
Leer Más
1. LOS ELEMENTOS Y MOLÉCULAS BIOLÓGICAMENTE IMPORTANTES. En las células vivas encontramos un conjunto de elementos químicos que reciben el nombre de elementos biogénicos o bioelementos, comunes al mundo inanimado. De los aproximadamente 100 elementos químicos presentes en la corteza terrestre, unos 25 se encuentran en todos los seres vivos. Seis de estos elementos (C, H, N, O, P, S), llamados primarios, constituyen más del 99% del peso de la célula. El átomo de C, debido a su reducido tamaño y a sus cuatro electrones en su capa externa, puede formar cuatro enlaces covalentes fuertes con otros átomos. También se puede unir con otros átomos de C formando cadenas y anillos, y dando lugar a moléculas grandes y complejas. Los otros átomos abundantes en las células también son pequeños y capaces de formar enlaces covalentes muy fuertes. El resto de los elementos presentes en las células se hallan en proporciones más pequeñas y reciben el nombre de bioelementos secundarios (Na, Mg, Cl, Fe) y oligoelementos (Cu, Mn, Co, I, Zn…); estos últimos se encuentran en una proporción inferior al 0,1%, sin embargo desempeñan importantes funciones en los seres vivos. Los bioelementos se combinan entre sí para formar las moléculas componentes de las células, que pueden separarse mediante métodos físico. Estas sustancias, que forman parte de los seres vivos se denominan biomoléculas o principios inmediatos. ENLACES COVALENTES Y GRUPOS QUÍMICOS FRECUENTES EN LAS MOLÉCULAS ORGÁNICAS. ENLACES COVALENTES: Los átomos de las moléculas orgánicas suelen estar unidos por enlaces covalentes. Cada átomo puede formar un número determinado de enlaces covalentes con una disposición espacial definida. También pueden formar dobles enlaces con una disposición espacial diferente. ESQUELETOS DE CARBONO: La importancia del carbono en los seres vivos se debe a su capacidad para formar fuertes enlaces covalentes con otros átomos de carbono. Así, se pueden unir formando cadenas, o estructuras ramificadas, o anillos. HIDROCARBUROS: Están formados por la unión de carbono e hidrógeno. Son compuestos no polares y, por lo general, insolubles en agua. RESONANCIA: Cuando se produce resonancia en un compuesto cíclico, se genera un anillo aromático. La cadena de carbonos puede incluir dobles enlaces. Si éstos se encuentran en átomos de carbono alternos, los electrones de enlace se mueven dentro de la molécula, estabilizando la estructura por resonancia. COMPUESTOS C-O: Muchos compuestos orgánicos contienen un carbono unido a un oxígeno. Por ejemplo: El –OH recibe el nombre de grupo hidroxilo. El C=O recibe el nombre de grupo carboxilo. El –COOH recibe el nombre de grupo carboxilo. En agua pierde un ión H+ y se convierte en –COO-. Los ésteres están formados por la combinación de un ácido y un alcohol. COMPUESTOS C-N: Las aminas y las amidas son dos ejemplos de compuestos que tienen un carbono unido a un nitrógeno. Las aminas, en agua, se combinan con ión H+ y quedan cargadas positivamente. Por lo tanto, son básicas. Las amidas se forman por la combinación de un ácido y una amina. A diferencia de las aminas, no poseen carga en solución acuosa. El nitrógeno se presenta también en diversos compuestos cíclicos como las bases nitrogenadas. FOSFATOS: El fosfato inorgánico es un ión estable formado a partir del ácido fosfórico, H3PO4. A menudo se representa como P o Pi. MOLÉCULAS INORGÁNICAS: EL AGUA El agua es el compuesto más abundante de la vida. Normalmente suele constituir entre el 6095% de la masa de un organismo. Sin agua, la vida no sería posible. Su importancia para los seres vivos es doble, ya que a la vez es un componente fundamental de las células y, para muchos, su hábitat. Se trata de una sustancia de características especiales. Las especiales propiedades del agua derivan de su estructura molecular, la cual presenta tres características fundamentales: su pequeño tamaño, su polaridad y su gran capacidad para formar enlaces entre sus propias moléculas. La polaridad es debida a una desigual distribución de los electrones, lo cual hace que la zona de la molécula próxima al O sea débilmente negativa y la del H débilmente positiva. Cada molécula de agua forma, por tanto, un dipolo. El enlace que se forma entre dos moléculas de agua recibe el nombre de puente de hidrógeno. Estos enlaces tienen una fuerza de aproximadamente 1/20 de la de un enlace covalente. Las moléculas de agua se unen transitoriamente por enlaces de hidrógeno formando redes. Debido a su naturaleza polar, las moléculas de agua se agrupan alrededor de los iones y de otras moléculas polares. Estas sustancias serán, por consiguiente, hidrofílicas e hidrosolubles. Las moléculas no polares interrumpen la estructura a través de enlaces H del agua. Son por tanto hidrofóbicas y bastante insolubles en agua. PROPIEDADES Y FUNCIONES DEL AGUA PROPIEDAD Disolvente Cohesión y Adhesión Calor Específico y Conductividad térmica Calor de Vaporización y Calor de Fusión Capilaridad CARACTERÍSTICAS El agua es un buen disolvente de sustancias polares. Estas sustancias se denominan hidrofílicas. Las sustancias no polares o hidrofóbicas no son solubles en el agua y tienden a agruparse, formando interfases con el agua. Es la capacidad de mantener unidas entre sí las moléculas de una misma sustancia. En el caso del agua es mayor que en otras sustancias. Esta propiedad influye en la adhesión del agua, es decir, en su capacidad de unirse a otra sustancia, la cual es también mayor que en otros líquidos. El agua tiene un alto calor específico y una buena conductividad térmica. Por tanto, una gran variación de energía calorífica provocará un cambio de temperatura relativamente pequeño. Ambos son altos. En consecuencia, cualquiera de los cambios de estado citados va a captar o liberar grandes cantidades de energía. Es la propiedad de una sustancia, por la que ésta tiene la capacidad de ascender por un capilar. FUNCIÓN BIOLÓGICA La mayoría de las reacciones metabólicas se dan en agua. Permite el transporte y la difusión de sustancias. Formación de membranas. Transporte de savia en las plantas. Absorción. Regulación de la temperatura en organismos. Distribución del calor. Regulación de la temperatura (sudoración, prevención del congelamiento). EL AGUA COMO DISOLVENTE: DISPERSIONES Y DISOLUCIONES Se denomina dispersión a la repartición homogénea de partículas pequeñas de una sustancia (fase dispersa o soluto) en el seno de otra (fase dispersante o disolvente). Si la fase dispersante está en estado líquido, hablamos de disoluciones. Se distinguen tres tipos de dispersiones: - Dispersión grosera: Donde las partículas son visibles a simple vista o al microscopio. Dispersión coloidal: Donde las partículas son visibles al ultramicroscopio. Dispersión molecular: Donde las partículas son invisibles. Las disoluciones coloidales pueden presentarse en dos estados. SOL, que es el estado coloidal propiamente dicho, y GEL que es un coloide que ha perdido agua. El paso de SOL a GEL está determinado por numerosos factores como pH, temperatura, concentración salina… Algunas de las propiedades de las disoluciones, de interés biológico son: - - DIFUSIÓN: Es la distribución uniforme de las partículas de un fluido en el seno de otro. Este proceso se debe al continuo movimiento en que se encuentran las partículas de los líquidos y los gases. La difusión ocurre a favor de un gradiente, es decir, las partículas se mueven de una región de mayor concentración a una región de menor concentración. La difusión es un proceso fundamental para el movimiento de sustancias dentro de las células. ÓSMOSIS: Es un tipo especial de difusión que se manifiesta al poner en contacto dos disoluciones de distinta concentración separadas por una membrana semipermeable. La tendencia a igualar las concentraciones se pone de manifiesto con el paso de disolvente (NO SOLUTO), de la disolución más diluida a la más concentrada. La presión mecánica para contrarrestar el paso de agua a través de la membrana semipermeable se denomina presión osmótica. Si comparamos dos disoluciones de distinta concentración la de mayor concentración se denomina hipertónica, con respecto a la de menor concentración, que se denomina hipotónica. Si las dos disoluciones tienen la misma concentración se denominan isotónicas. Si el medio externo es hipotónico, la célula se hinchará. A este fenómeno se le conoce como turgencia. Si, por el contrario, el medio externo es hipertónico, la célula perderá agua, fenómeno conocido como plasmólisis. IONIZACIÓN DEL AGUA. ÁCIDOS Y BASES. Un ácido es una sustancia que, en disolución cede un ión H+ (protón). Una base es una sustancia que, en disolución, capta un ión H+ (protón). El agua, pues, por sí misma, tiene una ligera tendencia a ionizarse, pudiendo actuar así como ácido y como base. El pH es la concentración de protones que hay en un medio acuoso. El mantenimiento de un pH constante y próximo a la neutralidad en el medio interno es necesario para que puedan desarrollarse las reacciones metabólicas correspondientes. Todos los seres vivos mantienen el pH de su medio interno constante gracias a las llamadas soluciones amortiguadoras o tampón. Estos sistemas tampón consisten en un par ácido-base conjugada, y mantienen el pH constante por su tendencia a combinarse con iones H+, eliminándolos así del medio si su concentración es demasiado alta, o liberándolos al medio cuando su concentración sea demasiado baja. Los dos sistemas tampón más importantes son: - Sistema tampón fosfato. Constituido por los iones dihidrógeno fosfato y monohidrógeno fosfato. Este sistema amortiguador mantiene el pH intracelular en 7,2. Sistema tampón bicarbonato. Es el principal sistema tampón en la sangre de las personas. Está constituido por el par ácido-base H2CO3-HCO3. El ácido carbónico se disocia en protones e iones bicarbonato, y a su vez el ácido carbónico está en equilibrio con el CO2 disuelto en la sangre. MOLÉCULAS INORGÁNICAS: LAS SALES MINERALES En todos los seres vivos encontramos determinadas cantidades de sales minerales. Estas pueden ser insolubles en agua y depositarse en los órganos esqueléticos para darles consistencia, o pueden ser solubles en agua. Éstas últimas se encuentran disociadas en sus iones. MOLÉCULAS ORGÁNICAS En los organismos se encuentran cuatro tipos diferentes de moléculas orgánicas: glúcidos, lípidos, proteínas y ácidos nucleicos. Todas estas moléculas contienen carbono, hidrógeno y oxígeno. Además algunas contienen nitrógeno, fósforo y azufre. GLÚCIDOS o HIDRATOS DE CARBONO. Los glúcidos son las sustancias más numerosas presentes en los organismos vivos; también son conocidos por los nombres de hidratos de carbono y de azúcares. Su forma empírica general es Cx(H2O)y. Los glúcidos se dividen en tres clases principales: monosacáridos, disacáridos y polisacáridos. Los monosacáridos constituyen las unidades a partir de las cuales se forman los demás. GLÚCIDOS AZÚCARES MONOSACÁRIDOS - - Moléculas pequeñas (baja Mt). Sabor dulce. Solubles en agua. Cristalinos. “Azúcares simples”. (CH2O)n Todos son reductores. POLISACÁRIDOS DISACÁRIDOS - - Moléculas pequeñas. Sabor dulce. Solubles en agua. Cristalinos. Se forman por la unión de dos monosacárido s por enlace glucosídico. C12H22O11 (dos hexosas). Algunos reductores, otros no. - - Macromoléculas (alta Mt). No tienen sabor dulce. Insolubles o poco solubles en agua. No cristalinos. Se forman por la unión de muchos monosacáridos por enlaces glucosídicos. Su fórmula general: Cx(H2O)y. No reductores. LOS MONOSACÁRIDOS El número de carbonos de cada monosacárido es el que determina su clasificación en triosas, tetrosas… Estos azúcares son aldehídos y cetonas polihidroxílicos, clasificados en aldosas y cetosas. Los monosacáridos desempeñan dos importantes funciones en las células. Por una parte, son fuentes de energía al oxidarse en la respiración celular, por otra parte son las unidades de síntesis de otras moléculas más complejas. ISOMERÍA Siempre que dos sustancias distintas tengan la misma fórmula molecular se dice que ambas son isómeras entre sí. Existen dos tipos de isomería: estructural y espacial. La isomería espacial o estereoisomería es una característica de todos los compuestos que poseen en su molécula algún carbono unido a cuatro radicales diferentes. Estos carbonos se denominan asimétricos. Los dos estereoisómeros de cada monosacárido se denominan D y L. Para reconocerlos hay que fijarse en el carbono más alejado del aldehído o cetona. Si su –OH se sitúa a la derecha, será D, si se encuentra a la izquierda, L. Estas formas D y L se denominan estereoisómeros enantiomorfos y son imágenes especulares. Estos estereoisómeros poseen actividad óptica y son capaces de desviar el plano de vibración de la luz polarizada. Si la desviación se produce hacia la derecha, el azúcar se denomina dextrógiro y se representa con un (+). Si se produce hacia la izquierda, se trata de un monosacárido levógiro, y se representa por el signo (-). NO EXISTE RELACIÓN ALGUNA ENTRE LOS ISÓMEROS DEXTRÓGIRO Y LEVÓGIRO Y LAS CONFIGURACIONES D Y L. FORMAS CÍCLICAS (representaciones de Haworth) Cuando las moléculas de monosacáridos se disuelven en agua su esqueleto se pliega sobre sí mismo y puede cerrarse dando lugar a una estructura anular. El enlace (hemiacetálico) se establece entre el grupo carbonilo y un hidroxilo de la misma molécula, formándose un anillo pentagonal (furanósico) o hexagonal (piranósico). Si comparamos las figuras anulares con la abierta, veremos que los grupos –OH de los carbonos asimétricos que estaban a la derecha del eje, ahora se sitúan debajo del plano hexagonal, y los de la izquierda, encima. El problema es que al ciclarse aparece un nuevo carbono asimétrico, que recibe el nombre de carbono anomérico y, por tanto, son posibles dos nuevas formas isoméricas (anómeros). En el caso de la D-glucosa, al ciclarse, puede acabar con el –OH del carbono 1 debajo o encima del plano, isómeros que se denomina alfa y beta, respectivamente. TRIOSAS TETROSAS PENTOSAS HEXOSAS PRINCIPALES FUNCIONES DE LOS MONOSACÁRIDOS Intermediarios en la respiración celular, en la fotosíntesis y en otras vías metabólicas de los seres vivos. Raras en la naturaleza. Principalmente se encuentran en las bacterias. - Síntesis de ácidos nucleicos. - Síntesis de coenzimas. - Síntesis de AMP, ADP y ATP. - Síntesis de algunos polisacáridos. Fuente de energía en la respiración celular. La glucosa es el sustrato más común en la respiración y el más común de los monosacáridos. - Síntesis de disacáridos. - Síntesis de polisacáridos. LOS DISACÁRIDOS Se forman por la unión de dos moléculas de monosacáridos, generalmente hexosas, mediante un enlace glucosídico. Este tipo de reacción se denomina de condensación y se realiza siempre con desprendimiento de una molécula de agua. El enlace glucosídico puede romperse añadiendo agua, reacción denominada hidrólisis. El enlace glucosídico puede establecerse entre el carbono anomérico de un monosacárido y uno de los dos carbonos alcohólicos del otro monosacárido. Pero también puede establecerse entre los carbonos anoméricos de los monosacáridos. En el primer caso, el disacárido será un reductor debido a que posee un hidroxilo unido a un carbono anomérico libre. En el segundo caso, el disacárido no tendrá poder reductor. MALTOSA: Está formada por la unión de dos moléculas de glucosa. La primera glucosa siempre posee configuración alfa, mientras que la segunda puede ser alfa o beta. El enlace se establece entre el carbono 1 de la primera glucosa y el carbono 4 de la segunda; enlace α (1 – 4). LACTOSA: Resulta de la unión de una molécula de galactosa y otra de glucosa mediante enlace glucosídico β (1 – 4). La galactosa tiene configuración beta, mientras que la glucosa puede ser alfa o beta. SACAROSA: Se forma por la unión de una molécula de α-glucosa con otra de β-fructosa mediante un enlace (1-2), es decir, entre los carbonos anoméricos. Es el azúcar de mesa. LOS POLISACÁRIDOS Los polisacáridos son polímeros de monosacáridos; es decir, se forman por la unión de muchas moléculas de monosacáridos mediante la formación de muchos enlaces glucosídicos y la correspondiente pérdida de moléculas de agua. Al igual que los disacáridos, estos enlaces pueden romperse mediante hidrólisis. Pueden estar constituidos por un solo tipo de monómero (homopolisacáridos), o por más de un tipo de monómero (heteropolisacáridos). Los más abundantes en los seres vivos son el almidón, el glucógeno y la celulosa, y son polímeros derivados de la misma unidad, la glucosa. ALMIDÓN: Es un polisacárido de reserva, en concreto el más importante de las plantas. Está constituido por dos polímeros de D-glucosa: amilosa y amilopectina. La amilosa está formada por una cadena de miles de restos de α-D-glucosa con enlaces glucosídicos 1-4. Esta cadena se encuentra plegada en forma de hélice. La amilopectina es más compleja. Está formada por una larga cadena como la de la amilosa pero con ramificaciones. Estas ramas son cortas y se inician mediante enlaces 1-6. La hidrólisis de este polisacárido es catalizada por la enzima α-amilasa, que lo transforma en maltosa, que finalmente pasará a glucosa. Figura 2. Estructura de molécula de amilopectina GLUCÓGENO: Es nuestra principal reserva de glucosa. En vertebrados lo encontramos principalmente en el hígado y músculos. Algunos hongos también lo tienen. Estructuralmente es parecido a la amilopectina, sólo que posee más ramificaciones y éstas son más pequeñas. Esta estructura tan ramificada hace que ésta pueda descomponerse más fácilmente que el almidón. CELULOSA: Es otro polímero de glucosa, pero con una estructura y función muy diferentes a la de los anteriores. Es una molécula que forma estructuras en las plantas, concretamente en la pared de las células vegetales. Se considera la molécula orgánica más abundante en la Tierra. Su estructura está formada por largas cadenas de β-glucosa, con enlaces 1-4. OTROS EJEMPLOS DE POLISACÁRIDOS QUITINA PECTINA HEMICELULOSA MUREÍNA ÁCIDO HIALURÓNICO CODROITINSULFATO HEPARINA GOMAS Y MUCÍLAGOS Esqueleto de artrópodos. Pared celular de las células vegetales. Compone, junto con la pectina, la matriz de la pared celular. Componente estructural de la pared bacteriana. Tejido conectivo de vertebrados, lubricante en el líquido sinovial, en articulaciones y en humor vítreo del ojo. Componente de sustancia intercelular de cartílagos y huesos. Actúa como coagulante de la sangre de los mamíferos. Secretados por las plantas, con función protectora. LÍPIDOS Los lípidos constituyen un grupo heterogéneo de sustancias cuya característica común es que son poco o nada solubles en disolventes polares como el agua, debido a que poseen numerosos enlaces apolares C-H; por otra parte son muy solubles en disolventes orgánicos como el benceno, el éter, el cloroformo… Están constituidos por carbono, hidrógeno y oxígeno. Algunos contienen además nitrógeno y fósforo. ÁCIDOS GRASOS Los ácidos grasos están formados por una larga cadena de hidrocarburos con un grupo carboxilo (-COOH). Los ácidos grasos en cuyas cadenas existen dobles enlaces se denominan insaturados, y aquellos en los que no hay dobles enlaces son saturados. En los ácidos grasos insaturados, los dobles enlaces originan una inclinación de la cadena hidrocarbonada. Los ácidos grasos saturados poseen un punto de fusión más alto que los ácidos grasos insaturados. Así, los insaturados dan lugar a acilglicéridos que son líquidos a temperatura ambiente (aceites), mientras que los saturados originan grasas que son sólidas (mantecas, tocino). Dada la naturaleza apolar de los enlaces C-H, la larga cadena hidrocarbonada de los ácidos grasos será hidrofóbica, mientras que el extremo que contiene l grupo carboxílico polar será hidrofílico. Por lo tanto, las moléculas de ácidos grasos son heteropolares. Los ácidos grasos en agua pueden formar una película superficial o pequeñas micelas. LÍPIDOS CON ÁCIDOS GRASOS ACILGLICÉRIDOS O GRASAS: Los ésteres son compuestos que se forman por la combinación de un ácido y un alcohol. El grupo carboxilo de los ácidos grasos puede reaccionar con un grupo alcohol dando lugar a un éster. Esta reacción se denomina esterificación. Los acilglicéridos son ésteres de ácidos grasos y glicerina que es un alcohol de 3 carbonos con tres grupos hidroxilo. Dependiendo del número de ácidos grasos que se esterifiquen con la glicerina, se obtienen los monoacilglicéridos, diacilglicéridos o triacilglicéridos. Los más fecuentes son los triacilglicéridos. Éstos no poseen una región polar en su molécula, por lo que se les suele denominar grasas neutras. Los acilglicéridos con ácidos grasos insaturados, son líquidos oleosos a temperatura ambiente y se denominan aceites. Las grasas (tocino o manteca) contienen ácidos grasos saturados y son sólidas a temperatura ambiente. Los aceites predominan en las plantas y las grasas sólidas en los animales. Las grasas actúan como almacenes de energía, se almacenan en las células adiposas y son utilizadas para obtener energía cuando las necesidades energéticas del cuerpo lo demanden. Cuando se degradan, las grasas pueden producir más del doble de energía que los glúcidos, propiedad que se relaciona