/content/short/1420765112 5 of 7 Table S2. Comparison of cerebral KV and mesenteric KV, KV1, KV2, KV3, KV4, and KV7 current properties Current (ref.) V0.5, mV Slope k, mV τact, ms τdeact, ms 4-AP sensitive below 1 mM Cerebral KV currents Non-Tg TgNotch3WT TgNotch3R169C 6±1 3.7 ± 0.8 2.6 ± 0.6 14 ± 1 14.3 ± 0.8 15.8 ± 0.6 43 ± 3.6 34 ± 3.5 41.1 ± 6.5 28 ± 3.1 27.4 ± 3.3 26.7 ± 3.4 + + + −5.7 ± 1.6 −3.7 ± 1.2 14.6 ± 1.6 14.9 ± 1.9 31.1 ± 8.8 35.3 ± 12.1 32.5 ± 2.8 29.3 ± 1.7 + + Mesenteric KV currents Non-Tg TgNotch3R169C KV currents in expression systems KV1.1 (1, 2) KV1.2 (2, 3) KV1.3 (2) KV1.4 (1, 4) KV1.5 (2, 5, 6) −32 ± 2 −5 ± 4 to 27 ± 6 −26 ± 8 −12.8 ± 3.4 −14 ± 3 to 5 ± 2.1 8.5 ± 0.5 14 14 ± 5 13 ± 2 23 23 ± 7 7±1 38 39 ± 6 13.4 15–55 NR 9.1 ± 1.7 to 12 ± 1 0.4 ± 0.1 (fast) and 15.3 ± 1.1 (fast) and 1.6 ± 0.2 (slow) 59.5 ± 5.6 (slow) KV1.6 (7) −20.8 ± 7.5 8.1 ± 1.5 NR NR KV1.7 (8) −8 NR NR NR KV1.8 (9) 3.6 NR 18 NR KV2.1 (7, 10, 11) −1.7,12, and 20.5 3–9.6 15.6 ± 0.9 20.9 ± 2.1 KV2.1/KV5.1 (10, 11) 18.5 17.3 20.4 ± 1.2 24.5 ± 2.3 KV2.1/KV6.1 (10, 11) −9.4 11.8 28.2 ± 2.0 24.4 ± 0.7 KV2.1/KV6.2 (12) −10 NR NR NR KV2.1/KV6.3 (13, 14) −13.3 ± 1.8 to −4.2 ± 0.7 9.8 ± 0.4 58 ± 5 NR KV2.1/KV8.1 (15) 10.2 ± 0.9 NR 27.2 ± 2.5 NR KV2.1/KV9.1 (11) NR NR 14.3 ± 0.8 36.4 ± 6.4 KV2.1/KV9.2 (11) NR NR 20.3 ± 1.3 45.2 ± 2.1 KV2.1/KV9.3 (16) −8.7 ± 1.2 12.7 ± 1.1 NR NR KV2.2 (11, 17) −16.6 ± 1.1 18 21.8 ± 0.5 39.1 ± 5.1 KV2.2/KV5.1 (11) NR NR 24.5 ± 2.3 130.8 ± 9.7 KV2.2/KV6.1 (11) NR NR 24.4 ± 0.7 147.7 ± 13.8 KV2.2/KV8.1 (15) 8.5 ± 1.4 NR 45.4 ± 3.6 NR KV2.2/KV9.1 (11) NR NR 17.9 ± 0.3 27.2 ± 0.8 KV2.2/KV9.2 (11) NR NR 23.6 ± 1.4 43.4 ± 1.2 KV3.1 (2) 16 ± 1 8.7 ± 0.4 NR 1.4 ± 0.2 KV3.2 (6, 18, 19) 6 ± 2.4 to 12.1 ± 1.3 8.4 ± 0.3 NR NR KV3.3 (1, 19) −3.4 ± 5.1 to 7 6–8.4 ± 0.4 NR NR KV3.4 (1, 20) 13–19 7–11 NR NR KV4.1 (1, 21) −47 22 NR NR KV4.2 (1, 22) −20 to −4 13–20 NR 40 KV4.3 (1, 23) −29.1 ± 0.7 to −20 4.5–13 NR 20–40 KV7.1 (24-27) −23.6 ± 1.6 to −18 ± 1.6 11.1–12.6 144 ± 30 621.4 ± 27 to 740 KV7.2 (24, 28, 29) −37 ± 2 to −27.3 ± 2.4 NR 35.6 ± 5.7 and 106.7 ± 7.1 and 149.1 ± 11.8 642.3 ± 62.2 KV7.2/KV7.3 (24, 29, 30) −40 ± 1 to −26.4 ± 2.3 6.8 ± 0.1 50.1 ± 3.4 and 144 ± 30 and 695 239.3 ± 17.5 KV7.3 (24, 29, 31) −36 ± 1.3 5.5 ± 1.1 NR NR KV7.4 (24, 31) −18.6 ± 0.3 to −0.6 ± 3.5 9.8 ± 0.2 636.3 ± 45.6 49 ± 4.2 and 350.3 ± 27.5 KV7.5 (24, 32) −46 ± 1 to −25.1 ± 2.6 NR 37.2 ± 2.2 and NR 247 ± 17 + + + + + − + − − NR NR NR NR NR NR NR NR − NR NR NR NR NR + + − + − − − − − − − NR NR Half-maximal activation (V0.5) voltage, slope (k), activation time constant (τact), deactivation time constant (τdeact), and 4-AP sensitivity were used as channel fingerprints to identify the KV channel subtype involved in cerebral and mesenteric arteries. Vascular SMCs potentially express KV1–9 subtypes (19, 33, 34); parameters of currents recorded in native myocytes point to a predominant role for the KV1 channel subfamily in both cerebral and mesenteric SMCs. NR, not reported. 1. Coetzee WA, et al. (1999) Molecular diversity of K+ channels. Ann N Y Acad Sci 868:233–285. 