con su alta proporción en H+. Además, las grasas sirven como aislantes térmicos. En los animales, las grasas forman una capa debajo de la piel conocida como panículo adiposo. Esta capa está especialmente desarrollada en los animales que viven en climas fríos. Las grasas también se acumulan alrededor de determinados órganos (corazón), y sirven para protegerlos contra lesiones mecánicas. Uno de los procedimientos químicos más famosos que se realizan hoy en día, en relación con los acilglicéridos y las esterificaciones es la SAPONIFICACIÓN. Se entiende por saponificación la reacción que produce la formación de jabones. La principal causa es la disociación de las grasas en un medio alcalino, separándose glicerina y ácidos grasos. Estos últimos se asocian inmediatamente con los álcalis constituyendo las sales sódicas de los ácidos grasos: el jabón. CERAS: Son ésteres de un ácido graso y un alcohol monovalente de cadena larga. Esto determina que las ceras sean sólidas y tengan puntos de fusión elevados. FOSFOLÍPIDOS: Son lípidos de composición química muy parecida a la de las grasas, es decir, glicerina más ácidos grasos. La diferencia está en que aquí uno de los tres ácidos grasos es sustituido por un grupo fosfato, el cual se une a otro compuesto químico, generalmente un aminoalcohol (Etanolamina, Colina, Serina) En los fosfolípidos se diferencian dos partes: por un lado las largas “colas” hidrofóbicas de los ácidos grasos; por otro, una “cabeza” polar, y por tanto hidrofílica, formada por el fosfato y su acompañante. Esto permite que estas sustancias formen la base estructural de las membranas celulares. Los liposomas son bicapas de fosfolípidos en forma de vesículas esféricas y se utilizan como modelo de membrana en estudios experimentales. Frecuentemente utilizado en cosmética. ESFINGOLÍPIDOS Y GLICOLÍPIDOS: Los esfingolípidos y los glicolípidos son lípidos en los que el alcohol es la esfingosina, de un alto número de carbonos, dos grupos hidroxilo en las posiciones 1 y 3, un grupo amino, y un doble enlace entre los carbonos 4 y 5. Los glicolípidos se caracterizan por la presencia, en su molécula, de monosacáridos o derivados de monosacáridos. Se diferencian en cerebrósidos o gangliósidos. Los cerebrósidos están formados por la esfingosina, que se une por su grupo amino a un ácido graso, y UN ÚNICO MONOSACÁRIDO que se une a este. En el caso de los gangliósidos, se produce la misma unión, pero esta vez, se une UN OLIGOSACÁRIDO (conjunto de monosacáridos). LÍPIDOS SIN ÁCIDOS GRASOS Estos lípidos son estructuralmente muy distintos de los que hemos estudiado hasta ahora. En este grupo se incluyen los esteroides y los terpenos. ESTEROIDES: Los esteroides son lípidos con un núcleo de 4 anillos al cual se unen diversos grupos –CH3, -OH u otros, según cada caso. El representante más conocido es el colesterol, componente de las membranas celulares, de la mielina, y sustancia clave en el metabolismo de los esteroides. Otros esteroides importantes son las hormonas sexuales, las hormonas de la corteza suprarrenal, vitamina D, ácidos biliares… TERPENOS: Los terpenos derivan sus fórmulas del isopreno, hidrocarburo de cinco carbonos. Son de cadena larga y abundan especialmente en las plantas. Son terpenos la vitamina A, E y K y los carotenoides (pigmentos fotosintéticos y precursores de la vitamina A). FUNCIONES DE LOS LÍPIDOS Acilglicéridos o grasas: Reservas energéticas en las células. Ceras: Principalmente se utilizan como material impermeable por las plantas y los animales. Fosfolípidos: Componentes de membranas. Esfingolípidos: Componentes de membranas. Esteroides: - Ácidos biliares: Componentes de la bilis. - Hormonas sexuales. - Colesterol. Componente de membranas celulares. - Vitamina D. - Hormonas de la corteza suprarrenal. Terpenos: - Esencias y aromas de los aceites esenciales de las plantas. - Giberelinas. Crecimiento de la planta. - Vitaminas A, E y K, y carotenoides. Lipoproteínas: Las membranas son estructuras de lipoproteínas. Son también las formas de transporte de lípidos en la sangre y en la linfa. Glicolípidos: Componentes de la membrana celular, DETERMINANTES ANTIGÉNICOS. También presentes en las membranas de los cloroplastos. PROTEÍNAS Las proteínas son las moléculas orgánicas más abundantes en las células. Constituyen el 50% de su peso seco. Hay muchos tipos de proteínas: enzimas, hormonas, inmunoglobulinas, proteínas de membrana… Su diversidad funcional es abrumadora, sin embargo, químicamente no son tan complejas. Las proteínas son polímeros lineales de aminoácidos unidos mediante enlaces peptídicos. Si los polímeros constan de al menos 100 aminoácidos se denominan péptidos. LOS MONÓMEROS DE LAS PROTEÍNAS: LOS AMINOÁCIDOS Los aminoácidos son las unidades químicas o bloques de construcción del cuerpo que forman las proteínas. Son variados químicamente, pero todos ellos contienen un grupo de ácido carboxílico y un grupo amino, ambos unidos al mismo átomo de carbono. Las características básicas del –NH2 y las ácidas del –COOH, hacen que los aminoácidos puedan presentarse como ácidos y como bases según el pH del medio, es decir, son anfóteros. Cada aminoácido tiene un determinado pH para el cual se comporta de forma neutra, llamada zwitterion, es decir, como un dipolo con cargas positivas en el extremo básico y cargas negativas en el extremo ácido. Este pH específico causante de la neutralidad se denomina punto isoeléctrico. Si este pH aumentase el carácter anfótero del aminoácido le haría ceder un ión H+ y actuar como un ácido equilibrando el pH. Si el pH disminuyera, se comportaría como una base. Vemos, por tanto, que estas sustancias pueden actuar como sustancias amortiguadoras en los seres vivos, evitando los peligrosos cambios de pH. ENLACES PEPTÍDICOS Dos aminoácidos se unen covalentemente entre sí, mediante la formación del llamado enlace peptídico entre el grupo ácido de uno y el amino de otro, con la correspondiente eliminación de agua. Este enlace es más rígido y corto que un enlace simple, de geometría esencialmente plana y estabilizado por resonancia. En este enlace hay libre rotación de los enlaces unidos al carbono α. El compuesto resultante es un dipéptido. Este puede seguir uniéndose con nuevos aminoácidos para formar una larga cadena que se denomina polipéptido. En cualquier polipéptido el grupo amino se escribe a la izquierda y el grupo carboxilo a la derecha. ESTRUCTURA DE LAS PROTEÍNAS La variedad de las proteínas es potencialmente ilimitada, diversidad que se sustenta en los diferentes niveles estructurales que se definen en estas moléculas. Esta diversidad permite a las proteínas desempeñar una gran cantidad de funciones estructurales y metabólicas. Se han descrito hasta cuatro niveles de organización en las proteínas: Estructura primaria Es la secuencia de aminoácidos de la proteína. Nos indica qué aminoácidos componen una determinada cadena peptídica. Estructura secundaria Es la disposición de la secuencia de aminoácidos en el espacio. Gracias a la capacidad de giro de sus enlaces, presentan una gran estabilidad. Esta estructura se nos presenta de dos maneras: - Hélice plegada (α-hélice): En forma helicoidal. Conformación β: Forma una cadena de zigzag denominada lámina de disposición plegada. Estructura terciaria Informa sobre la disposición de la estructura secundaria al plegarse sobre sí misma originando así una conformación globular. Favorece la propiedad de la solubilidad en el agua. Estructura cuaternaria Informa de la unión mediante enlaces débiles (no covalentes) de varias cadenas polipeptídicas con estructura terciaria. El ejemplo más claro es la hemoglobina. PROPIEDADES DE LAS PROTEÍNAS SOLUBILIDAD Las proteínas son solubles en agua cuando adoptan una conformación globular. DESNATURALIZACIÓN La desnaturalización de una proteína consiste en la ruptura de los enlaces que mantenían la estructura cuaternaria, terciaria y secundaria, manteniéndose así sólo la primaria. Se da con temperaturas superiores a los 50/60ºC o inferiores a los 10/15ºC. Puede ser tanto reversible como irreversible, depende de los tipos de desnaturalización. ESPECIFICIDAD Cada especie de seres vivos es capaz de crear sus propias proteínas. Esto explica algunos fenómenos biológicos como la compatibilidad o incompatibilidad de trasplantes de órganos. FUNCIONES DE LAS PROTEÍNAS - Función defensiva (Creación de anticuerpos). Función enzimática (Creación de enzimas). Función homeostática (Mantenimiento del pH constante). Función estructural (Creación de tejidos y estructuras). Función de transporte. Funciones contráctiles. ENZIMAS Las enzimas son proteínas, las más abundantes, que catalizan reacciones químicas específicas en los seres vivos. Son proteínas globulares formadas por cadenas que se pliegan, formando un surco denominado centro activo, lugar donde se producen las reacciones químicas. MECANISMO DE ACCIÓN DE LAS ENZIMAS. CATÁLISIS ENZIMÁTICA. En toda reacción química se produce la transformación de unas moléculas iniciales, denominadas sustratos (S) en unas sustancias finales o productos (P). Esta transformación requiere un aporte inicial de energía. La energía añadida incrementa la energía cinética de las moléculas que reaccionan, permitiendo que un mayor número de ellas choquen con suficiente fuerza para superar la repulsión. La energía que deben poseer las moléculas para iniciar la reacción se conoce como energía de activación. Las enzimas son catalizadoras de reacciones bioquímicas. Gracias a las enzimas, las células son capaces de desarrollar reacciones químicas a gran velocidad y a temperaturas realmente bajas. E+S ES P+E La enzima actúa sobre el sustrato; el sustrato se une a la región determinada de la enzima, llamada centro activo. Una vez formados los productos, la enzima puede cometer un nuevo ciclo de reacción. El centro activo es una región de la enzima donde se une el sustrato. Se distinguen dos zonas: A) Sitio de unión: Formado por los aminoácidos en contacto con el sustrato. B) Sitio Catalítico: Lugar donde se produce la reacción. Las enzimas aceleran la reacción sin alterar el equilibrio. La catálisis hace disminuir el tiempo necesario para alcanzar el equilibrio. TIPOS DE ENZIMAS - OXIDORREDUCTASAS: Se encuentran en reacciones redox. TRANSFERASAS: Se encuentran en reacciones de transferencia de grupos funcionales. HIDROLASAS: Se encuentran en reacciones de hidrólisis. LIASAS: Se dan en reacciones de adición de dobles enlaces. ISOMERASAS: Se dan en reacciones de isomerización. LIGASAS: Se dan en reacciones de unión de una o más moléculas. ESPECIFICIDAD DE LAS ENZIMAS Cada enzima sólo cataliza un tipo de reacción, y casi siempre actúa sobre un único sustrato. Esta especificidad se debe a la complementariedad que debe existir entre el sustrato y el centro activo de la enzima. (CONCEPTO LLAVE-CERRADURA). COFACTORES Y COENZIMAS Son pequeñas moléculas NO PROTEÍNICAS que transportan grupos químicos entre enzimas. Se denomina holoenzima, o encima activa, al conjunto de la coenzima y de la enzima inactiva o apoenzima. Las vitaminas son sustancias orgánicas de composición variable imprescindibles para el normal funcionamiento del metabolismo. Son coenzimas o componentes de coenzimas. El hombre no puede sintetizar las vitaminas y debe tomarlas en la dieta como tales o como sustancias transformables en vitaminas, llamadas provitaminas. REGULACIÓN DE LA ACTIVIDAD ENZIMÁTICA Los seres vivos tienen varias formas de regular la actividad enzimática. Efectos de la Temperatura A medida que la Tª aumente, la enzima verá incrementada su actividad hasta el momento de su desnaturalización. Efectos del pH Si el pH del medio se aleja del óptimo de la enzima, ésta verá modificada su concentración de protones, lo que modificará la actividad enzimática, hasta hacerse no funcional. Efectos de la concentración [ ] Si se mantiene constante la concentración de la enzima, cuando la concentración del sustrato aumente, la velocidad de reacción se verá incrementada. INTERACCIONES ALOSTÉRICAS La interacción alostérica es un mecanismo por el cual una enzima puede activarse o inactivarse temporalmente. Estas interacciones ocurren en enzimas que tienen al menos dos sitios de unión, uno, el conocido como centro activo, y el otro, conocido como centro regulador, al que se une una molécula conocida como efector alostérico. Las interacciones alostéricas suelen localizarse en punto estratégicos de las vías enzimáticas, en los primeros pasos o en los puntos de ramificación. Unas veces el propio sustrato actúa como efector activador induciendo cambios en la conformación del centro activo que favorecen la catálisis enzimática. Otras veces, es el producto final el que actúa como efector alostérico inhibiendo la función de una de las enzimas; este proceso se denomina inhibición por retroalimentación o inhibición feed-back. INHIBICIÓN Los inhibidores son sustancias que disminuyen o incluso anulan la velocidad de las reacciones catalizadas por enzimas. Puede ser de distintos tipos: - - Inhibición reversible: Unión no covalente de un inhibidor a una enzima. A) Inhibición competitiva: Unión del inhibidor al centro del sistema compitiendo con el sustrato. B) Inhibición no competitiva: Unión del inhibidor a una parte de la enzima distinta al centro activo. Inhibición irreversible: Unión covalente de un inhibidor a una enzima. Inhibición competitiva Inhibición no competitiva ÁCIDOS NUCLEICOS Los ácidos nucleicos son esenciales para la vida. Ellos constituyen el material genético de todos los seres vivos, es decir, la información para la síntesis de proteínas. Los ácidos nucleicos, ácido desoxirribonucleico (DNA) y ácido ribonucleico (RNA), son polímeros de nucleótidos. LOS MONÓMEROS DE LOS ÁCIDOS NUCLEICOS: LOS NUCLEÓTIDOS Un nucleótido está formado por la unión de tres moléculas: un monosacárido de 5 carbonos (pentosa), una base nitrogenada y el ácido fosfórico. La pentosa puede ser de dos tipos: ribosa o 2-desoxirribosa. En el RNA hay ribosa (ribonucleótidos) y en el DNA hay desoxirribosa (desoxirribonucleótidos). Las bases nitrogenadas son moléculas cíclicas, derivadas de dos anillos básicos: la purina y la pirimidina. Las bases nitrogenadas que se encuentran más frecuentemente en el DNA y RNA son la adenina y guanina (derivadas de la purina), y la citosina, la timina y el uracilo (derivadas de la pirimidina). De éstas, la adenina, citosina y guanina se encuentran tanto en el DNA como en el RNA, mientras que la timina es exclusiva del DNA, y el uracilo es característico del RNA. En la formación de los nucleótidos, la pentosa y la base nitrogenada se unen mediante un enlace N-glucosídico entre el carbono 1´ de la pentosa y el nitrógeno 1 de la base nitrogenada en las bases pirimidínicas, y el nitrógeno 9 en las púricas. Este compuesto se llama nucleósido; es decir, es el producto de la condensación entre el hidroxilo (-OH) de la posición 1´ del azúcar (pentosa), y un nitrógeno de una base mediante un enlace N-glucosídico. Finalmente, el ácido fosfórico (H3PO4) se une a la pentosa por el carbono 5´mediante un enlace éster, formándose el nucleótido. POLINUCLEÓTIDOS Dos nucleótidos pueden unirse entre sí mediante enlaces fosfodiéster entre el carbono 5´ de un nucleótido y el carbono 3´del otro. Las moléculas de DNA son polinucleótidos constituidos por cadenas de desoxirribonucleótidos. Las moléculas de RNA son polinucleótidos constituidos por unidades de ribonucleótidos. ÁCIDO DESOXIRRIBONUCLEICO (DNA) En las células eucariotas el DNA se encuentra localizado esencialmente en el núcleo, aunque también se encuentra en las mitocondrias y en los cloroplastos. En las células procariotas el DNA se encuentra en el citoplasma. En las células procariotas que poseen un único cromosoma, todo el DNA se halla esencialmente en forma de una única macromolécula, mientras que en las células eucariotas que contienen varios cromosomas, existen varias macromoléculas de DNA. Estructura primaria del DNA La estructura primaria está determinada por la secuencia de desoxirribonucleótidos. La diferencia entre cada cadena vendrá dada por la secuencia de bases nitrogenadas a lo largo del esqueleto de DNA. En dicha secuencia reside la información para la síntesis proteica. Estructura secundaria del DNA (Modelo de doble hélice de Watson y Crick) Esta estructura se basa en el modelo para la estructura tridimensional del DNA propuesta por Watson y Crick. Obtuvieron este modelo basándose en los datos experimentales obtenidos por difracción de rayos x. Los estudios acerca de la composición en bases del DNA de diferentes organismos indicaban que la cantidad de adenina es igual a la de timina, y que la de guanina es igual a la de citosina (A = T; G = C). Los datos obtenidos por la difracción por rayos x mostraban patrones que reflejaban los giros de una hélice. A partir de estos datos intentaron crear un modelo estructural de DNA que concordara con los datos obtenidos, y que pudiera indicar el método mediante el cual se pudiera replicar la información genética. - - - La molécula de DNA está formada por dos cadenas polinucleótidas. Cada cadena forma una hélice dexrógira. Las cadenas discurren en dirección opuesta; es decir una normal 5´- 3´, y otra boca abajo 3´- 5´. Son antiparalelas. Los ejes de pentosa – fosfato de las cadenas se sitúan periféricamente, mientras que las bases se disponen perpendicularmente hacia el interior. La doble hélice tiene siempre el mismo grosor, por lo que el emparejamiento obligado es siempre entre una purina y una pirimidina. Este apareamiento permite la formación del máximo número de enlaces de hidrógeno (tres para el par C-G y dos para el A-T). El orden de las bases de una cadena determina el de la otra, es decir, que son complementarias. La estructura de la doble hélice queda estabilizada por puentes de hidrógeno. ÁCIDO RIBONUCLEICO (RNA) El RNA es el ácido nucleico más abundante en las células. Las moléculas de RNA son polímeros formados por la unión de ribonucleótidos de adenina, guanina, citosina, uracilo, y en menos proporción otras bases nitrogenadas como el pseudouracilo. Los RNA, excepto en algunos casos, son monocatenarios y suelen tener únicamente estructura primaria. Esta estructura viene definida por la secuencia de bases a lo largo de la cadena de RNA. En algunos casos puede presentar regiones de apareamiento complementarias en la cadena, capaces de formar una doble hélice. Tipos de RNA Existen tres tipos diferentes de RNA: RNA mensajero, RNA ribosómico, y RNA de transferencia. RNA mensajero - - Contiene la información procedente del DNA. Su función es transportar la información copiada desde el núcleo hasta los ribosomas, donde actúa como patrón para la ordenación secuencial durante la síntesis de proteínas. Los codones son tripletes de nucleótidos que especifican la secuencia de aminoácidos de la cadena polipeptídica. RNA ribosómico - Son moléculas lineales de una sola cadena polinucleotídica con algunas bases metiladas y que pueden presentar numerosas zonas de emparejamiento. Existen tres tipos de RNAr en las células procariotas y cuatro tipos en las células eucariotas. RNA de transferencia - Se encuentra en el citoplasma en forma de moléculas dispersas. Su función es transportar aminoácidos específicos hasta los ribosomas, donde se sintetizan las proteínas. Todos los RNAt comparten la presencia de guanina en el extremo 5´ terminal, y en su extremo 3´, donde se enlaza el aminoácido, la secuencia C-C-A. Algunas regiones del RNAt presentan estructuras secundarias, constituidas por una doble hélice gracias a la disponibilidad de apareamiento en algunas regiones. Presentan un anticodón, que consiste en un triplete de nucleótidos que actúa como secuencia complementaria de bases que porta el RNAm como codón característico. - Presenta estructura terciaria. La molécula se encuentra plegada en forma de L con el brazo anticodón en un extremo y el brazo aminoácido en el otro. También conocida como estructura en forma de hoja de trébol. LA CÉLULA Distinguimos dos tipos de células: - CELULAS EUCARIOTAS: Se llama célula eucariota a todas las células con un núcleo celular delimitado dentro de una doble capa lipídica, la envoltura nuclear, la cual es porosa y contiene su material hereditario, fundamentalmente su información genética. Existen dos tipos de células eucariotas: A) CÉLULAS ANIMALES B) CÉLULAS VEGETALES - CÉLULAS PROCARIOTAS: Se llama procariota a las células sin núcleo celular definido, es decir, cuyo material genético se encuentra disperso en el citoplasma, reunido en una zona denominada nucleoide. LA CÉLULA EUCARIOTA ORGÁNULOS PRESENTES EN ESTAS CÉLULAS 1. MEMBRANA PLASMÁTICA - Se trata de una barrera intercambiadora de materia entre la célula y el exterior. Es una estructura bastante fluida y cambiante. Presenta permeabilidad variable para líquidos y otras sustancias. Interviene en actividades enzimáticas, fijación de virus… Grosor de aproximadamente unos 7-9 nm. Presenta estructura trilaminar [dos capas osmiófilas (oscuras) y una capa osmiófoba (clara)]. Composición proteínica y lipídica, con pequeña cantidad de glúcidos. Fácilmente atravesada por sustancias liposolubles. Se utiliza el llamado MODELO DE MOSAICO FLUIDO, para explicar la disposición de los componentes moleculares en la membrana: A) Proteínas intrínsecas fuertemente ligadas a la membrana. B) Proteínas extrínsecas débilmente ligadas a la membrana. C) Una doble capa de lípidos. D) Cadenas de carbohidratos unidas a las proteínas intrínsecas. El transporte de sustancias a través de esta bicapa se produce de 4 maneras diferentes: - Difusión. Ósmosis. Transporte activo. Transporte por vesículas. DIFUSIÓN Las moléculas simples se desplazan desde la región de mayor concentración a la de menor concentración. Algunas proteínas contribuyen a este proceso, lo que se denomina difusión facilitada. ÓSMOSIS Consiste en el paso del agua, a través de una membrana selectivamente permeable, de la menos concentrada a la más concentrada. TRANSPORTE ACTIVO Se trata de un método de trabajo en contra de gradiente en el que se realiza un gran gasto energético y es requerida la implicación de ciertas proteínas. Hay dos tipos de este transporte: - Transporte activo primario (bomba de sodio). Cotransporte (simporte) (bomba de sodio/potasio). Figura 1. Transporte activo y transporte pasivo. Figura 2. Transporte activo primario y cotransporte. TRANSPORTE POR VESÍCULAS En este caso las sustancias no atraviesan la membrana propiamente dichas, sino que salen o entran de ésta en el interior de vesículas, que se forman a partir de trozos de membrana. Lo hacen por medio de la fagocitosis, pinocitosis y endocitosis mediada por receptores. DIFERENCIACIONES DE LA MEMBRANA - Microvellosidades: Expansiones citoplasmáticas cuya función es aumentar el poder de absorción. Estructuras de unión entre las células: Algunas células se encuentran unidas por diferentes estructuras, así como desosomas, uniones estrechas y hendiduras. 2. ENVOLTURA Y PARED CELULAR En la mayoría de las células eucariotas hay una especie de capa difusa en la cara externa de la membrana plasmática, que ha recibido diversos nombres, envoltura celular, glucocálix… La pared celular es una gruesa membrana de secreción, elaborada por la célula, propia de las células vegetales. Se pueden distinguir tres capas en ella: - Lámina media: Sustancia intercelular que mantiene unidas las células. Entre sus componentes predominan los pectatos. Pared primaria: Se desarrolla en las células jóvenes y está compuesta por celulosa, pectatos, hemicelulosa y otros polisacáridos. Pared secundaria: Se deposita en las células que han dejado de crecer, entre la membrana plasmática y la pared primaria en capas sucesivas. Está compuesta por celulosa y otros polisacáridos. 3. EL CITOPLASMA (citosol y citoesqueleto) Es uno de los tres conjuntos de estructuras celulares que está formado por una gran variedad de estructuras bien definidas. Citosol También llamado hialoplasma, es una disolución coloidal que puede experimentar cambios entre una fase viscosa y otra fase fluida. Se ha descubierto en su seno toda una red de estructuras tubulares y filamentosas que constituyen el llamado citoesqueleto. Como función cabe destacar que el hialoplasma es el lugar de la mayoría de las reacciones metabólicas de la célula. Citoesqueleto Tiene una estructura muy compleja y cambiante. Se han identificado al menos tres tipos de componentes: microtúbulos, filamentos de actina y filamentos intermedios. Los microtúbulos son tubos cilíndricos formados por tubulina, que forman parte del esqueleto celular. Los filamentos de actina están formados por moléculas de actina y tienen que ver con la motilidad celular. Finalmente, los filamentos intermedios contienen proteínas fibrosas y se les atribuyen funciones esqueléticas y de movimiento celular. 4. CENTRÍOLOS Son orgánulos que suelen encontrarse por pares en la mayoría de las células animales y algunas de las vegetales. Tienen forma de cilindro formado por 9 tripletes de microtúbulos. Desempeñan una importante función en la división celular. Relacionados con ellos se encuentran los cuerpos basales y los kinetosomas, que se encuentran en la base de cilios y flagelos. 5. CILIOS Y FLAGELOS Son estructuras finas que sobresalen, a modo de pelillos, de la superficie de muchas células eucariotas. Se diferencian por la longitud, los cilios son cortos, y los flagelos largos. Están presentes en muchos organismos unicelulares móviles y en algunos tejidos de pluricelulares. Su estructura presenta 9 dobletes de microtúbulos. Están implicados en el movimiento celular y la captura de alimento. 6. RETÍCULO ENDOPLASMÁTICO El retículo es un sistema de sáculos limitados por membranas de naturaleza lipoproteíca distribuido por el citoplasma. Está presente en todas las eucariotas. Se distinguen dos regiones en este orgánulo: retículo endoplasmático rugoso y retículo endoplasmático liso. Retículo Endoplasmático Rugoso (RER) Constituido por cisternas y túbulos que llevan ribosomas adheridos. En este orgánulo se puede distinguir una membrana, una cavidad interna y los ribosomas. Su función es el almacén y transporte de las proteínas sintetizadas por los ribosomas de su superficie. La cadena polipeptídica es llevada hasta el retículo endoplasmático rugoso por los primeros aminoácidos de la cadena, la llamada secuencia señal. De aquí podrá pasar al llamado retículo de transición y finalmente al complejo de Golgi, desde donde será secretada o enviada a otro orgánulo. Retículo Endoplasmático Liso (REL) Constituido por un laberinto de túbulos interconectados y vesículas. Los túbulos del retículo endoplasmático liso no tienen ribosomas adheridos a sus membranas. En ocasiones se observan conexiones entre los elementos del retículo endoplasmático rugoso y los del liso. Este tipo de retículo puede asociarse a diversas funciones, según el tipo de célula: - En el hígado está relacionado con la síntesis de glucógeno e interviene en procesos de desintoxicación. En las células que sintetizan esteroides, interviene en la síntesis y transporte de lípidos. En el intestino interviene en la síntesis de triglicéridos. 7. RIBOSOMAS Son los orgánulos celulares más numerosos y universales, encontrándose en todo tipo de células. Son pequeñas partículas NO MEMBRANOSAS constituidas por dos subunidades unidas. Están compuestos de RNA ribosómico y proteínas. Intervienen en la síntesis de proteínas. En muchos casos, varios ribosomas aparecen en la superficie del retículo unidos por una molécula de RNAm, constituyendo los llamados polirribosomas. 8. EL APARATO DE GOLGI El aparato de Golgi está constituido por dos elementos: cisternas aplanadas y vesículas. Las cisternas, con aspecto de sacos aplanados tienden a agruparse en un número variable constituyendo los llamados dictiosomas. Rodeando al dictiosoma, existen vesículas redondeadas. En la cara interior cóncava se originan vesículas cargadas de material a partir de las cisternas. A la cara exterior convexa llegan vesículas procedentes del retículo endoplasmático rugoso, más concretamente el de transición, portando el material elaborado en el retículo. El aparato de Golgi tiene un importante papel en procesos de transporte y de secreción, así como: - Secreción de proteínas y glicoproteínas. Formación de Lisosomas. Secreción de la pared celular de las células vegetales. Formación de nuevas membranas. 9. LISOSOMAS Los lisosomas son pequeñas vesículas limitadas por una membrana, que contienen enzimas destinadas a la digestión intracelular. Se localizan en todas las eucariotas y se suelen distinguir dos tipos: lisosomas primarios, que todavía no han intervenido en fenómenos de degradación, y lisosomas secundarios, que se originan al unirse lisosomas primarios a vacuolas fagocitarias. 10. MITOCONDRIAS Las mitocondrias son orgánulos presentes en todas las células eucariotas tanto animales como vegetales. Pueden ser esféricas, alargadas o con aspecto filamentoso. Están rodeadas por dos membranas, separadas por un espacio intermembranal. En el interior se encuentra la matriz. De las membranas, la más interesante es la interna, la cual presenta unos pliegues o crestas. La mitocondria es la sede de la mayor parte de la respiración celular. Su función es producir energía, que será almacenada en forma de ATP. En la matriz abundan enzimas de otras fases respiratorias como el Ciclo de Krebs, la βoxidación de los ácidos grasos… Cabe destacar también la presencia de DNA, ribosomas y RNA de tipo bacteriano. 11. CLOROPLASTOS En las células vegetales y en algunos protozoos suele encontrarse un grupo de orgánulos, los plastos, caracterizados por almacenar diversas clases de pigmentos u otras sustancias. Los cloroplastos son un tipo de plastos que contienen pigmentos fotosintetizadores, especialmente la clorofila. Son estructuras que se encuentran preferentemente en los órganos verdes de las plantas. Tienen dos membranas y un espacio intermembranal, pero la membrana interna no presenta crestas. Además hay un tercer sistema membranoso formado por los tilacoides. Éstos son pequeños sacos aplanados que se presentan apilados formando la grana. Dentro de los tilacoides se encuentran la clorofila, carotenoides… En el interior del plasto está el estroma, análogo de la matriz de la mitocondria (medio interno del plasto), en el que se encuentran, entre otros, DNA propio y pequeños ribosomas. 12. EL NÚCLEO CELULAR El núcleo se encuentra en todas las células eucarióticas. Es el mayor orgánulo de la célula. Presenta dos aspectos diferentes según el momento de su vida en el que se encuentra. Es como si se sucediesen dos núcleos a lo largo de la vida celular: cuando se está dividiendo se observa el llamado núcleo mitótico y el resto de su vida, el núcleo interfásico. Núcleo interfásico Aparece rodeado por dos membranas. La externa es una prolongación del retículo endoplasmático rugoso, y la interna presenta en su cara interna unas proteínas especiales. Entre ambas se encuentra un espacio intermembranoso. Esta doble membrana es atravesada por poros. El interior está ocupado por una matriz gelatinosa, el nucleoplasma. En él se sitúa el nucléolo, estructura esférica formada por DNA, RNAr y enzimas, y sobre todo la cromatina. Suelen distinguirse dos tipos de cromatina, la eucromatina, de estructura poco densa y formada por DNA genéticamente activo, y la heterocromatina, visible al microscopio óptico, con el DNA muy empaquetado y activo. Núcleo mitótico. Los cromosomas. Durante la mitosis, núcleo cambia radicalmente de aspecto. La cromatina se hace mucho más densa y da lugar a estructuras denominadas cromosomas. El resto de los orgánulos del núcleo, membrana y nucléolo desaparecen durante la profase para volver a formarse en la telofase. Características externas de los cromosomas. Los cromosomas metafísicos aparecen formados por dos mitades genéticamente idénticas, las cromátidas. Cada cromátida presenta dos partes a modo de brazos, separadas por un estrechamiento o constricción primaria, donde se localiza el centrómero. Esta estructura mantiene unidas las dos cromátidas del cromosoma metafásico. En algunos cromosomas, pueden aparecer otras constricciones secundarias que separan fragmentos de cromosoma denominados satélites. Los cromosomas presentan formas muy variadas. Se distinguen varios tipos: metacéntricos, con brazos iguales, acrocéntricos, con brazos desiguales, y telocéntricos, con un solo brazo. El tamaño también es muy variable. Seguramente la característica externa más importante es el número de cromosomas. Este número es constante en todas las células de una especie, excepción hecha de los gametos. Lo más frecuente es que las células normales de un organismo tengan dos juegos de n cromosomas cada uno, repetidos en cuanto a su aspecto, es decir, tienen 2n cromosomas (el llamado número diploide), mientras que los gametos tienen siempre un solo juego de n cromosomas, o número haploide. En consecuencia, en cada célula diploide encontraremos n parejas, estando formada cada una por dos cromosomas, denominados homólogos, aparentemente iguales. Estructura de los cromosomas Los cromosomas de eucariotas están formados por DNA unido a unas proteínas básicas, principalmente histonas (proteínas tetraméricas). El complejo formado en esta unión es la cromatina, unidas estructural básica de los cromosomas y principal componente nuclear. La cromatina puede presentar aspectos diferentes durante la vida celular porque tiene una gran capacidad de organizarse a diferentes niveles. En el nivel más sencillo, la cromatina aparece constituida por una doble hélice de DNA extendida que, de vez en cuando, se enrolla alrededor de una “carrete” de 8 histonas originándose una estructura que recuerda a un rosario o collar de perlas. Estos carretes se denominan nucleosomas. En el siguiente nivel, estos nucleosomas vuelven a enrollarse entre sí formando una fibra más gruesa, el llamado nivel de empaquetamiento de la cromatina interfásica. En posteriores condensaciones, estas fibras forman bucles, los cuales se vuelven a enrollar dando lugar al conocido cromosoma metafásico. Funciones del núcleo El núcleo, más en concreto, la cromatina, realiza dos funciones fundamentales: dirige la vida celular gracias a la información genética contenida en su DNA y duplica este material genético y lo transmite a las células hijas en cada división. CÉLULA PROCARIOTA Composición de la célula procariota - Cápsula de polisacáridos, lugar donde aparecen algunas bacterias. Membrana fotosintética (sólo en bacterias fotosintetizadoras). Plásmido (Pequeños fragmentos extras de DNA. Puede haber varios. Gran importancia en genética). Flagelo (puede haber varios). Mesosoma (pliegues hacia dentro de la membrana). Pared Celular (compuesta por diversos polímeros). Membrana celular. Ribosomas. Citoplasma. DNA circular (aunque también se le define como cromosoma, es muy diferente del eucariótico: sin histonas y sin empaquetamiento). METABOLISMO El metabolismo es el conjunto de reacciones que tienen lugar en las células, catalizadas por enzimas, destinadas a la transformación de diferentes compuestos para obtener energía química, ya sea de la materia orgánica o de la luz solar, convertir los nutrientes exógenos en los elementos de las macromoléculas propias de la célula, y formar y degradar biomoléculas necesarias para las funciones celulares. Se compone de dos rutas: catabolismo y metabolismo. CATABOLISMO Es la ruta metabólica de degradación oxidativa de las moléculas orgánicas, cuya finalidad es la obtención de energía, necesaria para que la célula pueda desarrollar sus funciones vitales. Este proceso que ocurre en tres etapas: 1) Las macromoléculas poliméricas se hidrolizan en sus monómeros; es decir, los polisacáridos en monosacáridos, las grasas en ácidos grasos y glicerina, y las proteínas en aminoácidos. Esta fase, la llamada digestión, se produce, sobre todo, fuera de las células. 2) Los monómeros penetran en el citosol de las células y son degradadas a un número muy reducido de compuestos sencillos. El producto fundamental de las reacciones de esta fase es la acetil CoA. 3) Esta última etapa se desarrolla en las mitocondrias, la acetil CoA se oxida para dar CO 2, H2O y la energía liberada, que se conserva en forma de ATP. Esta etapa está integrada por dos rutas catabólicas: el Ciclo de Krebs y la Fosforilación Oxidativa. GLUCÓLISIS Se trata de una vía metabólica encargada de oxidar la glucosa con la finalidad de obtener energía para la célula. Consiste en diez reacciones enzimáticas que convierten la glucosa en dos moléculas de piruvato; el cual es capaz de seguir otras vías metabólicas y así continuar entregando energía al organismo. Visión General - - Durante la glucólisis se obtiene un rendimiento neto de: A) 2 moléculas de ATP que sirven como fuente de energía. B) 2 moléculas de NADH que tienen diferentes destinos, actuando como poder reductor. La glucólisis es la primera ruta metabólica a la que se recurre. Se encuentra dividida en dos fases: A) FASE 1: También llamada etapa preparativa, es una etapa degradativa, no es oxidativa, y se consumen 2 moléculas de ATP por cada glucosa. B) FASE 2: También conocida como etapa oxidativa. Se oxida el NAD + para dar NADH + H+ y se forman 4 moléculas de ATP por transferencia de grupos fosfato al ADP. Funciones Las funciones de la glucólisis son: - Generación de moléculas de alta energía (ATP y NADH), utilizados en la respiración aeróbica y en las llamadas fermentaciones. Generación de piruvato, que pasará al ciclo de Krebs como parte de la respiración aeróbica. Producción de intermediarios de 6 y 3 carbonos que pueden ser utilizados en otros procesos celulares. Balance Entradas 1 molécula de ATP 1 molécula de ATP NAD+ 1 molécula de Glucosa Salidas 2 moléculas de ATP 2 moléculas de ATP 2 NADH + H+ 2 moléculas de Piruvato Balance NETO: 2 moléculas de ATP, 2 NADH + H+ y 2 moléculas de Piruvato FERMENTACIONES La fermentación es un proceso catabólico mediante el cual se oxida materia rica en glúcidos produciendo moléculas más pequeñas y generando energía para el organismo que la realiza. El piruvato permanece en el citosol y reacciona con los átomos de hidrógeno resultantes de la deshidrogenación de las moléculas de glucosa que han sido captadas por la coenzima celular NAD+. Esta reacción completa el proceso anaeróbico al permitir la regeneración de NADH + H + y posibilitar que prosiga el proceso de oxidación de las moléculas de glucosa. Se pueden distinguir varios tipos de fermentaciones: A) FERMENTACIÓN ALCOHÓLICA: Proceso por el cual las levaduras tranforman el piruvato en etanol y CO2, moléculas que se difunden al exterior. B) FERMENTACIÓN LÁCTICA: Proceso que consiste en la reducción del piruvato a lactato. Es característico de bacterias y de las células del músculo estriado al ser sometido a condiciones extremas de trabajo en las que el aporte de O2 a la célula se ve reducido. RESPIRACIÓN CELULAR La tercera fase del catabolismo de la glucosa consiste en la oxidación total de la molécula hasta la obtención de los compuestos de carbono e hidrógeno energéticamente más estables (CO2 y H2O). La respiración celular está constituida por dos procesos: el Ciclo de Krebs y la Fosforilación Oxidativa. La serie de reacciones características de estos procesos se producen en las mitocondrias por lo que el proceso se inicia con el paso de piruvato al interior de las mitocondrias. CICLO DE KREBS También llamado ciclo del ácido cítrico o de los ácidos tricarboxílicos, es una ruta metabólica que consiste en un conjunto de reacciones por las cuales se consigue energía y la oxidación completa de la molécula de acetilo a CO2. Es un proceso que comienza con la oxidación y degradación del piruvato a acetilo. Posteriormente, las reacciones de oxidación del acetilo forman un ciclo al unirse éste a una molécula de 4 carbonos (oxalacetato), el cual se regenera en un proceso que se desarrolla en 8 etapas. Balance Mediante este proceso obtenemos 3 NADH + H+, 1 FADH2 y 1 molécula de GTP. Sin embargo, en la glucólisis, por cada molécula de glucosa obtenemos 2 moléculas de piruvato, por lo que estos resultados se duplican, obteniendo así 6 NADH + H+, 2 FADH2 y 2 moléculas de GTP. FOSFORILACIÓN OXIDATIVA Es un proceso por el que se genera ATP cuando, mediante una serie de transportadores de electrones, se transfieren electrones desde el NADH o FADH2 al O2. Se lleva a cabo por sistemas respiratorios que se localizan en la membrana interna de las mitocondrias. La oxidación del NADH produce 3 moléculas de ATP mientras que la oxidación del FADH2 genera 2 moléculas de ATP. La oxidación y la fosforilación son dos procesos acoplados. Podemos dividirlo en dos fases: - - FASE I: Consiste en el paso de electrones del NADH y FADH2 a la cadena transportadora de electrones. Las reacciones liberan energía. Esta energía liberada la usan las proteínas para aceptar los electrones y empujar los protones hacia fuera. Se produce un gradiente electroquímico de protones que se almacenan en el espacio intermembranal. Los electrones de baja energía se transfieren a un aceptor final, el oxígeno molecular (O2), que se reduce hasta agua. FASE II: Consiste en la síntesis de ATP a partir de ADP y grupos fosfato. El movimiento controlado de protones de regreso a la matriz mitocondrial mediante la enzima ATP sintetasa proporciona la energía necesaria para fosforilar ADP en ATP. OTRAS VÍAS CATABÓLICAS Después de la ingesta de alimento, la mayor parte de acetil CoA procede de la glucosa derivada de los alimentos, pero las células también pueden obtener energía a partir de las grasas o las proteínas. La oxidación de estas moléculas sigue en principio vías metabólicas particulares, para incorporarse posteriormente a la vía principal, el Ciclo de Krebs. Catabolismo de las grasas La hidrólisis de las grasas en el sistema digestivo origina ácidos grasos y glicerina. Los ácidos grasos se oxidan en la matriz mitocondrial mediante un proceso conocido como β-oxidación. La β-oxidación es una ruta catabólica espiral en la que cada vez que se repite una secuencia de cuatro reacciones (1 vuelta de espiral) la cadena del ácido graso se acorta dos átomos de carbono, que salen en forma de acetil CoA. Previo a este proceso cíclico de reacción, la molécula de ácido graso es activada formando acilCoA, que pasa a la mitocondria, donde se activa el proceso cíclico que