2. Grissmer S, et al. (1994) Pharmacological characterization of five cloned voltage-gated K+ channels, types Kv1.1, 1.2, 1.3, 1.5, and 3.1, stably expressed in mammalian cell lines. Mol Pharmacol 45(6):1227–1234. 3. Ramaswami M, et al. (1990) Human potassium channel genes: Molecular cloning and functional expression. Mol Cell Neurosci 1(3):214–223. 4. Judge SIV, Yeh JZ, Goolsby JE, Monteiro MJ, Bever CT, Jr (2002) Determinants of 4-aminopyridine sensitivity in a human brain kv1.4 k(+) channel: Phenylalanine substitutions in leucine heptad repeat region stabilize channel closed state. Mol Pharmacol 61(4):913–920. 5. Clément-Chomienne O, Ishii K, Walsh MP, Cole WC (1999) Identification, cloning and expression of rabbit vascular smooth muscle Kv1.5 and comparison with native delayed rectifier K+ current. J Physiol 515(Pt 3):653–667. 6. Nilsson J, Madeja M, Arhem P (2003) Local anesthetic block of Kv channels: Role of the S6 helix and the S5-S6 linker for bupivacaine action. Mol Pharmacol 63(6):1417–1429. Dabertrand et al. www.pnas.org/cgi/content/short/1420765112 6 of 7 7. Grupe A, et al. (1990) Cloning and expression of a human voltage-gated potassium channel. A novel member of the RCK potassium channel family. EMBO J 9(6):1749–1756. 8. Bardien-Kruger S, et al. (2002) Characterisation of the human voltage-gated potassium channel gene, KCNA7, a candidate gene for inherited cardiac disorders, and its exclusion as cause of progressive familial heart block I (PFHBI). Eur J Hum Genet 10(1):36–43. 9. Lang R, et al. (2000) KCNA10: A novel ion channel functionally related to both voltage-gated potassium and CNG cation channels. Am J Physiol Renal Physiol 278(6):F1013–F1021. 10. Kramer JW, Post MA, Brown AM, Kirsch GE (1998) Modulation of potassium channel gating by coexpression of Kv2.1 with regulatory Kv5.1 or Kv6.1 alpha-subunits. Am J Physiol 274(6 Pt 1):C1501–C1510. 11. Salinas M, Duprat F, Heurteaux C, Hugnot JP, Lazdunski M (1997) New modulatory alpha subunits for mammalian Shab K+ channels. J Biol Chem 272(39):24371–24379. 12. Zhu XR, Netzer R, Böhlke K, Liu Q, Pongs O (1999) Structural and functional characterization of Kv6.2 a new gamma-subunit of voltage-gated potassium channel. Receptors Channels 6(5):337–350. 13. Sano Y, et al. (2002) Molecular cloning and characterization of Kv6.3, a novel modulatory subunit for voltage-gated K(+) channel Kv2.1. FEBS Lett 512(1–3):230–234. 14. Ottschytsch N, Raes A, Van Hoorick D, Snyders DJ (2002) Obligatory heterotetramerization of three previously uncharacterized Kv channel alpha-subunits identified in the human genome. Proc Natl Acad Sci USA 99(12):7986–7991. 15. Salinas M, de Weille J, Guillemare E, Lazdunski M, Hugnot JP (1997) Modes of regulation of shab K+ channel activity by the Kv8.1 subunit. J Biol Chem 272(13):8774–8780. 16. Hulme JT, Coppock EA, Felipe A, Martens JR, Tamkun MM (1999) Oxygen sensitivity of cloned voltage-gated K(+) channels expressed in the pulmonary vasculature. Circ Res 85(6):489–497. 