consiste en dos oxidaciones, una hidratación y una lisis de la molécula, originándose así Acetil CoA y un nuevo acil CoA con dos átomos menos de carbono. Posteriormente, el Acetil CoA se incorpora al Ciclo de Krebs hasta su oxidación total. Los ácidos grasos producen una fuente energética más rica que la misma cantidad de glúcidos o proteínas. Catabolismo de las proteínas Una vez descompuestas en los aminoácidos constituyentes, éstos se incorporan por dos procesos distintos a las vías catabólicas principales: - Transaminación Desaminación. Catabolismo de los ácidos nucleicos Los ácidos nucleicos no son moléculas energéticas para la célula. De su catabolismo se obtienen una serie de sustancias que puede aprovechar la célula, o que, de lo contrario, tendrá que excretar. ANABOLISMO Se trata de una fase metabólica en la que a partir de unos pocos precursores sencillos y relativamente oxidados se obtienen moléculas orgánicas más complejas y reducidas. Podemos distinguir dos tipos de anabolismo: - ANABOLISMO AUTÓTROFO: Consiste en la síntesis de moléculas orgánicas sencillas a partir de nutrientes inorgánicos como CO2, NO3-, SO42-… ANABOLISMO HETERÓTROFO: Consiste en la síntesis de moléculas orgánicas más complejas a partir de nutrientes orgánicos más sencillos. VÍAS ANABÓLICAS EN LA CÉLULA HETERÓTROFA La célula heterótrofa no es capaz de sintetizar sus nutrientes orgánicos a partir de materia inorgánica, por ello utiliza moléculas simples provenientes de la degradación por vías catabólicas de otras moléculas más complejas para construir sus macromoléculas. Las vías anabólicas siguen, en general, reacciones comunes a las vías catabólicas, diferenciadas únicamente en unas pocas reacciones estratégicas. El hecho de que una misma reacción se utilice para la síntesis y degradación supone un ahorro para la célula al utilizar prácticamente las mismas enzimas. Síntesis de Glúcidos La regulación de la concentración de glucosa es uno de los aspectos importantes del metabolismo celular. Esta regulación se realiza mediante dos procesos: - - Glucogenogénesis: Ruta anabólica por la que tiene lugar la síntesis de glucógeno a partir de un precursor más simple. Se lleva a cabo principalmente en el hígado y en menor medida en los músculos. Glucogenolisis: Es un proceso que se lleva a cabo cuando el organismo requiere un aumento de glucosa, y consiste en la degradación de glucógeno. Sin embargo, hay situaciones en las que es insuficiente el aporte de glucosa y el organismo debe sintetizar glucosa a partir de metabolitos no glucídicos. A este proceso se le denomina gluconeogénesis, y es una ruta metabólica mediante la cual se da la producción de glucosa a partir de precursores no glucosídicos, tales como el lactato, piruvato… Puede considerarse la reacción inversa a la glucólisis, sin embargo, difiere en los puntos donde la reacción es irreversible. Síntesis de otras moléculas Síntesis de Lípidos Las grasas del organismo provienen principalmente por ingestión, pero pueden formarse también a partir de glúcidos y aminoácidos. Los componentes de las grasas o triacilglicéridos (glicerol y ácidos grasos), se sintetizan independientemente. El glicerol se genera por la reducción de un metabolito de la glucólisis. Los ácidos grasos se sintetizan a partir del acetil CoA, al que se incorporan a través de un conjunto cíclico de reacciones, originando moléculas con un número par y progresivo de carbonos. Síntesis de Aminoácidos Los organismos obtienen sus aminoácidos por medio de la dieta y también por procesos de síntesis. Sólo los seres autótrofos pueden sintetizar todos sus aminoácidos; otros no son capaces de sintetizar algunos de sus aminoácidos y los obtienen exclusivamente por la dieta. Estos aminoácidos se denominan aminoácidos esenciales. El organismo humano, por ejemplo, sólo es capaz de sintetizar 11 de los 20 aminoácidos necesarios. Por contraposición, se denominan aminoácidos no esenciales al resto de los aminoácidos. Por reacciones de transaminación se produce la síntesis de los aminoácidos no esenciales. La síntesis de las proteínas, polimerización de aminoácidos, es un proceso complejo que requiere la intervención de los ácidos nucleicos. Síntesis de Nucleótidos En la síntesis de nucleótidos, las rutas metabólicas de formación de las bases púricas y pirimidínicas son las más complejas. Todos los organismos a excepción de algunas bacterias son capaces de fabricar estas bases a partir de moléculas más sencillas. Las vías de síntesis difieren según sean las bases púricas o pirimidínicas. FOTOSÍNTESIS La fotosíntesis es un proceso anabólico que consiste en la transformación de energía solar en energía de enlace químico; es decir, la transformación de una forma de energía en otra más asequible para los seres vivos. ENERGÍA + CO2 + H2O GLÚCIDOS + H2O + O2 Es la reacción inversa a la degradación oxidativa de los glúcidos. El proceso abarca una serie de reacciones múltiples y encadenadas. ESTRUCTURAS QUE INTERVIENEN CLOROPLASTOS Son los orgánulos propios de los vegetales. Lugar donde se produce la fotosíntesis, concretamente en la membrana tilacoidal, el espacio intratilacoidal y el estroma. PIGMENTOS FOTOSINTÉTICOS Un pigmento es cualquier sustancia que absorbe luz. La clorofila es el pigmento fotosintético principal. Existen varias clases de clorofila, (clorofila A y clorofila B) que se distinguen por su estructura molecular. La mayoría de las células fotosintéticas, contienen, además, carotenoides, que son pigmentos rojos, anaranjados o amarillos que absorben la luz que la clorofila no puede absorber. FOTOSISTEMAS El fotosistema es una estructura constituida por el complejo de antena, encargado de asociar las moléculas de clorofila y carotenos, un dador y aceptor de electrones y un centro de reacción. Los fotosistemas captan energía luminosa que activará un electrón de la clorofila hacia un nivel energético superior. Existen dos tipos de fotosistemas, que se diferencian por su capacidad para captar energía: - - FOTOSISTEMA I: Posee un centro de reacción que absorbe la energía de la luz cuya longitud de onda es como máximo de 700 nm. En este sistema el aceptor de electrones es X y el dador es la plastocianina. FOTOSISTEMA II: Absorbe la energía de la luz cuya longitud de onda es como máximo de 680 nm. Los electrones son aceptados por la feofitina, y el agua actúa como dador. FASES DE LA FOTOSÍNTESIS La fotosíntesis presenta dos fases claramente diferenciadas: reacciones que captan energía y reacciones de fijación del carbono. 1) REACCIONES QUE CAPTAN ENERGÍA Son el conjunto de reacciones que transforman la energía solar en energía química. El resultado de éstas es la síntesis de ATP y obtención de poder reductor. Tienen lugar en la membrana tilacoidal de los cloroplastos. Se distinguen dos procesos: Fotofosforilación NO cíclica y Fotofosforilación cíclica. A) FOTOFOSFORILACIÓN NO CÍCLICA Es un proceso que supone el transporte de electrones, a través de moléculas especializadas, desde el H2O hasta el NADP+, proceso en el que se libera energía que es utilizada en la fosforilación del ADP. Es un proceso no espontáneo, por lo que es necesario el aporte de energía solar que es captada en los cloroplastos. El proceso consta de los siguientes pasos: Activación energética, por la luz, de un par de electrones del centro de reacción de los fotosistemas. Este par de electrones activados pasa a un nivel energético superior y es captado por una molécula con afinidad por los electrones. Transporte del par de electrones a través de diferentes moléculas dadoras y aceptoras de electrones (cadena de transporte electrónico). Reducción de la coenzima NADP+ al aceptar un par de electrones provenientes de la cadena de transporte, y un par de protones del medio. Fotólisis de una molécula de agua originándose un par de electrones, un par de protones y átomos de oxígeno. Utilización de la energía liberada en la cadena de transporte de electrones para la síntesis de ATP, proceso conocido como fotofosforilación, similar a la fosforilación oxidativa que tiene lugar en las mitocondrias. B) FOTOFOSFORILACIÓN CÍCLICA En la fotosíntesis se produce también un flujo cíclico en el que está implicado únicamente el fotosistema I y por el que los electrones de la clorofila son activados y vuelven otra vez a la molécula de clorofila, utilizando la energía desprendida para la síntesis de ATP. Al incidir la luz sobre el fotosistema I se activan un par de electrones que son captados por un transportador electrónico, cediéndolos a sucesivos componentes de la cadena transportadora de electrones, y utilizándose la energía liberada para la síntesis de ATP. Los electrones vuelven al fotosistema I. En este circuito, al no traspasarse los electrones al NADP+, se sintetiza únicamente ATP. 2) REACCIONES DE FIJACIÓN DEL CARBONO Son reacciones en las que el ATP y el NADPH + H + se utilizan como fuente de energía y de poder reductor para transformar sustancias inorgánicas oxidadas en moléculas orgánicas reducidas. Estas reacciones se producen en el estroma del cloroplasto y el proceso consta de una serie de reacciones conocidas como Ciclo de Calvin. CICLO DE CALVIN El ciclo de Calvin es un proceso que consiste en la síntesis de compuestos de carbono. Este conjunto de reacciones se producen en el estroma de los cloroplastos. El proceso comienza al reaccionar la ribulosa-1-5-difosfato con el CO2, originándose un compuesto de 6 carbonos que se escinde en dos moléculas de 3-fosfoglicerato. Sigue el ciclo de reacciones con la activación energética y reducción del 3-fosfoglicerato originando gliceraldehído-3-fosfato, molécula a partir de la cual se inician otras vías como síntesis de azúcares, grasas… Además, parte de las moléculas de gliceraldehído-3-fosfato se dedican a regenerar la ribulosa-1-5-difosfato para garantizar la continuidad del proceso. Balance En cada vuelta del ciclo de Calvin se consumen 3 moléculas de ATP y 2 NADPH2, siendo necesarias 3 vueltas del ciclo para formar una molécula de glicerldehído-3-fosfato. FACTORES QUE CONTROLAN LA FOTOSÍNTESIS La velocidad a la que discurre la fotosíntesis en cada organismo fotosintético está controlado por una serie de factores ambientales, como el CO2, H2O y la luz, por participar en la reacción. Sin embargo, la temperatura también influye en el proceso al condicionar el óptimo grado de actuación de las enzimas. EL CICLO CELULAR Las células realizan diversas funciones a lo largo de su vida. La división o reproducción celular es una parte del llamado Ciclo Celular. Este ciclo consta de varias partes: G1, S, G2, que componen la llamada interfase, y M (mitosis) y C (citocinesis) que forman parte de la división propia. MITOSIS La mitosis es un mecanismo de división celular cuya función principal es repartir los cromosomas maternos, ya duplicados previamente, en dos lotes iguales, de manera que las dos células hijas reciban la misma información genética que la madre. Se le llama también “división replicativa”. Se trata de un fenómeno de gran complejidad, con una serie de acontecimientos que han sido agrupados convencionalmente en 4 fases: profase, metafase, anafase y telofase. Una vez acabada la mitosis, se produce la llamada citocinesis o división del citoplasma. PROFASE Es la fase más larga. En ella, continúa condensándose la cromatina, de manera que poco a poco, se pueden distinguir los cromosomas. Aparece cada uno de ellos duplicado, formando por dos mitades, las llamadas cromátidas, que permanecen unidas entre sí por el centrómero. Estos cromosomas de la profase son cromosomas dobles, estructuras que forma la célula para poder repartir equitativamente su material genético. Mientras tanto, los pares de centriolos ya divididos se separan y van a colocarse en polos opuestos. Al mismo tiempo, van apareciendo diferentes microtúbulos que dan lugar al huso. Para este momento, la membrana nuclear y los nucléolos habrán desaparecido. METAFASE Es bastante corta. En ella, los cromosomas se mueven hasta colocar sus centrómeros en el ecuador del huso, equidistantes de los centriolos. En este momento, su condensación ha llegado al punto máximo. ANAFASE Es la fase más breve. Los centrómeros se parten en dos, y cada cromátida emigra hacia un polo diferente, transformándose en un cromosoma independiente. Al final, en cada polo se encontrarán dos conjuntos de cromosomas hermanos. TELOFASE Los cromosomas se descondensan y los microtúbulos van desapareciendo. La membrana nuclear se reconstruye alrededor de cada conjunto de cromosomas. Los nucléolos reaparecen. Al final, tenemos dos verdaderos núcleos interfásicos, cada uno con los mismos cromosomas que tenía la madre. A continuación se producirá la separación de estos, para obtener dos nuevas células. CITOCINESIS Una vez repartido el material genético nuclear se va a proceder a hacer lo mismo con el citoplasma. Los diferentes orgánulos se van repartiendo por invaginación de la membrana a la altura del ecuador del huso. En las vegetales se da la formación de una especie de tabique separador entre ambas mitades. Este tabique o placa celular se forma por una gran cantidad de vesículas procedentes del Aparato de Golgi. MEIOSIS La meiosis es una de las formas de reproducción celular. Este proceso se realiza en las glándulas sexuales para la producción de gametos. Es un proceso en el cual, una célula diploide experimenta dos divisiones, con la capacidad de dar 4 células haploides. En las dos divisiones de la meiosis, llamadas I y II, se observan una serie de acontecimientos bastante semejantes a los descritos en la mitosis. PROFASE I Es la más larga y complicada de toda la meiosis. Es necesario recordar que antes de la profase ya se habían duplicado los cromosomas, estando formadas las dos cromátidas. Los dos cromosomas homólogos que forman cada pareja se sitúan paralelamente. Este proceso se denomina sinapsis. Dado que hay cuatro cromátidas implicadas, el complejo se conoce como tétrada. Habrá tantas tétradas como pares de cromosomas tenga la célula. Una vez emparejados, se establecen entre las cromátidas unos puntos de unión, llamados quiasmas, en los cuales se intercambian trozos de cromosoma. En estos quiasmas se rompen las cromátidas y vuelven a unirse, pero cruzadas. En cada quiasma hay un intercambio entre dos de las cuatro cromátidas. Este proceso se conoce como entrecruzamiento o “crossing over”, en consecuencia, se produce un intercambio de genes. Las tétradas se van separando. Ya al final de la profase los cromosomas se han condensado mucho y tanto las cromátidas como los quiasmas son más fáciles de observar. El resto de acontecimientos son bastante parecidos a los de una profase mitótica normal: los centriolos han emigrado hacia los polos, la membrana nuclear ha desaparecido… METAFASE I Las tétradas se colocan en el ecuador, con sus pares de cromátidas enfrentadas. El plano ecuatorial no pasa por el centrómero de cada cromosoma, sino por los quiasmas y separa los cromosomas homólogos. ANAFASE I Los centrómeros no se dividen. Se separan los dos cromosomas homólogos, desaparecen los quiasmas y emigra cada uno a un polo opuesto. En consecuencia, se reduce el número de cromosomas a la mitad, si bien es cierto que son dobles. TELOFASE I Los pares de cromosomas homólogos completan su migración a los dos polos como resultado de la acción del huso. La membrana nuclear se vuelve a formar alrededor de cada juego de cromosomas, el huso desaparece y la citoquinesis continúa. Tras la citocinesis, cada una de las células hija tiene un núcleo con cromosomas recombinados haploides. MEIOSIS II Proceso similar a la mitosis. Las cromátidas de cada cromosoma ya no son idénticas, en razón de la recombinación. Se separan las cromátidas produciendo dos células hija, cada una con 23 cromosomas (haploides), y cada cromosoma con una sola cromátida. Cada una de estas células hija realizará el proceso que consta de las cuatro fases (profase, metafase, anafase y telofase) y finalmente, obtendremos 4 células haploides con distinto contenido genético, siendo diferentes a la célula inicial. LA GAMETOGÉNESIS En la especie humana, la meiosis se localiza en los órganos sexuales, ovarios y testículos. Aquí, a partir de células diploides, se van a producir, tras la meiosis, las únicas células haploides de nuestro cuerpo, los gametos. Aunque este proceso es similar en el hombre y la mujer, existen ciertas diferencias que deben ser destacadas. En el hombre, el proceso llamado espermatogénesis, comienza en la pubertad y en él, los espermatozoides haploides se forman a partir de unas células madre diploides, las espermatogonias. Estas células se dividen por mitosis, dando lugar a los espermatocitos primarios. En cada una de estas células ocurre la meiosis I, originándose dos espermátidas haploides. Finalmente, hay una diferenciación de estos últimos formándose los conocidos espermatozoides. En la mujer, se da la oogénesis, proceso similar al de los hombres, con ciertas diferencias importantes. Este proceso comienza durante el período embrionario, fase en la que las oogonias se transforman en oocitos primarios. Ahora comienza la meiosis I, pero ésta se detiene en la profase y así permanece hasta que se alcanza la madurez sexual. En este momento se reanuda la meiosis I formándose un oocito secundario grande y un pequeño primer corpúsculo polar. A continuación se produce la meiosis II, la cual se detiene al llegar a la metafase II. En este estado se produce la ovulación en la que el oocito secundario abandona el ovario. Sólo si tiene lugar la fecundación se completa la meiosis II, originándose un óvulo y un segundo corpúsculo polar. MEIOSIS Y GENÉTICA Basándose en diversas evidencias, Sutton y Boveri enunciaron la teoría cromosómica de la herencia según la cual los genes se localizan en los cromosomas. LEYES DE MENDEL 1º LEY Conocida como ley de la uniformidad de la generación F1. Esta ley establece que si se cruzan dos razas puras (homocigotos) para un determinado carácter, los descendientes (híbridos) de la primera generación serán todos iguales entre sí, en genotipo y fenotipo, e iguales a uno de los progenitores, en fenotipo. 2º LEY Conocida como ley de segregación de los alelos. Esta ley establece que para que ocurra la reproducción sexual, previo a la formación de gametos, cada alelo de un par se separa del otro miembro para determinar la constitución genética del gameto hijo. Retrocruzamiento En el caso de los genes que manifiestan herencia dominante, no existe ninguna diferencia aparente entre los individuos heterocigóticos (Aa) y los homocigóticos (AA), pues ambos individuos presentarían un fenotipo amarillo. La prueba del retrocruzamiento, o simplemente cruzamiento prueba, sirve para diferenciar el individuo homo- del heterocigótico. Consiste en cruzar el fenotipo dominante con la variedad homocigótica recesiva (aa). - Si es homocigótico, toda la descendencia será igual, en este caso se cumple la primera Ley de Mendel. - Si es heterocigótico, en la descendencia volverá a aparecer el carácter recesivo en una proporción del 50%. Codominancia Se denomina codominancia al fenómeno producido cuando un individuo expresa en su fenotipo tanto el carácter dominante como recesivo para determinada característica. Entendiendo como dominante aquel alelo que al ser comparado con otro, tiene más probabilidad de expresarse fenotípicamente. En otras ocasiones, cuando mezclamos dos individuos homocigóticos distintos las características de ambos progenitores aparecen en los individuos descendientes. Un ejemplo lo podemos observar en distintas variedades de plantas, como por ejemplo las rosas (Rosa sp.). Al cruzar un homocigoto de flores rojas (RR) con un homocigoto de flores blancas (BB) se producen heterocigotos que presentan partes rojas y partes blancas. La herencia de los grupos sanguíneos en la especie humana es también un ejemplo de codominancia. 3º LEY Conocida como ley de la independencia de los genes o ley de asociación independiente, establece que los genes que determinan cada carácter se transmiten independientemente. Ligamiento de Genes El ligamiento genético es la asociación de loci (posiciones de un gen en un cromosoma) genéticos que tienden a heredarse juntos. Si entre 2 loci que están en el mismo cromosoma no existen puntos de recombinación estos loci tienden a heredarse juntos. Los loci que están ligados genéticamente se heredan juntos en una mayor proporción de la esperada según el principio de transmisión independiente. Que dos loci estén ligados se mide por su frecuencia de recombinación. Así, se dice que están ligados cuando ésta es menor del 50%. Si ésta frecuencia es de 0, el ligamiento es completo. Teniendo en cuenta la frecuencia de recombinación entre genes se puede aproximar la distancia que separa a esos genes en los cromosomas. En los casos en que disponemos de genomas completamente secuenciados se ha comprobado que es una buena aproximación pero hay que tener en cuenta la existencia de los “hotspots” de recombinación, que presentan una frecuencia elevada de recombinación. Estos puntos alteran esas predicciones de localización ya que separan genéticamente a genes muy cercanos físicamente en el cromosoma. Recombinación Fenómeno que tiene lugar cuando, en la descendencia, aparece una combinación genética que no se encontraba en los progenitores. La recombinación implica que una hebra de ADN o de ARN se divide y se vincula a una molécula de un material genético que resulta diferente. Esto hace que los descendientes dispongan de una combinación genética que resulta distinta a la de sus padres. Dicha particularidad, de acuerdo a la biología evolutiva, aporta ciertas ventajas a los organismos, permitiendo que eviten mutaciones que resultan dañinas. Existen diferentes clases de recombinación. La recombinación general u homóloga ocurre en la meiosis I, más precisamente su profase, y tiene lugar en aquellas secuencias de ADN que son muy parecidas entre sí pero que no resultan iguales. La recombinación no homóloga, en cambio, acontece entre secuencias que no cuentan con una relación de homología. La recombinación específica de sitio, por su parte, se produce por una rotura de sectores de homología específica de un par de secuencias de ADN diferentes o que forman parte de una misma molécula. La noción de recombinación también aparece en otras cuestiones vinculadas a la biología. La frecuencia de recombinación, en este sentido, es un indicador que revela la distancia existente entre dos loci (es decir, entre dos posiciones de un gen en un cromosoma). Un nódulo de recombinación, por último, es el sitio donde tiene lugar el entrecruzamiento de cromosomas de tipo homólogo. Estos nódulos están incluidos en los complejos sinaptonémicos. Entrecruzamiento El entrecruzamiento es un proceso mediante el cual se intercambian partes de cromosomas homólogos; es decir, es el proceso por el cual dos cromosomas se aparean e intercambian secciones de ADN. Los entrecruzamientos no ocurren forzosamente en el 100% de las células que están en la meiosis. Es un fenómeno que depende del azar. Su probabilidad es mayor cuanto más alejados estén los loci (posiciones de un gen en un cromosoma) de ambos genes. Es la distancia la que decide. A mayor distancia, más % de entrecruzamientos, y en consecuencia, mayor frecuencia de recombinantes. En conclusión, la frecuencia de recombinaciones entre dos genes puede usarse como medida de la distancia física entre ambos. HERENCIA DEL SEXO La composición cromosómica no es totalmente igual en ambos sexos. Siempre hay una pareja de cromosomas especial, diferente en el macho que en la hembra; son los cromosomas sexuales o heterocromosomas. Todos los demás, llamados autosomas, presentan el mismo aspecto en ambos sexos. En las células femeninas de todas las especies animales, incluida la nuestra, los dos cromosomas sexuales son iguales y se denominan X. Sin embargo en los machos, uno de los cromosomas es en apariencia igual a un X, mientras que el otro es diferente, el llamado Y. Es decir, en hembras encontramos XX y en machos XY. ¿Cómo se hereda el sexo? Resulta que la hembra se comporta como el homocigoto recesivo para el sexo (genotipo XX), dando en la meiosis un solo tipo de gametos para este carácter (todos con X). El macho será heterocigoto recesivo (XY), dando gametos diferentes, unos con X y otros con Y. En este tipo de especies se dice que las hembras son homogaméticas y los machos heterogaméticos. MUTACIONES Una mutación es un cambio en el material genético. Puede ser que cambie la composición del DNA, su cantidad o simplemente la disposición u organización del mismo en el genotipo del individuo. Se han utilizado diferentes criterios a la hora de clasificar las mutaciones. Si nos fijamos en el tipo de células afectadas, tenemos las heredables, que se dan en los gametos, y las no heredables, que se dan en otras células, en cuyo caso sólo afectarían al individuo donde se producen. Otro criterio es la cantidad de material genético afectado por el cambio. Según este aspecto, se suele hablar de mutaciones génicas y mutaciones cromosómicas. Las génicas afectan a un solo gen, mientras que las cromosómicas a una porción mayor de DNA. MUTACIONES CROMOSÓMICAS Son aquellas mutaciones que afectan a una porción significativa de DNA. Cuando estas mutaciones suponen cambios en el número de cromosomas se denominan genómicas, y si afectan a la estructura de los mismos, estructurales. - - Mutaciones genómicas: Son mutaciones en las que puede haber variación en uno o más cromosomas enteros (aneuploides) o en juegos completos de cromosomas (euploides). Mutaciones estructurales: Son mutaciones que afectan a un fragmento de cromosoma. Muchas malformaciones humanas derivan de este tipo de problemas. Hay varios tipos de mutaciones estructurales: A) MUTACIÓN POR DELECCIÓN: Se pierde una parte de cromosoma (pérdida de genes). B) DUPLICACIÓN: Ganancia de genes. C) TRASLOCACIÓN: Variación en la posición de los genes. D) INVERSIÓN: Cambio de posición en los genes. E) FUSIÓN: Unión de dos cromosomas o fragmentos de cromosomas. MUTACIONES GÉNICAS Las mutaciones génicas son las que afectan a un solo gen. Pueden ser puntuales a nivel del DNA. Consisten bien en una sustitución (cambio de una base), bien en una inserción (adición de un nucleótido) o bien en una delección (pérdida de un nucleótido). Las modificaciones sufridas por el DNA pueden tener o no tener repercusiones sobre la síntesis de polipéptidos: - Una mutación que no entraña modificación alguna se dice que es una mutación silenciosa. Una mutación neutral se da cuando la modificación del aminoácido no altera la función de la proteína. Una mutación en falso sentido se da si la función de la proteína se altera o modifica, y por tanto, la mutación se expresa. CAUSA Y FRECUENCIA DE LAS MUTACIONES La mayoría de las mutaciones se dan de forma espontánea y al azar en nuestra naturaleza, sin que se conozcan las causas concretas que lo provocan. En los complicados procesos de duplicación del DNA, en la mitosis y en la meiosis, pueden aparecer errores que den lugar a una mutación espontánea. Aparte de estos cambios espontáneos, se conocen muchos agentes químicos, físicos e incluso biológicos, capaces de producir alteraciones en el material genético. Estos agentes pueden incidir en las células de manera natural (temperatura, rayos cósmicos, ultravioletas…). Pero existen muchos que proceden de las actividades humanas (sustancias del tabaco, aditivos, radiaciones de energía altas…), capaces de incrementar la frecuencia de las mutaciones. CONSECUENCIAS Los efectos de las mutaciones pueden ser muy variables. La mayoría son perjudiciales, ya que lo normal es que las nuevas formas estén peor adaptadas que las anteriores. Tal es el caso de las mutaciones génicas que conducen a la inactivación de proteínas cruciales para el organismo. También hay muchos casos en los que las mutaciones no se manifiestan, y finalmente, en algunos casos pueden tener efectos inmediatos beneficiosos. Pero la consecuencia principal de cara a la evolución es que contribuyen a aumentar la variación genética de las especies, al dar lugar a nuevos alelos y nuevas combinaciones de los mimos. REPLICACIÓN DE LOS GENES Una propiedad esencial del material genético es su capacidad para hacer copias exactas de sí mismo. El mecanismo de replicación del DNA sugerido por Watson y Crick se llama “replicación semiconservativa” ya que se conserva la mitad de la molécula. Cada cadena vieja forma el molde para la producción de una nueva. Proceso de replicación El proceso de replicación del DNA consiste en la separación de las dos cadenas del DNA y en la síntesis de otras dos nuevas con una secuencia de bases complementaria de las anteriores. El proceso real requiere una doble hélice de DNA, nucleósidos trifosfato, y un cojunto de enzimas diferentes que catalizan determinados pasos del proceso. El DNA debe desenrollar su doble hélice para que ambas cadenas puedan actuar como moldes. La replicación siempre comienza con una secuencia específica de nucleótidos, conocida como origen de replicación. Una vez separadas las dos cadena, proceso que requiere enzimas conocidas como helicasas, que catalizan la ruptura de los puentes de hidrógeno entre las bases complementarias, comienza la síntesis de las dos nuevas cadenas catalizada por la enzima conocida como DNA polimerasa. La DNA polimerasa cataliza la formación de un enlace fosfodiéster entre el nuevo desoxirribonucleótido trifosfato y la cadena que se está sintetizando, con la liberación de dos moléculas de ácido fosfórico. La energía necesaria para formar el enlace fosfodiéster proviene de la hidrólisis de los enlaces entre fosfatos. La zona de síntesis aparece como una “burbuja de replicación”, donde las cadenas viejas comienzan a ser separadas por la helicasa y las cadenas nuevas están siendo sintetizadas. La molécula forma una estructura en forma de Y conocida como horquilla de replicación. Puesto que la DNA polimerasa sólo cataliza la replicación en el sentido 5´ - 3´, y teniendo en cuenta que las cadenas son antiparalelas, sólo una de ellas puede sintetizarse de forma continua; la llamada cadena o hebra adelantada. La otra cadena, la conocida como cadena o hebra retardada, se sintetiza discontinuamente, a partir de fragmentos de DNA, denominados fragmentos de Okazaki, que se unen entre sí mediante una enzima llamada DNA ligasa. La replicación del DNA es, por tanto, asimétrica. Para que ocurra la síntesis de la cadena retardada, también debe estar el inicio de la nueva cadena, ya que la DNA polimerasa necesita una cadena previa a la que seguir uniendo nucleótidos. Este inicio lo provee un cebador, formado por una corta cadena de RNA cuya síntesis está catalizada por una enzima conocida como RNA primasa. Durante el proceso de maduración, además de unirse los fragmentos de Okazaki, se eliminan los fragmentos de RNA, que son sustituidos por DNA. EXPRESIÓN DE LOS GENES. EL CÓDIGO GENÉTICO Y LA SÍNTESIS DE PROTEÍNAS Tras diversos experimentos, Beadle y Tatum demostraron que ciertas mutaciones afectaban a enzimas que catalizaban diferentes pasos de la vía de reacción para la síntesis de aminoácidos. Basándose en sus estudios, Beadle y Tatum formularon el principio un gen – una enzima. Posteriormente se corrigió este principio, pues si bien las enzimas son proteínas, no todas las proteínas son enzimas, y un gen – una enzima fue sustituido por un gen – una proteína. Al descubrirse que muchas proteínas estaban formadas por más de una cadena polipeptídica, el principio se modificó de nuevo, quedando como un gen – una cadena polipeptídica. DEL DNA A LA PROTEÍNA Como resultado de los estudios sobre el DNA, los científicos propusieron que la secuencia de nucleótidos del DNA podría determinar la secuencia de aminoácidos en la cadena polipeptídica. La cuestión era: ¿de qué manera el orden de bases del DNA especifica la secuencia de aminoácidos de una proteína? Para entonces existían diferentes pistas que indicaban que el RNA podía desempeñar un papel en la traducción de la información genética del DNA a una secuencia de aminoácidos. El RNA se sintetiza muy rápidamente en páncreas y otros órganos productores de gran cantidad de proteínas. La evidencia adicional fue suministrada por los virus. Cuando una célula bacteriana es infectada por un bacteriófago que contiene DNA, se sintetiza RNA del DNA viral antes de que comience la síntesis de proteínas virales. Basado en estas evidencias, Crick proclamó lo que llamó dogma central de la genética molecular: El DNA especifica RNA, que a su vez especifica proteínas. El genotipo (DNA) determina el fenotipo dictando la composición de las proteínas. Sin embargo, las proteínas no alteran el genotipo, es decir, las proteínas no envían interacciones de regreso al DNA. TRANSCRIPCIÓN. SÍNTESIS DEL RNA Se denomina transcripción al proceso mediante el cual la información genética del DNA se transfiere a moléculas de RNA. Es decir, la transcripción consiste en la síntesis de moléculas de RNA con secuencias de bases complementarias a los correspondientes genes del DNA. La transcripción comienza cuando la enzima RNA polimerasa reconoce, y se une a una secuencia de nucleótidos denominada promotor. A partir de este punto, la enzima va desenrollando la doble hélice de DNA, una de cuyas cadenas servirá de molde para la síntesis de una cadena de RNA complementaria. La enzima RNA polimerasa va desplazándose a lo largo de la cadena de DNA molde y cataliza la unión de sucesivos ribonucleótidos al OH 3´ de la cadena de RNA en formación, con la liberación de dos moléculas de ácido fosfórico. La energía necesaria para crear el enlace fosfodiéster proviene de la hidrólisis de los enlaces entre fosfatos. El proceso de crecimiento de la cadena de RNA continúa hasta que la enzima encuentra una señal especial de terminación. En ese punto la RNA polimerasa se separa del DNA y libera la molécula de RNA transcrita. La cadena que debe transcribirse es determinada por la orientación del promotor, ya que sólo puede ser leído correctamente por la RNA polimerasa en un solo sentido. En las células procarióticas una misma RNA polimerasa transcribe los tres tipos de RNA. En las células eucarióticas hay tres RNA polimerasas diferentes: una transcribe los genes que se traducirán a proteínas; es decir, cataliza la síntesis de RNAm, una segunda transcribe los genes de los RNA ribosómicos grandes, y una tercera transcribe para una variedad de RNA pequeños, incluyendo RNAt y los RNA pequeños del ribosoma. MODIFICACIÓN Y MADURACIÓN DEL RNAm TRANSCRITO EN EUCARIOTAS En las células procariotas, cada molécula de RNAm es una secuencia continua de nucleótidos que codifica la secuencia de aminoácidos de una proteína; es decir, en las células procariotas el RNAm transcrito es traducido directamente. En las células eucariotas, el RNA nuclear contiene numerosas secuencias de nucleótidos, denominadas intrones, que no codifican para ningún aminoácido y que son el resultado de la transcripción de regiones de DNA. Las regiones que sí codifican los aminoácidos de la proteína se denominan exones. Antes de que la molécula de RNAm deje el núcleo, los intrones deben ser cortados del RNA transcrito con el fin de convertirlo en una molécula de RNAm que pueda codificar la síntesis de una proteína. Puesto que los exones se unen uno a otro tras haber sido seleccionados y eliminados los intrones, esta reacción de transformación del RNA se conoce como maduración. EL CÓDIGO GENÉTICO Las proteínas contienen 20 aminoácidos diferentes, pero el DNA y el RNA contienen, cada uno, solo cuatro nucleótidos diferentes. Si sólo cada uno de los cuatro nucleótidos “codifica” un aminoácido, entonces solamente cuatro aminoácidos podían ser especificados por las cuatro bases. Entonces podría haber un número máximo de 16 aminoácidos, lo cual era insuficiente para codificar los 20 aminoácidos. Por tanto, como mínimo eran necesarios tres nucleótidos en la secuencia para especificar cada aminoácido. En este caso, resultarían 64 combinaciones posibles, o codones, lo cual es más que suficiente para los 20 aminoácidos. El código genético consiste en 64 combinaciones de tripletes (codones) y sus aminoácidos correspondientes. De las 64 combinaciones de tripletes, 61 especifican aminoácidos particulares y 3 son codones de terminación. Hay más de un codón para la mayoría de los aminoácidos; por esta razón, se dice que el código genético es degenerado. Para todos los organismos el código genético es universal. Los experimentos realizados hasta la fecha indican que el código genético nuclear es universal, de manera que un determinado triplete o codón lleva información para el mismo aminoácido en diferentes especies. SÍNTESIS DE PROTEÍNAS Las instrucciones para la síntesis de proteínas están codificadas en las moléculas de DNA. La replicación semiconservadora del DNA transmite estas instrucciones de la célula madre a las células hija y, de esta manera, cada célula nueva hereda la información necesaria para sintetizar las proteínas específicas que determinan su estructura y sus funciones. Las instrucciones del DNA se transcriben; es decir, en una molécula de RNAm y son estas “copias” las que se usan en la síntesis de proteínas para determinar las secuencias de aminoácidos. La síntesis de proteínas se realiza en el citoplasma y requiere, además de las moléculas de RNAm, los otros dos tipos de RNA (RNAr y RNAt). Adaptación de los aminoácidos a sus correspondientes RNAt Para cada aminoácido existe un RNAt específico, pero ¿cómo reconoce cada molécula de RNAt a su aminoácido correspondiente? Existen un grupo especial de enzimas, denominadas aminoacil-RNAt sintetasas, que acoplan cada aminoácido a su molécula apropiada de RNAt. Es el RNAt, a través de su anticodón, el que reconoce el codón del RNAm específico del aminoácido. Para cada aminoácido existe una sintetasa diferente. Esta reacción consume ATP, y puede resumirse así: Aminoácido + RNAt + ATP Aminoacil-RNAt + AMP + PP Traducción La síntesis de proteínas se conoce como traducción, dado que es la transferencia de información de un lenguaje (el de nucleótidos) a otro (el de los aminoácidos). La traducción se realiza en los ribosomas. Estos poseen la estructura adecuada para poder albergar a los sucesivos RNAt y a la vez son capaces de desplazarse a lo largo de la molécula de RNAm codón a codón. Cada ribosoma contiene dos lugares de unión para los RNAt. En uno de ellos se sitúa la molécula de RNAt que contiene el último aminoácido unido a la cadena polipeptídica y se denomina lugar de unión del peptidil RNAt o centro P. Al otro se une la molécula de RNAt que aporta el aminoácido siguiente de la cadena, y se denomina lugar de unión del aminoacil RNAt o centro A. El proceso de síntesis ocurre en tres etapas: iniciación, alargamiento y terminación. Iniciación La subunidad ribosómica más pequeña se une al extremo 5´de una molécula de RNAm. La primera molécula de RNAt que lleva el aminoácido modificado se coloca en su lugar para aparearse con el codón iniciador AUG de la molécula de RNAm. La formilmetionina será el primer aminoácido de la cadena polipeptídica recién sintetizada, aunque luego puede ser removido. La combinación de la subunidad ribosómica pequeña, el RNAm y el aminoacil RNAt iniciador, se conoce como el complejo de iniciación. Luego la subunidad mayor del ribosoma se coloca en su lugar, el formilmetionil-RNAt ocupa el centro P, dejando vacante el centro A. La energía necesaria para esta etapa la suministra la hidrólisis del GTP y el proceso requiere además factores de iniciación. Alargamiento Durante esta etapa, la cadena polipeptídica va creciendo por adición de sucesivos aminoácidos. En esta etapa tienen lugar varios procesos: a) Reconocimiento del siguiente codón por el aminoacil RNAt correspondiente que se incorpora al centro A del ribosoma. Esto implica apareamiento de bases complementarias de codón (RNAm) y anticodón (RNAt). Se requieren factores de alargamiento y consumo de energía (GTP). b) Formación del enlace peptídico entre el aminoácido unido al RNAt del centro P y el recién incorporado en el centro A, de forma que la cadena polipeptídica quede unida al RNAt del centro A. Esta reacción es catalizada por la enzima peptidil transferasa que forma parte de la subunidad mayor del ribosoma. c) Traslocación. El ribosoma se desplaza tres nucleótidos hacia el extremo 3´ del RNAm. El RNAt libre ocupa el centro P. Este proceso requiere nuevamente factores de alargamiento y GTP. Esta etapa continúa hasta que el ribosoma llega a alguno de los codones de terminación. Terminación Cuando el ribosoma llega a alguno de los codones de terminación, para los que no existen RNAt con anticodones complementarios, la síntesis se detiene gracias a la participación de ciertos factores de liberación. Estos factores pueden incorporarse al centro A vacío del ribosoma, alterando la especificidad de la enzima peptidil transferasa, que ahora es capaz de hidrolizar el enlace de la cadena polipeptídica al RNAt. De esta forma queda libre la proteína recién sintetizada y el último RNAt. Finalmente, el ribosoma se separa del RNAm y las dos subunidades se disocian, Una vez concluida la síntesis de la cadena polipeptídica, ésta deberá sufrir los plegamientos necesarios para adquirir su conformación nativa (estructura secundaria y terciaria). REGULACIÓN DE LA EXPRESIÓN GÉNICA 1. REGULACIÓN GÉNICA EN PROCARIOTAS. TEORÍA DEL OPERÓN El mecanismo de biosíntesis de proteínas podría sugerir una síntesis continua e indiscriminada de todas las proteínas. Sin embargo, existen pruebas de que sólo se sintetizan en un momento dado aquellas proteínas necesarias en ese momento y en cantidades adecuadas. Esto precisa un mecanismo de regulación de la actividad del gen. Uno de estos mecanismos es el del Operón, propuesto por Jacob y Monod en 1961. En el modelo del operón se diferencian dos tipos de genes: los genes estructurales, que codifican las proteínas estructurales y enzimáticas, y los genes reguladores, que codifican las proteínas llamadas represores, cuya misión es controlar la transcripción de los genes estructurales. El gen regulador está más o menos alejado, en la cadena de DNA, de los genes estructurales. Junto a éstos hay dos zonas específicas: la zona promotor, donde se fija la RNApolimerasa, y la zona operador, donde se fija el represor. La transcripción del gen regulador y la posterior traducción del RNAm en los ribosomas originan el represor que se asocia a la zona del operador e impide que la RNA-polimerasa transcriba los genes estructurales. Existen moléculas denominadas inductores, que al asociarse a los represores los inactivan, provocando cambios en su estructura, permitiendo que la RNApolimerasa transcriba los genes estructurales. Estos operones funcionan según la denominada inducción enzimática. Otros operones funcionan según la denominada represión enzimática. El gen regulador codifica la formación de un represor inactivo, pero si aparece una molécula específica llamada correpresor el represor se vuelve activo y el complejo represor-correpresor se fija en la zona del promotor e impide la transcripción de los genes estructurales. 