17. Schmalz F, et al. (1998) Molecular identification of a component of delayed rectifier current in gastrointestinal smooth muscles. Am J Physiol 274(5 Pt 1):G901–G911. 18. Hernández-Pineda R, et al. (1999) Kv3.1-Kv3.2 channels underlie a high-voltage-activating component of the delayed rectifier K+ current in projecting neurons from the globus pallidus. J Neurophysiol 82(3):1512–1528. 19. Cox RH (2005) Molecular determinants of voltage-gated potassium currents in vascular smooth muscle. Cell Biochem Biophys 42(2):167–195. 20. Elinder F, Madeja M, Arhem P (1996) Surface charges of K channels. Effects of strontium on five cloned channels expressed in Xenopus oocytes. J Gen Physiol 108(4):325–332. 21. Jerng HH, Shahidullah M, Covarrubias M (1999) Inactivation gating of Kv4 potassium channels: Molecular interactions involving the inner vestibule of the pore. J Gen Physiol 113(5):641–660. 22. Tseng GN, Jiang M, Yao JA (1996) Reverse use dependence of Kv4.2 blockade by 4-aminopyridine. J Pharmacol Exp Ther 279(2):865–876. 23. Faivre JF, et al. (1999) Characterisation of Kv4.3 in HEK293 cells: Comparison with the rat ventricular transient outward potassium current. Cardiovasc Res 41(1):188–199. 24. Blom SM, Schmitt N, Jensen HS (2009) The acrylamide (S)-2 as a positive and negative modulator of Kv7 channels expressed in Xenopus laevis oocytes. PLoS ONE 4(12):e8251. 25. Barhanin J, et al. (1996) K(V)LQT1 and lsK (minK) proteins associate to form the I(Ks) cardiac potassium current. Nature 384(6604):78–80. 26. Yang WP, et al. (1997) KvLQT1, a voltage-gated potassium channel responsible for human cardiac arrhythmias. Proc Natl Acad Sci USA 94(8):4017–4021. 27. Chouabe C, et al. (1997) Properties of KvLQT1 K+ channel mutations in Romano-Ward and Jervell and Lange-Nielsen inherited cardiac arrhythmias. EMBO J 16(17):5472–5479. 28. Biervert C, et al. (1998) A potassium channel mutation in neonatal human epilepsy. Science 279(5349):403–406. 29. Yang WP, et al. (1998) Functional expression of two KvLQT1-related potassium channels responsible for an inherited idiopathic epilepsy. J Biol Chem 273(31):19419–19423. 30. Wang HS, et al. (1998) KCNQ2 and KCNQ3 potassium channel subunits: Molecular correlates of the M-channel. Science 282(5395):1890–1893. 31. Selyanko AA, et al. (2000) Inhibition of KCNQ1-4 potassium channels expressed in mammalian cells via M1 muscarinic acetylcholine receptors. J Physiol 522(Pt 3):349–355. 32. Schroeder BC, Hechenberger M, Weinreich F, Kubisch C, Jentsch TJ (2000) KCNQ5, a novel potassium channel broadly expressed in brain, mediates M-type currents. J Biol Chem 275(31): 24089–24095. 33. Gutman GA, et al. (2005) International Union of Pharmacology. LIII. Nomenclature and molecular relationships of voltage-gated potassium channels. Pharmacol Rev 57(4):473–508. 34. Jepps TA, Olesen SP, Greenwood IA (2013) One man’s side effect is another man’s therapeutic opportunity: Targeting Kv7 channels in smooth muscle disorders. Br J Pharmacol 168(1): 19–27. Dabertrand et al. www.pnas.org/cgi/content/short/1420765112 7 of 7



© Copyright 2026