2. REGULACIÓN GÉNICA EN EUCARIOTAS En las células eucariotas, al existir el proceso de maduración del RNA transcrito en el núcleo, éste podría ser una de las etapas de regulación de la expresión de los genes en las distintas células. Así, aunque un gen se transcribiera en todas las células, sólo en algunas de ellas llegaría a madurar, siendo en éstas en las que finalmente se expresaría el gen. MANIPULACIÓN GENÉTICA. TRABAJANDO CON GENES El estudio de la replicación y expresión de los genes así como de los procesos de transferencia de información genética de una célula a otra, ha permitido desarrollar técnicas de manipulación genética, conocidas con el nombre de tecnología del DNA recombinante. Estas técnicas no sólo han permitido continuar investigando la estructura y función de los genes sino que ofrecen grandes posibilidades al campo de la biotecnología. PROCESOS NATURALES DE TRANSFERENCIA DE GENES DE UN CÉLULA A OTRA En los organismos eucarióticos, la reproducción sexual permite la mezcla de la información genética de dos individuos diferentes. Las células bacterianas pueden adquirir nueva información genética de tres maneras diferentes: la transformación, que es la captación de fragmentos de DNA del medio circundante; la conjugación, que es la transferencia directa de DNA de una célula a otra y la transducción, transferencia por virus de material genético de una célula a otra. El DNA adquirido por parte de la célula receptora puede recombinarse con regiones homólogas de la propia célula receptora o puede ser mantenido como un minicromosoma (un plásmido), replicándose de forma autónoma y transmitiéndose a las células hija en la división celular. TRANSFORMACIÓN BACTERIANA La transformación bacteriana es el proceso por el cual se modifica el genoma de una bacteria por la captación de moléculas de DNA presentes en el medio. Cuando una población de bacterias con una incapacidad genética (por ejemplo, incapacidad para formar la cápsula), se expone a un DNA purificado de bacterias capaces de realizar esa función, algunas de las células adquieren la capacidad del DNA donante. Estas transformaciones son estables y son heredables debido a que el gen funcional adquirido pasa a formar parte del DNA genómico en las células receptoras. CONJUGACIÓN La conjugación es la transferencia directa de DNA desde una bacteria a otra como consecuencia de un contacto entre células. El proceso ocurre de la siguiente manera: Dos células, una capaz de donar un cromosoma a la otra, establecen contacto a través de un puente de conjugación de naturaleza proteica. La replicación del DNA comienza por una posición especial del cromosoma de la célula donante y el DNA recién replicado es transferido a la célula receptora. La transferencia se interrumpe cuando el puente se rompa, por lo tanto, cuanto más tiempo hayan estado en contacto las dos células antes de que se rompa el puente, más genes habrán sido transferidos. El DNA transferido puede sustituir, por recombinación, a la región correspondiente del genoma de la célula receptora. TRANSDUCCIÓN La transducción es la transferencia de DNA celular de una célula huésped a otra por medio de virus. Cuando ciertos bacteriófagos, conocidos como bacteriófagos atenuados, infectan a bacterias pueden ocurrir dos cosas: que el fago explote los recursos de la célula para replicarse, producir nuevos fagos y terminar lisando a la bacteria (ciclo lítico); o que el DNA viral se incorpore al cromosoma bacteriano como profago, replicándose con él y siendo transferido a las células hijas (ciclo lisogénico). Además de su propio DNA, los fagos son capaces de adquirir DNA de la bacteria que infectan y transferirlo a otras bacterias durante el transcurso de una infección posterior. Según los fagos adquieran DNA en un ciclo lítico o en un ciclo lisogénico, se distinguen dos tipos de transducciones: a) Transducción general. Durante el ciclo lítico, el DNA de la bacteria se fragmenta y estos fragmentos de DNA bacteriano son empaquetados accidentalmente en la estructura del fago. Estos fagos transductores introducen DNA bacteriano recién adquirido en otras bacterias al infectarlas y los genes transferidos pueden incorporarse al cromosoma de la célula bacteriana receptora. b) Transducción restringida. Cuando los profagos se separan del cromosoma bacteriano para iniciar un ciclo lítico; puede que la escisión del DNA vírico sea precisa o puede ocurrir que lleven con ellos un fragmento del cromosoma de la célula huésped en lugar de algunos genes fágicos. Durante la próxima infección, la bacteria adquiere, además de los genes fágicos, información genética del huésped anterior. TÉCNICAS UTILIZADAS EN LA METODOLOGÍA DEL DNA RECOMBINANTE PARA INVESTIGAR LA ESTRUCTURA DEL DNA Las técnicas más importantes utilizadas en la tecnología del DNA recombinante son: métodos para obtener segmentos específicos de DNA de un tamaño adecuado para su análisis y secuenciación, clonado de DNA que permite la producción de gran cantidad de dichos segmentos, y secuenciación del DNA que permite la determinación del orden de nucleótidos en un segmento de DNA. MÉTODOS PARA OBTENER SEGMENTOS ESPECÍFICOS DE DNA Son dos los métodos utilizados para obtener segmentos de DNA de un tamaño adecuado que permita su posterior análisis: la utilización de enzimas de restricción y la utilización de la transcriptasa inversa. - Utilización de enzimas de restricción. Las enzimas de restricción son enzimas que se unen a un DNA de cadena doble y lo cortan en sitios específicos. Su función es hidrolizar el DNA extraño, es decir, el DNA procedente de otra cepa bacteriana o de fagos. Cada enzima reconoce una secuencia específica de cuatro a seis nucleótidos. No todas las enzimas de restricción hacen cortes rectos en ambas cadenas. Algunas cortan las cadenas con algunos nucleótidos de diferencia, dejando extremos adhesivos o “pegajosos”. Estos extremos pueden unirse con otro segmento de DNA de cualquier origen que haya sido cortado por la misma enzima de restricción y que tenga extremos adhesivos complementarios. Permiten, por lo tanto, unir los segmentos de DNA al DNA de un vector (un plásmido o un fago) para que puedan ser clonados. Los segmentos de DNA producidos por enzimas de restricción que actúan sobre el DNA de una célula se conocen como DNA genómico o gDNA. - Utilización de la transcriptasa inversa. Otro método para obtener segmentos específicos de DNA consiste en utilizar una enzima de los virus denominados retrovirus. En estos virus, el RNA sirve como molde para sintetizar DNA gracias a una enzima viral conocida como transcriptasa inversa. La molécula de DNA sintetizada por la transcriptasa inversa se conoce como DNA complementario o cDNA. Tanto el RNA nuevo como el RNAm para la síntesis de proteínas virales se transcribe a partir de este cDNA. Este método presenta la ventaja, frente a la utilización de los enzimas de restricción, de que el cDNA representa genes y no fragmentos de DNA. CLONADO DE DNA Una vez cortado el genoma mediante las enzimas de restricción e introducidos los fragmentos resultantes en un vector (plásmido), el paso siguiente es clonar las moléculas de DNA recombinante. Para ello la población de moléculas de DNA recombinante se mezcla con células en las cuales el vector es capaz de replicarse y, en condiciones adecuadas, las células adquieren un solo vector. A continuación, la suspensión de células se extiende sobre una placa de Petri de tal forma que las células queden separadas unas de otras. Para identificar los clones que contienen DNAs recombinantes, se seleccionan plásmidos que contengan genes que confieran resistencia a algún antibiótico y se insertan los fragmentos de DNA extraño en uno de esos genes. De esta manera, las colonias que contengan vectores recombinantes pueden distinguirse de las que poseen plásmidos sin insertos, gracias a que las que contienen DNA recombinante dejan de ser resistentes a ese antibiótico. Mediante estas técnicas de clonado se obtienen un conjunto de clones que contienen, cada uno, un DNA vector con un inserto. Al conjunto de clones con estas características se le denomina biblioteca recombinante. De la misma manera que pueden clonarse DNAs genómicos pueden clonarse DNAs complementarios y obtenerse bibliotecas de cDNA. SECUENCIACIÓN DEL DNA Una vez clonados los distintos fragmentos de DNA, pueden ser estudiados para conocer su secuencia de nucleótidos. El método más utilizado, consiste en marcar las múltiples copias del fragmento de DNA de cadena sencilla en el extremo 5´con un fosfato radiactivo. La solución que contiene el DNA marcado se divide en cuatro porciones, cada una de ellas se somete a un tratamiento químico diferente que rompe la molécula por una de las cuatro bases que es eliminada. Los fragmentos resultantes son separados sobre placas calibradas para indicar la posición ocupada por los fragmentos de diferentes longitudes. Combinando la información obtenida en cada porción, puede conocerse la secuencia completa del fragmento de DNA. Teniendo en cuenta que los fragmentos obtenidos por las distintas enzimas de restricción se superponen, las secuencias de nucleótidos pueden reunirse como un rompecabezas para mostrar la secuencia completa de una molécula de DNA o de un gen aislado. TÉCNICAS UTILIZADAS EN LA METODOLOGÍA DEL DNA RECOMBINANTE PARA TRANSFERIR UN GEN DE UNA ESPECIE A OTRA El conjunto de técnicas utilizadas para introducir un gen extraño en un organismo se conoce como ingeniería genética. La finalidad de esta transferencia puede ser la producción de grandes cantidades de una sustancia, la obtención de variedades de plantas o de animales que presenten la característica ligada al gen transferido o en un futuro próximo, la terapia génica en el hombre. Para la transferencia de genes se requieren distintas técnicas: 1. Aislamiento del gen, bien utilizando enzimas de restricción y separando el gen deseado por hibridación, o bien obteniendo el cDNA a partir del RNAm utilizando la transcriptasa inversa. 2. Inserción de un gen en un vector adecuado. Estos vectores deben ser fácilmente manipulables y deben contener alguna característica que permita distinguir, en la nueva generación, las células que han incorporado el vector con inserto de las que no lo han hecho. 3. Clonación y aislamiento de los individuos recombinantes. 4. Expresión del gen. La expresión de un gen clonado, es decir, la transcripción y traducción del gen, requiere que el gen vaya acompañado de las señales necesarias para su transcripción. Es necesario, por lo tanto, añadir estas señales al gen e introducir el gen así completado en un nuevo vector conocido como vector de expresión. Estos vectores de expresión son transferidos a células huéspedes capaces no sólo de producir el polipéptido codificado, sino de transmitir esta propiedad a sus descendientes. GENES Y CÁNCER El cáncer es una enfermedad en la cual las células escapan a los factores que regulan el crecimiento celular normal. En condiciones normales, las células se multiplican únicamente cuando es necesario, durante el desarrollo y, más tarde, para reponer las células perdidas debidas al proceso normal de renovación, o a procesos traumáticos. Cuando el autocontrol se rompe, las células se multiplican, amontonándose, y originan un tumor maligno. Los tumores pueden ser sólidos (cánceres de mama, útero…) o pueden ser cánceres sistémicos (leucemias, linfomas…). La mayoría de los tumores son clones de células, descendientes todas de una célula que se transformó en célula tumoral. Los tumores malignos, además de invadir los tejidos adyacentes, pueden extenderse a otras partes del organismo, ya que las células pueden separarse del tumor inicial y viajar a través del sistema linfático y circulatorio a otros tejidos, originando tumores secundarios. A este proceso se le conoce con el nombre de metástasis. El desarrollo del cáncer se ha relacionado con cambios en el material genético. La mayoría de los tumores son clones de células cancerosas, lo que indica que el cáncer es una propiedad heredada de las células; además anormalidades cromosómicas son visibles en las células cancerosas y la mayoría de los carcinógenos o cancerígenos (agentes que producen cáncer) son también mutágenos. Al estudiar las células transformadas se han descubierto un grupo de genes conocidos como oncogenes. Los oncogenes se asemejan mucho a los genes normales de las células eucariotas en los que se encuentran. Según la “teoría de la mutación”, el cáncer es causado cuando algo funciona mal en la expresión de genes normales como resultado de mutaciones en los genes. Estos genes parece ser que codifican proteínas reguladoras implicadas en el control del crecimiento celular o en la división celular. Sólo unos pocos cánceres humanos se han asociado a virus. Estos virus, que lo mismo que los carcinógenos o cancerígenos provocan cambios en la constitución genética de las células, pueden ser virus DNA o retrovirus del RNA. Según la “teoría viral”, los virus pueden causar cáncer de distintas maneras: 1. Los virus poseen genes que codifican proteínas cuya acumulación altera los mecanismos de control del crecimiento celular. 2. Los virus actúan como vectores de oncogenes. Los oncogenes víricos codifican proteínas capaces de transformar a las células normales en células cancerosas. Se han propuesto mecanismos comunes para el cáncer producido por mutación y el cáncer inducido víricamente. Uno de estos mecanismos postula que en los dos casos, la transformación neoplástica (crecimiento celular incontrolado) es atribuible a la hiperproducción de una proteína reguladora celular. MICROBIOLOGÍA ¿Qué son los microbios? Historia de la microbiología. Los microorganismos se definen como seres vivos microscópicos. Los procesos que realizan estos organismos se conocen desde hace muchos años y se han intentado controlar. Por ejemplo, en papiros egipcios antiguos aparecen instrucciones para preparar pan fermentado, vino y cerveza. Es muy probable desde aquellas épocas se conozca la conservación de alimentos mediante secado, salado y deshidratación, para aprovechar los excedentes de producción por medio del control de la actividad microbiana que produce la descomposición de los mismos. En el siglo XIX se produjo un gran impulso de la microbiología, que se basó en el desarrollo de las técnicas de la microscopía óptica y de manipulación de los microorganismos. Así, a principios del siglo XIX, se empiezan a fabricar los primeros microscopios compuestos con lentes acromáticas, que permiten ver con claridad objetos del tamaño de las bacterias y mejorar las condiciones de las observaciones microscópicas. A finales del siglo XIX ya eran conocidas la mayoría de las características morfológicas externas de las bacterias, a excepción de algunas muy pequeñas invisibles al microscopio óptico. Tipos de microorganismos Hacia el año 1800, los microbios se consideraban animales por su movilidad y se denominaban infusorios. Posteriormente, un mayor conocimiento de los organismos microscópicos llevó a considerar que entre ellos existen formas distintas, aunque todavía hacia 1860 se les incluía entre los animales o vegetales, teniendo en cuenta su movilidad y capacidad autótrofa. En 1866 Haeckel propuso que vegetales y animales constituyen reinos distintos del constituido por organismos de estructura sencilla, tales como las algas, hongos, protozoos y bacterias. A estos últimos los denominó protistas, entre los que incluía los unicelulares y los pluricelulares que no poseen una gran diferenciación celular ni tejidos diferenciados. El actual análisis del material genético ha llevado a la contemporánea clasificación en la que los seres vivos se clasifican en cinco reinos: móneras (organismos procariotas), protistas (eucariotas unicelulares o pluricelulares de estructura muy sencilla), hongos, metafitas (vegetales pluricelulares con tejidos diferenciados) y metazoos (animales pluricelulares). Técnicas para la realización de cultivos puros Los microorganismos no requieren gran espacio para vivir. Suele ser suficiente una placa de Petri o un tubo de ensayo. El recipiente para el cultivo debe estar esterilizado, libre de otros microorganismos vivos que pueden crecer en el cultivo. Tras la introducción de los microorganismos en el cultivo debe mantenerse protegido de cualquier posible contaminación exterior. La fuente principal de contaminación es la atmósfera. Los cultivos se inoculan (inóculo = material utilizado para realizar la siembra con microorganismos) con hilo metálico enrollado (llamado asa) que previamente a su uso ha sido esterilizado en una llama. La esterilización Esterilizar supone dejar un objeto libre de todo organismo vivo. El método de esterilización más usado es el calor. En algunos casos la esterilización se realiza también por medios químicos. Por ejemplo en la industria, las placas de Petri se esterilizan con óxido de etileno. En algunas ocasiones se utiliza la filtración, especialmente de disoluciones termolábiles (que se destruyen con temperaturas altas). Los medios de cultivo Se llaman así a los materiales preparados para que sobre ellos o dentro de ellos crezcan los microorganismos. Un medio de cultivo debe contener cantidades adecuadas de nutrientes para poder permitir un buen crecimiento de los microorganismos. Las concentraciones altas de algunos resultan inhibitorias o incluso tóxicas. Los nutrientes más necesarios son los que aportan elementos tales como nitrógeno, carbono, azufre y fósforo. En algunos casos son necesarios nutrientes especiales. En el caso de los cultivos de diatomeas se requiere nutrientes que aporten silicio. Microorganismos procariotas: Las Bacterias Los microorganismos típicos son las bacterias. Probablemente son los primeros organismos que surgieron en nuestro planeta. Son organismos sencillos, aunque de morfología, tamaño, estructura y metabolismo variado. Tienen una gran capacidad reproductora y de adaptación a distintos medios, por lo que están presentes en todo tipo de ambientes. Suelen vivir aislados o en colonias. En cuanto a su forma se conocen cuatro tipos: bacilos (alargado), espirilos (alargado y curvado), vibrio (curvado) y coco (esférico). Los bacilos pueden aparecer en cadenas lineales (su división se hace en una sola dirección) y los cocos suelen aparecer en cadenas lineales (estreptococos), grupos planos (estafilococos) o en grupos más o menos cúbicos (sarcinas), según su división se realice linealmente o en dos o tres direcciones. Estructura de las bacterias Las bacterias tienen una organización muy compleja, ya que pueden encontrarse en diferentes estructuras. Estos elementos estructurales son: - - - - - - Cápsula: Capa gelatinosa, constituida por polisacáridos, con funciones protectoras frente a sustancias químicas. Pared Celular: Compuesta de mureína, se encuentra dispuesta en varias capas. Su función es mantener la forma de las bacterias. Membrana celular: Similar a la de las células eucariotas en tamaño y composición. Entre las proteínas de la membrana de procariotas se encuentran enzimas implicados en procesos metabólicos como la respiración celular. Mesosomas: Son invaginaciones de la membrana superficial. En ellos se encuentran los complejos enzimáticos respiratorios. Membranas fotosintéticas: En bacterias fotosintéticas, se encuentran sacos membranosos planos que contienen pigmentos fotosintéticos. Flagelos: Son prolongaciones externas largas cuya función es producir el desplazamiento de las bacterias. Pelos o fimbrias: Son prolongaciones tubulares externas de la membrana superficial, a través de la pared celular. Si son largas se denominan pelos, y si son cortas y numerosas se denominan fimbrias. Citoplasma: Es muy uniforme y no se encuentran orgánulos limitados por membranas. Se distingue una zona especial llamada nucleoide o región nuclear, donde se encuentra el DNA circular. Es frecuente encontrar en el citoplasma pequeños fragmentos de DNA llamados plásmidos, que contienen información genética transferible de unas bacterias a otras. En el citoplasma bacteriano también se encuentran gránulos sólidos, llamados inclusiones, de composición diversa, que suelen ser sustancias de reserva o residuos del metabolismo. Muchas bacterias acuáticas suelen tener vesículas gaseosas, que pueden ser muy abundantes, y cuya función es asegurar la flotabilidad. Ribosomas: Son más pequeños que los de las células eucariotas. Endosporas: Son formas bacterianas de vida latente con una cubierta exterior muy dura y resistente. Se forman para lograr sobrevivir en condiciones adversas y al recuperarse las condiciones normales la endospora forma una bacteria de características muy normales. Fisiología bacteriana Las bacterias presentan formas de nutrición muy diversas, pues se conocen tipos metabólicos muy distintos, encontrándose entre ellas autótrofas, heterótrofas y quimiosintéticas. Existen incluso algunas especies que pueden variar del tipo de metabolismo en función de los nutrientes que pueden obtener del medio que les rodea. Los procesos osmóticos tienen gran importancia en bacterias que viven en ambientes de alta salinidad, para regular la concentración de sales en el citoplasma. Reproducción bacteriana Las células bacterianas se reproducen habitualmente por una simple bipartición, sin que ocurran procesos complejos como los de la mitosis de las células eucariotas. En este proceso, tras la duplicación del DNA se produce un tabique en la parte central de la bacteria que la divide en dos. La bipartición, en condiciones de vida adecuadas, se produce muy rápidamente, pues una bacteria puede duplicarse cada veinte minutos. Tipos de bacterias Las bacterias son organismos de características muy diversas, tanto en su morfología, estructura interna, funcionamiento y formas de vida. Las eubacterias son las bacterias más conocidas o típicas. Entre ellas se distinguen distintos tipos, como son, entre otros, las bacterias verdes y púrpuras y cianobacterias, que son bacterias fotosintéticas que se diferencian en el tipo de clorofilas y otros pigmentos fotosintéticos que contienen. Las espiroquetas son las bacterias de cuerpo helicoidal. Los micoplasmas son las eubacterias conocidas más pequeñas. Tienen una estructura muy sencilla, no tienen pared celular y son parásitos de eucariotas. Existen otros tipos de eubacterias. Algunas toman su nombre de alguna característica muy típica como las bacterias fermentadoras (donde se incluyen las que realizan la fermentación láctica y acética), las enterobacterias (que son frecuentes en el tubo digestivo de los animales), las bacterias quimiosintéticas y las bacterias fijadoras de nitrógeno. Los Virus Los virus son un tipo de microorganismos muy simples que infectan células y pueden causar enfermedades. Tienen un tamaño muy pequeño, y su estructura es acelular, ya que no encaja con la estructura celular típica. Características de los Virus Los virus tienen características muy típicas que les hacen muy diferentes a todos los organismos. EL tamaño de los virus es muy pequeño. Su estructura no encaja con la estructura celular, por lo que se suelen considerar como estructuras acelulares. Se denominan viriones las partículas víricas encontradas aisladas. Estos viriones tienen una estructura sencilla, basada en una molécula de ácido nucleico, que puede ser DNA o RNA (nucleoide) envuelta por una cubierta (cápsida) de proteínas, constituida por un número fijo de subunidades proteicas idénticas (llamadas capsómeros) y ordenadas que le dan al virión una morfología externa muy característica. El ciclo de los Virus La vida de un virus alterna dos períodos diferenciados: la vida extracelular en que los viriones no poseen actividad metabólica, y la vida intracelular en la célula hospedante en que se produce la actividad del virus, fundamentalmente su reproducción o replicación. La replicación de los virus requiere que se desarrollen una serie de procesos como son la entrada en el citoplasma de la célula hospedadora del genoma del virus, la síntesis de los componentes del virus, la unión o ensamblaje de los componentes y, finalmente, la liberación al medio exterior de las partículas virales ya maduras. Ciclo lítico Para que un virus penetre en una célula es necesario que entre en contacto y se produzca un reconocimiento por medio un proceso denominado fijación o adsorción. Tras la fijación se produce la penetración del virus en el citoplasma de la célula que se produce de modos diversos. En algunos casos, solo penetra en la célula el genoma vírico. Esto se consigue porque el virus posee lisozimas que perforan la pared celular de las bacterias. En otros, el ácido nucleico penetra encerrado en la cápsida y en el citoplasma se separan ambos quedando libre el ácido nucleico. Su ácido nucleico es capaz de transformar el metabolismo de la célula huésped y orientarlo para producir nuevos virus. Una vez producidos los ácidos nucleicos y las proteínas de la cápsida se produce el ensamblaje de todos estos elementos para producir partículas virales (viriones), que finalmente son liberadas al exterior para así poder infectar nuevas células. En algunos casos se liberan los viriones a través de la membrana citoplasmática sin que aparentemente se produzca daño para la célula huésped. Pero en la mayoría de los casos la liberación de los viriones se produce porque las partículas virales provocan la lisis de la célula infectada y, por consiguiente, su muerte. A este tipo de replicación de virus se le denomina ciclo lítico. El ciclo lisogénico En ocasiones los virus al parasitar una célula no producen la lisis de la misma. En estos casos el ácido nucleico de la célula donde puede permanecer varias generaciones en forma latente hasta que en ciertas condiciones se produce un ciclo lítico y se liberan los viriones. Estos virus se denominan virus atenuados o profagos, y el proceso que se produce se denomina lisogenia, y la célula huésped célula lisogénica. Este tipo de infección lisogénica parece ser un mecanismo de enriquecimiento genético de los seres vivos pues los virus parecen funcionar en ocasiones como transportadores de genes, que al infectar una célula pueden permanecer como profagos incorporados permanentes, sin producirse nunca lisis celular. OTROS MICROORGANISMOS: ALGAS, PROTOZOOS, HONGOS, LEVADURAS… Entre los microorganismos, aparte de las bacterias y virus, se incluyen otros organismos como son algunas algas, protozoos y hongos. Algas Las algas son organismos acuáticos de vida libre, que viven tanto en el agua dulce como en la salada. Poseen cloroplastos con clorofila y otros pigmentos, por lo que realizan la fotosíntesis. Las más comúnmente observadas son pluricelulares y se clasifican en tres grandes grupos como son las algas verdes, algas pardas y algas rojas. Estas pluricelulares pueden llegar a alcanzar muchos metros de longitud y tienen aspecto de planta, pero su organización celular es muy simple. La mayoría son unicelulares flotantes, como las diatomeas o los dinoflagelados, que viven en las aguas superficiales de todos los océanos y lagos. Algunas son terrestres y viven en zonas húmedas en el suelo y sobre la corteza de los árboles. Hay algas que viven en simbiosis con invertebrados marinos, esponjas, corales, etc. Otras tienen relaciones muy estables con hongos formando complejas colonias conocidas como líquenes. Entre las variadas algas unicelulares las más sencillas son las flageladas fotosintéticas. Un ejemplo típico es el género Euglena, que tiene un orgánulo característico “la mancha ocular” con funciones fotorreceptoras y que controla el movimiento de la célula. Existen también algunas algas unicelulares no flageadas de las que las más características son las algas verdes desmidiáceas y las diatomeas. Las desmidiáceas son algas verdes unicelulares, con pared celular de celulosa, cuyas células planas tienen simetría bilateral. Las diatomeas tienen paredes silíceas de morfología compleja y curiosa. Pueden acumularse y conservarse, constituyendo las rocas llamadas tierras de diatomeas. Protozoos Son microorganismos eucariotas unicelulares que no poseen paredes celulares. Suelen ser incoloros y móviles. Se diferencian por su tamaño y por ser eucarióticos, de las algas por su falta de clorofila, de las levaduras y los hongos por su movilidad y falta de pared celular, y de los mohos viscosos por la no producción de cuerpos fructíferos. Son heterótrofos y se alimentan de otros organismos fagocitándolos, o bien de partículas orgánicas. Se encuentran en gran variedad de ambientes, generalmente en agua dulce o marina. Algunos son parásitos de animales y del ser humano, y también aparecen sobre el suelo o cortezas de árboles. El tipo de movimiento es una característica para clasificar estos organismos: flagelados, con flagelos; rizópodos, con movimiento ameboide: ciliados, con cilios; esporozoos, que son parásitos obligados y generalmente inmóviles. Hongos Son organismos que no poseen clorofila y, por lo tanto, son heterótrofos. Tienen células con mitocondrias y vacuolas, y una pared celular quitinosa. Generalmente son multinucleados y filamentosos. La unidad básica de su estructura en forma de hilo o tubo se denomina hifa. El conjunto ramificado de hifas se denomina micelio. Los tipos principales de hongos son los mohos y las levaduras, entre los que se encuentran muchos que son microscópicos, y las setas, que son macroorganismos. Los mohos Son hongos filamentosos, que crecen sobre el pan viejo, la fruta, el queso… A partir del micelio crecen hifas aéreas en las que se desarrollan las esporas llamadas conidios. Los conidios son muy resistentes a la desecación y suelen tener un color negro, azul verdoso… Las levaduras Son hongos unicelulares, en la mayoría de los casos ascomicetos. Producen esporas por reproducción sexual dentro de un saco cerrado, llamado asca. Las células de levadura son esféricas, ovales o cilíndricas, y se dividen por gemación. Mohos viscosos También son denominados hongos mucosos. Viven generalmente sobre los troncos en descomposición de los bosques húmedos. Son conocidos con el nombre de myxomicetes. La organización de su organismo se denomina plasmodio, que es una masa de citoplasma que posee muchos núcleos y carece de pared externa rígida. METABOLISMO DE MICROORGANISMOS Una de las formas más comunes del metabolismo en los microorganismos bacterianos es la forma de autotrofía de la quimiosíntesis. Estos organismos son capaces de producir la fosforilación de ADP a ATP y de lograr poder reductor para sintetizar compuestos orgánicos utilizando energía procedente de la oxidación de compuestos inorgánicos. SIMBIOSIS Y MICROORGANISMOS La simbiosis es la asociación continuada y en ocasiones muy íntima de distintos tipos de seres vivos, en la que los simbiontes logran una ventaja para su vida. El estudio de las diferentes formas de simbiosis entre microorganismos ha revelado que en estas formas de vida los simbiontes obtienen distintos tipos de beneficio. Un ejemplo es la protección de los microorganismos: tanto los endosimbiontes como los ectosimbiontes que viven en cavidades corporales están protegidos de la desecación o, en el caso de los que viven en animales homeotermos, de los cambios de temperatura. También parecen ofrecer una cierta protección a sus socios, siendo un ejemplo importante la protección de que parecen disponer los vertebrados gracias a los microorganismos que viven en ellos frente a la invasión de microorganismos patógenos. BIOTECNOLOGÍA ¿Qué es la biotecnología? Historia de la biotecnología. El término biotecnología fue creado en los años 70 en Leeds (Reino Unido). La biotecnología es la aplicación de organismos completos o sistemas biológicos, por ejemplo células o estructuras subcelulares, para la producción industrial de materiales útiles a gran escala. Los organismos más utilizados en los procesos biotecnológicos son los microorganismos por varias razones como el conocimiento profundo de su cultivo y manipulación a gran escala, sus características metabólicas, y porque la técnicas de ingeniería genética que se están desarrollando estos últimos años, que permiten introducir o modificar genes en las células modificando el funcionamiento de las mismas, se han desarrollado en gran medida en los microorganismos, pues son más fáciles de manipular que los organismos pluricelulares. Aunque la biotecnología se considera una nueva tecnología, podemos decir que tiene muchos años de historia, y que a lo largo de la misma ha pasado por varias etapas. Por ejemplo, el uso de microorganismos de modelo artesanal y empírico, en la que se controlaban algunos procesos biológicos sin conocer las causas de los mismos. La base científica de la biotecnología más moderna, denominada ingeniería genética, es la genética molecular, que se ha desarrollado a partir del conocimiento de la estructura de los ácidos nucleicos y de otros descubrimientos posteriores, como son el del código genético, de la DNA ligasa, de los enzimas de restricción… Biotecnología tradicional Durante muchos siglos se usaron los microorganismos de un modo empírico, pero sin llegar a conocer cómo y por qué se realizaban dichos procesos. Así se logró controlar ciertos procesos de producción de alimentos, como el pan o el yogurt. Hoy en día la fabricación de estos productos que compramos se fabrican masivamente en industrias y en muchos casos, incluso sin usar microorganismos, pues éstos han sido sustituidos por diversas sustancias que producen efectos similares. Biotecnología microbiana industrial Los microorganismos utilizados industrialmente son microorganismos que han sido aislados de ambientes naturales y a lo largo de los años han sido seleccionados y perfeccionados, a fin de mejorar su capacidad productiva. Esto origina que las cepas usadas en la industria tengan una alta especialización metabólica, sobreviviendo solamente en las especiales condiciones en que se mantienen en la industria. Los productos microbianos industriales que se producen suelen ser células microbianas, enzimas o metabolitos microbianos. Los objetivos de este tipo de producción son variados: farmacéutico (antibióticos); agrícola, alimentario (algunos aminoácidos) o el químico industrial. Asimismo se han usado los microorganismos como agentes ambientales para la degradación de compuestos orgánicos de desecho o incluso de productos tóxicos para la producción de productos energéticos (metano) o de alimentos proteicos. Ingeniería genética La ingeniería genética es el tipo de biotecnología más de moda hoy en día. Su inicio se sitúa a finales de los años 60 y a principios de los 70. Se denominan así las técnicas basadas en la recombinación del DNA, utilizadas para modificar el genoma de los organismos, bien introduciendo genes ajenos o extranjeros, bien modificando algunos de los genes propios. Ingeniería genética en ganadería La mejora de los animales criados en ganadería puede lograrse por técnicas de ingeniería genética. Un ejemplo de ello está siendo desarrollado en Australia para intentar controlar las garrapatas del ganado lanar. Ingeniería genética y medicina La ingeniería genética ha encontrado grandes aplicaciones en la medicina. Se ha logrado obtener insulina humana producida por bacterias transformadas por la inserción del gen que codifica dicha hormona, se aplica la hormona del crecimiento producida por estos procesos a personas con problemas de crecimiento, se está logrando incrementar o mejorar la producción de antibióticos y vacunas, etc. Uno de los productos de ingeniería genética de gran interés son los denominados anticuerpos monoclonales. Se han logrado clones de células producidas tras la fusión de células productoras de anticuerpos y células cancerosas. De esta fusión se desarrollan mielomas, que tienen la capacidad de crecer indefinidamente en el laboratorio. También se está tratando de desarrollar la terapia genética para curar enfermedades de origen genético por medio de la modificación de genes patógenos en las personas o estimulando la actividad inmunológica. INMUNOLOGÍA La inmunidad es el mecanismo por el cual, el organismo reconoce todas sus células estableciendo un equilibrio entre ellas y rechazando las células que le son extrañas o que rompen el equilibrio. Diferenciamos los siguientes mecanismos de inmunidad: A) B) - NO ESPECÍFICOS: Barreras anatómicas y fisiológicas. Respuesta inflamatoria. ESPECÍFICOS: Respuesta inmunológica. BARRERAS DEL ORGANISMO ANTE LA INFECCIÓN El organismo humano se encuentra rodeado de microorganismos. Frente a estos organismos poseemos una serie de barreras que impiden su paso: anatómicas, químicas, bioquímicas y ecológicas. A) Barreras anatómicas: La piel con su epitelio queratinizado es una barrera inexpugnable mientras que se mantenga sin lesiones. Las mucosas presentan epitelios vibrátiles y secreciones para impedir la penetración de los microorganismos. B) Barreras químicas: El sudor, fluido con un pH ácido, que impide el crecimiento de bacterias, o el jugo gástrico que elimina las bacterias introducidas junto con el alimento. C) Barreras bioquímicas: Las lágrimas, la saliva, contienen lisozima, una enzima antibacteriana. En la mujer, las secreciones vaginales, y en el hombre, la espermina, tienen propiedades antimicrobianas. D) Barreras ecológicas: Son el conjunto de condiciones que favorecen el crecimiento de microorganismos no patógenos, los cuales inhiben el crecimiento de los patógenos. LA INFLAMACIÓN Al introducirse microorganismos en los tejidos, se activan una serie de mecanismos que culminan con la actividad de granulocitos neutrófilos y macrófagos que fagocitan a los microorganismos, conteniendo así su reproducción y destruyéndolos. Esta respuesta es activada por sustancias que segregan las células afectadas por una herida. Los neutrófilos y los monocitos son atraídos por quimiotactismo a la zona dañada y pasan atravesando los capilares por medio de movimientos ameboides. Los neutrófilos, que tienen una capacidad fagocitaria limitada, son los primeros en actuar. Los monocitos, que acuden posteriormente, una vez en el tejido, se transforman en macrófagos. Alteraciones locales ayudan a la acción de estas células: el aumento del flujo sanguíneo en la zona y la dilatación de los vasos sanguíneos facilitan la salida al medio celular de plasma y de los fagocitos, proceso que recibe el nombre de inflamación. La formación de pus, constituido por células muertas de la zona afectada, bacterias y neutrófilos muertos, es consecuencia de esta acción. La fagocitosis puede ser insuficiente en algunos casos para eliminar los microorganismos introducidos, deteniéndose en este punto la acción defensiva del organismo. En estos casos se hace necesaria la respuesta inmunológica. LA RESPUESTA INMUNOLÓGICA La respuesta inmunológica es el mecanismo más elaborado que poseemos los vertebrados para defendernos de sustancias ajenas o que reconocemos como ajenas (antígenos). Los linfocitos, otro tipo de leucocitos, son los responsables de la reacción inmunológica. Se conocen dos tipos de linfocitos: B y T. Los linfocitos B desarrollan la respuesta inmunológica humoral, caracterizada por la síntesis de anticuerpos que liberan al medio interno reaccionando con los antígenos. Los linfocitos T actúan directamente uniéndose a células ajenas, es decir, ayudan en la respuesta inmunológica celular. Ambas respuestas actúan de forma coordinada. La respuesta inmunológica se caracteriza por tres propiedades bien definidas: - Especificidad de actuación sobre el elemento que ha provocado la respuesta. Memoria. Propiedad de desencadenar una respuesta más rápida y eficaz tras el contacto con el antígeno. El reconocimiento de lo propio. CONCEPTO DE ANTÍGENO Llamamos antígeno a cualquier sustancia que, introducida en el organismo, sea capaz de provocar la respuesta inmunológica. Tanto microorganismos, como proteínas, glúcidos… Los antígenos se encuentran en el organismo, ya sea libres en el medio interno o formando parte de una estructura celular. En los antígenos, no toda la molécula interviene en la inducción de anticuerpos. A la parte del antígeno que se une con el receptor de los linfocitos y que desencadena la respuesta inmunológica se le conoce como determinante antigénico o epítopo. Algunas moléculas pequeñas son reconocidas por los anticuerpos y se unen a ellas pero no son capaces de activar la respuesta inmunológica. Estas moléculas se conocen con el nombre de haptenos. EL SISTEMA INMUNOLÓGICO El sistema inmunológico está constituido por una serie de órganos aislados y de células: los linfocitos y los macrófagos. MACRÓFAGOS Los macrófagos son células que provienen de la diferenciación de los monocitos, de gran tamaño. Son capaces de fagocitar microorganismos y restos de células. Participan tanto en la respuesta inflamatoria como en la respuesta inmune. Intervienen fagocitando sustancias extrañas y cooperando con los linfocitos en la presentación del antígeno. LINFOCITOS Los linfocitos se originan a partir de células progenitoras situadas en el hígado durante el estado fetal y en la médula ósea después del nacimiento. En el organismo coexisten dos tipos de linfocitos: T y B, llamados así porque se diferencian y maduran, unos en el timo (linfocitos T), y otros en la médula ósea (linfocitos B). ÓRGANOS DEL SISTEMA INMUNOLÓGICO Los órganos del sistema inmunológico se encuentran distribuidos por el cuerpo y relacionados entre sí a través de la linfa y el sistema circulatorio. Se clasifican en primarios o secundarios según sean lugar de síntesis y maduración de linfocitos o lugar de circulación, acumulación y actuación. Órganos linfoides primarios Son los órganos donde se originan y maduran los linfocitos. La médula es el lugar de origen de todos los linfocitos y de maduración de los linfocitos B. El timo tiene un máximo desarrollo en el feto, y en él se maduran los linfocitos T. Órganos linfoides secundarios Ganglios, vasos linfáticos, Placas de Peyer, apéndice… Estos órganos son el lugar de acumulación de las células inmunológicas y es en los ganglios linfáticos y en el bazo donde se produce la reacción inmunológica y la acción fagocitaria. CONCEPTO DE ANTICUERPO Los anticuerpos son glucoproteínas (Inmunoglobulinas Ig) sintetizadas y segregadas por las células plasmáticas (linfocitos B activados). Son moléculas de estructura muy compleja. En ellos se da una disposición básica en forma de Y, constituida por cuatro cadenas proteicas, 2 ligeras (L) y dos pesadas (H), siendo en un mismo anticuerpo idénticas las cuatro entre las dos ligeras y las dos pesadas. Estas cadenas permanecen ligadas entre sí por fuertes uniones llamadas puentes disulfuro. En los dos tipos de cadenas se encuentran zonas constantes, con secuencias de aminoácidos idénticas de una molécula a otra, y zonas variables distintas de un anticuerpo a otro. Las regiones hipervariables son la zona de reconocimiento del antígeno o sitio activo de la molécula. La zona constante es la que determina la funcionalidad del anticuerpo. Se distinguen en ella de 3 a 4 zonas. Una de estas zonas es el sitio de unión del complemento. Otra de las zonas es la que facilita su unión a los fagocitos. Clases de inmunoglobulinas - INMUNOGLOBULINA G: Es la más abundante. Se une rápidamente con macrófagos y neutrófilos, provocando la destrucción del microorganismo. INMUNOGLOBULINA A: Corresponde al 13% del total de inmunoglobulinas. Actúa protegiendo la superficie corporal y conductos secretores. INMUNOGLOBULINA M: Representa el 6% del total de inmunoglobulinas. Se manifiesta en la respuesta primaria activando el complemento. INMUNOGLOBULINA D: Aparece en muy baja concentración (1%). Su función está relacionada con la activación de los linfocitos B. INMUNOGLOBULINA E: Se encuentra en concentraciones muy bajas en secreciones al exterior (0,002%). RESPUESTA INMUNOLÓGICA HUMORAL Los linfocitos B son los responsables de la respuesta inmunológica humoral, caracterizada por la producción de anticuerpos libres que circulan por la sangre y la linfa y neutralizan al antígeno. Maduración y diferenciación de los linfocitos B La maduración de los linfocitos B se produce en la médula ósea y es un proceso previo e independiente a la presencia del antígeno. Consiste en la aparición sucesiva de inmunoglobulinas fijas a la membrana. Estas inmunoglobulinas tienen una estructura idéntica a la de los anticuerpos que fabricará la célula, aunque se encuentran fijas a la membrana celular del linfocito por la zona constante de la molécula sobresaliendo la zona variable con sus dos sitios activos. Un determinado linfocito B posee todos sus receptores de membrana idénticos, por lo que será capaz de reconocer a un único determinante antigénico. Llamamos clon a toda población de linfocitos que posee un mismo receptor. Tras la maduración, los linfocitos son capaces de reconocer el antígeno y dar la respuesta humoral. Proceso de Respuesta Humoral La respuesta humoral se desarrolla tras el encuentro del antígeno con el linfocito B; el proceso lo podemos diferenciar en varias fases: A) FASE DE SELECCIÓN Y ACTIVACIÓN: Esta fase se inicia con el reconocimiento del antígeno, y selección de un clon de linfocitos. El clon de linfocitos se unirá a él, quedará seleccionado, y empezarán a producirse en el linfocito una serie de cambios que lo transformarán en célula productora de anticuerpos. Este modelo explicativo de la selección de un clon de linfocitos B se conoce como teoría de selección clonal. La selección del clon de linfocitos no es suficiente en la mayoría de los casos para desencadenar la respuesta humoral; es necesaria la cooperación de los linfocitos T 4, un subtipo de linfocitos T, y los macrófagos para la activación del clon seleccionado. Los linfocitos T4 se unen directamente a los linfocitos B o a los macrófagos, con lo que se activan iniciando la síntesis de unas moléculas químicas, las interleucinas, que producen una segunda señal necesaria para que siga adelante la respuesta humoral. B) FASE DE PROLIFERACIÓN: En esta fase se divide activamente el clon de linfocitos activados, diferenciándose en dos clases: células plasmáticas, especializadas en la síntesis de anticuerpos y células de memoria, que permanecen en el medio interno por largos períodos de tiempo y darán lugar a la respuesta inmunológica sólo si se encuentran con el mismo antígeno. Los anticuerpos así sintetizados circulan libremente por la sangre, linfa y órganos linfáticos secundarios hasta unirse selectivamente a los antígenos libres o de membrana. C) FASE EFECTORA: También conocida como unión antígeno-anticuerpo, que desemboca en la destrucción del antígeno, lo que ocurre gracias al complemento y los macrófagos. Los anticuerpos liberados al medio interno interactúan con el antígeno originando respuestas diferentes que conducen a su destrucción por distintos medios: 1) Inmovilización del antígeno, lo que conduce a la precipitación de este. 2) Activación de células fagocitarias, uniéndose específicamente a determinados receptores de fagocitos (macrófagos y granulocitos) lo que facilita el trabajo de estas células. 3) Acción lítica, proceso que se desencadena al activar el complemento, verdadero conjunto enzimático que produce la lisis de las células a partir de una reacción en cadena que desemboca en la formación de perforinas (enzimas líticas). RESPUESTA INMUNOLÓGICA CELULAR Los linfocitos T actúan directamente contra antígenos unidos al células, concretamente sobre células del organismo que han sido modificadas por una infección o una mutación o células ajenas al individuo. A esta forma de activación se le denomina respuesta celular. Los linfocitos T reconocen a las células infectadas o modificadas del organismo y las diferencian de las propias. Este reconocimiento se realiza en el período fetal durante el proceso de maduración. Maduración y diferenciación de los linfocitos T La maduración se desarrolla incorporando a la membrana unos receptores proteicos que están formados por solo dos cadenas proteicas y tienen un único sitio activo. Estas proteínas de membrana solo reconocen a determinantes antigénicos que se encuentran en la membrana de las células del organismo, de ahí que estos linfocitos actúen por acción directa sobre las células. Se ha de destacar el papel que en la respuesta celular juega un tipo de receptores constituido por las proteínas codificadas por un grupo de genes, el complejo principal de histocompatibilidad (CMH). Estos receptores que se encuentran en todas las células nucleadas del organismo. Desempeñan un papel importante en el origen y desarrollo de la respuesta celular, ya que los linfocitos T sólo pueden reconocer a los antígenos, y siempre y cuando estén asociados a dichas proteínas. Estos receptores proteicos presentan una gran variedad, de forma que no existen dos personas con el mismo tipo de glucoproteínas, excepto los gemelos univitelinos. Gracias a esta especificidad pueden desencadenar la respuesta celular. Tipos de linfocitos T Se diferencian los siguientes tipos de linfocitos T: a) Linfocitos citotóxicos (T8), se unen a los antígenos presentados por células modificadas o extrañas al organismo y actúan destruyendo dichas células lisándolas. b) Linfocitos auxiliares (T4), que reconocen los antígenos presentados por células del sistema inmunológico, y colaboran en la activación y amplificación de la respuesta inmunológica. c) Linfocitos supresores, que están relacionados con la supresión de la respuesta inmunológica. Proceso de respuesta celular Al igual que en la respuesta humoral, es el antígeno el que desencadena la respuesta celular al unirse a un receptor específico y seleccionar un clon de células. El proceso se puede esquematizar de la siguiente forma: A) FASE DE SELECCIÓN Y ACTIVACIÓN: Comienza con el reconocimiento de los determinantes antigénicos unidos a las células. Los linfocitos T sólo reconocen a antígenos presentes en la membrana de células y que estén asociados a las proteínas del CMH. La presentación de los antígenos por los macrófagos es el proceso que desencadena la selección de un clon de linfocitos T4 auxiliares. Los macrófagos fagocitan antígenos y algunos péptidos degradados son llevados hasta la membrana celular asociados a las proteínas del CMH, donde serán reconocidos por receptores específicos de los linfocitos T4. Por un proceso semejante, las células cancerosas o atacadas por un virus, o ajenas al organismo muestran determinantes antigénicos unidos a proteínas del CMH en su membrana, induciendo la unión de un linfocito T8 citotóxico. Los linfocitos T4 auxiliares seleccionados inician la síntesis de interleucinas que actúan como mediadores químicos. B) FASE DE PROLIFERACIÓN: La activación de los linfocitos T4 auxiliares conduce a la división por mitosis del clon de células seleccionado originándose linfocitos T 4 de memoria, que permanecerán largo tiempo en la sangre y linfa, y linfocitos T 4, que segregarán interleucinas que a su vez activarán al clon de linfocitos T8 citotóxicos seleccionados. Éstos, a su vez, al activarse se dividirán en T 8 de memoria y T8 efectores. C) FASE EFECTORA: También conocida como fase lítica, en la que los linfocitos T 8 efectores producen la lisis de las células reconocidas como ajenas. Uno de los mecanismos de lisis conocido es el de choque osmótico, producido por acción de las perforinas, un tipo de enzimas que inducen poros en la membrana de las células, por los que se introduce el agua. Los linfocitos T8, aunque reconocen y se unen a las células modificadas por el antígeno, no desencadenan su acción lítica hasta que no son activados por los T 4 auxiliares. IMAGEN↑ MEMORIA INMUNOLÓGICA Una de las características del sistema inmunológico es su capacidad para recordar, es decir, tras la exposición a un virus o bacteria la capacidad que tiene un organismo para adquirir inmunidad por largos períodos de tiempo frente a dicho organismo. Al inocular el antígeno, al cabo de unos 5 días aproximadamente se produce la respuesta inmunológica primaria. Si el organismo es puesto de nuevo en contacto con el mismo antígeno, la respuesta en esta ocasión será mucho más rápida, intensa y duradera, es la respuesta inmunológica secundaria. Esta memoria del sistema inmunológico es debida a la síntesis de células de memoria, producida en la fase de proliferación y que no intervienen directamente en el proceso infeccioso permaneciendo largo tiempo en el sistema sanguíneo y linfático. DISFUNCIONES DEL SISTEMA INMUNOLÓGICO ALERGIA Y ANAFILAXIS La alergia es una reacción inflamatoria generada como consecuencia de un estado de hipersensibilidad inmunológica. Son enfermedades que se manifiestan de distinta forma: asma, rinitis… Este trastorno del sistema inmunológico se produce al actuar las células inmunológicas sobre sustancias como el polen, ácaros… El proceso de la alergia se desarrolla en dos fases: a) Fase de sensibilización, que se inicia al introducirse el alérgeno por primera vez en el organismo. El contacto con las células inmunológicas produce la activación de un clon de linfocitos B, que se transforman en células secretoras de inmunoglobulinas E. Estas inmunoglobulinas se unen a los receptores de los mastocitos, que son células presentes en el tejido conjuntivo y mucosas, que presentan en su citoplasma gran cantidad de histamina y otros mediadores químicos. En esta fase no se producen síntomas aparentes de sensibilización. b) Fase de activación de los mastocitos, que se desencadena al contactar los mastocitos con el alérgeno por segunda vez. Esto provoca modificaciones en la permeabilidad celular, liberándose al medio interno la histamina, mediador químico que atrae a los fagocitos desencadenándose la respuesta inflamatoria o alérgica. En algunos casos esta respuesta alcanza una gran intensidad produciendo caída de la presión sanguínea, contracción de los bronquiolos, llegando incluso a provocar la muerte del individuo. Se denomina anafilaxis a este tipo de respuesta. ENFERMEDADES AUTOINMUNES La autoinmunidad es el ataque que sufren las células o moléculas del propio organismo por parte de células inmunológicas. Las enfermedades autoinmunes son consecuencia de un mal funcionamiento del sistema inmunológico que, incapaz de reconocer como propias las células del organismo, las destruye. INMUNODEFICIENCIAS La inmunodeficiencia es la insuficiencia de una o varias funciones del sistema inmunológico con consecuencias patológicas. Esta disfunción es consecuencia de anomalías en órganos productores de linfocitos B o linfocitos T, o de ambos. Algunas de estas deficiencias son congénitas, otras, por el contrario, son adquiridas por diferentes motivos: malnutrición, insuficiencia en la ingestión de proteínas… EL SIDA El Síndrome de la Inmunodeficiencia Adquirida (SIDA), es una enfermedad causada por el Virus de la Inmunodeficiencia Humana (VIH). Este es un virus de forma casi esférica con una cubierta externa constituida por una bicapa lipídica con abundantes protuberancias de origen proteico. Estas glucoproteínas pueden unirse estrechamente con las proteínas de los receptores T4. Esta unión específica es la que posibilita la infección preferente de los linfocitos T auxiliares. Actuación del virus del SIDA El VIH ataca preferentemente a los linfocitos T auxiliares, con lo que se produce a la larga el debilitamiento de la respuesta inmunológica del individuo. La introducción del VIH en el organismo desencadena en un primer momento la respuesta humoral al activar los linfocitos B y la consecuente producción de anticuerpos; sin embargo, esta respuesta no es suficiente para neutralizar al virus. La detección en la sangre del paciente de anticuerpos del virus es la base de la prueba serológica para la detección del SIDA, denominando seropositiva a la persona cuya sangre contiene dichos anticuerpos. El virus penetra en los linfocitos T4 fijándose a la membrana del linfocito y penetrando en el citosol de la célula huésped. Una vez en el interior, el RNA del virus se transcribe en una doble hélice de DNA por acción de la transcriptasa inversa. Este DNA se integra en el cromosoma del linfocito, y ya en el cromosoma del huésped, el virus aprovecha para multiplicarse y producir la lisis celular, liberándose nuevos viriones que seguirán el ciclo de infección de nuevas células. La reducción de los linfocitos T4 debilita las defensas inmunológicas del paciente y con ello aparecen diferentes enfermedades, llamadas oportunistas, ya que en condiciones normales el organismo podría superarlas. En determinadas ocasiones el material genético del virus se integra en el cromosoma del huésped pero no induce la producción de nuevos virus, permaneciendo latente (provirus) en el interior de las células infectadas. Estas personas son portadoras de la enfermedad aunque no la manifiesten. AYUDAS AL SISTEMA INMUNOLÓGICO VACUNAS La vacunación consiste en la inoculación de gérmenes defectuosos, muertos o afines al microorganismo infeccioso, ANTES DE CONTRAER LA ENFERMEDAD, para inducir la respuesta inmunológica característica. SUEROS La sueroterapia consiste en la inoculación al individuo, CON FINES CURATIVOS, de un suero que contiene anticuerpos específicos para la enfermedad que el paciente sufre, o como prevención en situaciones en que el riesgo de infección es muy alto. El suero se obtiene ya sea a partir de seres humanos, animales… Anticuerpos Monoclonales Un anticuerpo monoclonal es un anticuerpo homogéneo producido por una célula híbrida producto de la fusión de un clon de linfocitos B descendiente de una sola y única célula madre, y una célula plasmática tumoral. Son anticuerpos idénticos porque son producidos por un solo tipo de célula del sistema inmune, es decir, todos los clones proceden de una sola célula madre. Es posible producir anticuerpos monoclonales que se unan específicamente con cualquier molécula con carácter antigénico. Este fenómeno es de gran utilidad en bioquímica, biología molecular y medicina. TRASPLANTES Los trasplantes son procedimientos quirúrgicos mediante los cuales se transfiere un órgano o tejido de un área determinada de una persona (donante), a otra de otra persona (receptor). Los riesgos del trasplante derivan de: a) Reacciones de rechazo por parte del huésped, al ser reconocido el implante como extraño por el sistema inmunológico. Para evitarlo se somete previamente al receptor a irradiación para destruir sus células inmunológicas. b) Reacciones del tejido trasplantado contra el huésped, debido a los linfocitos T y B que pueden provocar reacciones inmunológicas contra los antígenos del receptor. Es necesario eliminar los linfocitos T y B del órgano a trasplantar. La destrucción se realiza con anticuerpos antilinfocitos. c) Posibilidad de infección durante el período posterior al trasplante. La ausencia de defensas del receptor, debido al tratamiento recibido, hasta que se recupera la respuesta inmunológica crea un serio peligro de infección que se trata con sueroterapia preventiva. APUNTES ADICIONALES TÉCNICAS DE IDENTIFICACIÓN DE DNA Transgenes Un transgen es un gen que se moviliza o se transfiere entre dos organismos distintos o líneas de una manera que no sea la reproducción sexual, para obtener una producción masiva del gen. Las causas naturales de dichas transferencias, a veces denominadas lateral u horizontal, incluyen transportar los genes por medio de bacterias o virus que pueden transferir el DNA entre distintas especies (plásmidos). Método de Sanger El método dideoxi de secuenciación ideado por Sanger está basado en el empleo de didesoxinucleótidos que carecen de uno de los grupos hidroxilo, de manera que cuando uno de estos nucleótidos se incorpora a una cadena de ADN en crecimiento, está cadena no puede continuar elongándose ya que la ADN polimerasa necesita un extremo 3’ OH para añadir el siguiente nucleótido y el desosinucleótido incorporado carece de este grupo hidroxilo. Se aísla y se clona el ADN que se desea secuenciar, este ADN se desnaturaliza y se emplea una sola hélice en la secuenciación. En la secuenciación se utiliza un cebador o “primer” marcado radiactivamente que suministra el extremo 3’OH que necesita la ADN polimerasa. Se preparan cuatro tubos de reacción, cada uno con el ADN molde de hélice sencilla que se desea secuenciar, con ADN polimerasa, con el cebador marcado y con los cuatro nucleótidos trifosfato. A cada tubo se le añade una pequeña proporción de un didesoxinucleótido trifosfato, un tubo con ddATP, otro con ddTTP, el tercero con ddGTP y el cuarto con ddCTP. En cada uno de estos tubos se producirán cadenas de ADN de distintas longitudes, terminando todas en el lugar en el que se incorporó el dideoxi correspondiente añadido al tubo. Posteriormente, estas piezas de ADN se separan mediante electroforesis vertical en geles de acrilamida. Las piezas más pequeñas migran más rápidamente que las grandes y la secuencia se puede leer directamente sobre el gel de acrilamida. En el siguiente esquema se indican los resultados que obtendríamos al realizar la autorradiografía del gel de secuenciación: PCR La reacción en cadena de la polimerasa, conocida como PCR por sus siglas en inglés (polymerase chain reaction), es una técnica de biología molecular desarrollada en 1986. Su objetivo es, mediante el uso de diferentes elementos químicos (cebadores, soluciones tampón…), obtener un gran número de copias de un fragmento de ADN particular, partiendo de un mínimo; en teoría basta partir de una única copia de ese fragmento original, o molde. Esta técnica sirve para amplificar un fragmento de ADN; su utilidad es que tras la amplificación resulta mucho más fácil identificar con una muy alta probabilidad, virus o bacterias causantes de una enfermedad, identificar personas (cadáveres) o hacer investigación científica sobre el ADN amplificado. Estos usos derivados de la amplificación han hecho que se convierta en una técnica muy extendida, sobre todo en el ámbito de la investigación forense, con el consiguiente abaratamiento del equipo necesario para llevar a cabo dicha técnica. PASTEURIZACIÓN La pasteurización es el proceso de calentamiento de líquidos (generalmente alimentos) con el objeto de la reducción de los elementos patógenos, tales como bacterias, protozoos, mohos y levaduras, etc que puedan existir. El avance científico de Pasteur mejoró la calidad de vida al permitir que productos como la leche pudieran transportarse sin descomponerse. En la pasteurización no es el objetivo primordial la "eliminación de los elementos patógenos" sino la disminución de sus poblaciones, hasta niveles que no causen intoxicaciones alimentarias (asumiendo que el producto pasteurizado se ha refrigerado correctamente y que se consume antes de la fecha de caducidad).
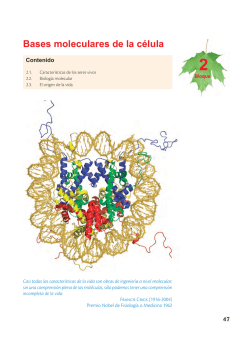


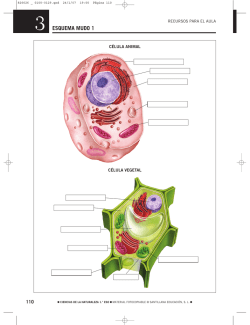




© Copyright 2026