
Descargar
-2 00 0 00 -16 - 20 - 20 0 0 00 -1600 -2000 -2 0 -1 6 0 0 00 -2000 -3 0 0 0 00 -2000 -16 -2 00 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 0 -3 000 -3 0 00 -20 00 60 40 00 0 - 30 00 -2 -40 -1 00 - -2 0 0 0 -16 00 -5 0 -10 -10 -10 1 -10 -10 -200 000 -10 -100-10 1-10 -1 --10 -1-1-10 -1 -10 -100 0 -1 6 00 -10 -5 - 4 00 0 - 16 -100 00 -2 0 00 -1 0 -2 0 0 0 00 - 2 00 0 -2 0 0 0 0 -1 00 0 -1 00 0 -1 0 -3 00 -4 -2000 -160 0 000 -1000 -1 -40 00 4 00 0 -4 - 40 0 0 00 00 4 000 00 0 - 00 -4 00 0 -2 00 0 0 -3 0 0 0 00 -40 -2 - 10 00 00 0 00 0 -2 0 -10 -3 00 -200 -5 -5 -5-5-5-10 - 16 0 -5 5 -5 -5--5-5 -5 -10 -1000 MAR CARIBE -5-5 -5 -10 -5 -5 -5-5 -1 -10 -5--5 0 10 -10 00 -5-5-5-1 -10 -10 -10 1-5 -5 -5-10 -10 -5 -10 -1-10 -5 -5 -5 -5 -5 -5 0-50-5 -5 -5-5 -5 -5-5 -5 -5 -10 -10 -5 -10 -5 -10 -10 -5 -5-5 -50-5 -5 -5 -5 -5-50 -5-10 -1 -1 -5-1-50 -10 -1 -5 0-5-5 0 -5 -5-5 -10 -10 -5 -5-5 -10 -10 -5 -5 -5 0-1-5 -1-5- -10-50 5 - 5-5-5-5 0 00 -1 0 0 0 -1 0 -20 00 0 -16 0 -1 1 60 0 -50 - 0 00 -1 0 -1600 60 - 30 00 Manaure Santa Marta D.T.C.H. Piojó Dibulla Riohacha -10 -5 -10 -5 PuertoSitioPueblo Ciénaga NuevoViejo Colombia LA GUAJIRA -5 -5 -10 -5 -3 0 00 -10 ATLÁNTICO Cartagena de Indias MAGDALENA -30 00 BOLÍVAR CESÁR San Onofre Santiago de Tolú San Antero Moñitos Puerto Escondido Los Córdobas SUCRE ArboletesCÓRDOBA Necoclí Acandí Turbo Unguía Apartadó Juradó ANTIOQUIA -30 0 00 0 -4 -300 0 Bahía Solano 0 -3000 -3 0 -4 00 -3 - 3 000 00 0 00 0 CHOCÓ 0 00 -3 - 30 -3000 0 00 1 -3 0 0 0 -1 0 0 -1 6 - - 4 00 0 - -3 0 0 0 -2 00 10 0 -1 0 00 00 -16 00 -1600 -1600 -16 00 -1600 -1600 -20 00 -10 00 -16 0 0 -1600 00 00 -4 Bajo Baudó 2 00 0 -1 6 0 0 0 COLOMBIA 00 0 00 -3 0 0 0 -3 0 0 00 00 -16 00 -1 6 0 0 00 -2 -20 -1 0 - 20 00 Nuquí 00 -3 0 0 0 -30 -4-4-4 00 0000 00 0 -30 00 -1600 00 -2 0 Bajo San Juan 00 -1 0 0 00 -3 -30 0 0 00 -3 0 00 - 00 00 0 00 - 1 60 0 -2000 -1600 0 00 -2 00 0 00 -2 -30 00 00 00 0 -20 Buenaventura 0 -1600 00 -1 -16 00 0 0 -3 0 00 0 -10 -300 00 López de Micay -2000 -4 0 Timbiquí Santa BárbaraGuapi 00 -300 0 0 -2000 -30 00 -1000 -3 00 -4 00 0 -30 -3000 00 0 0 - 1 000 -2 0 Banco de Tumaco Mosquera El Charco - 20 0 00 Francisco Pizarro -200 Banco Colombia 6 00 0 -1 -2 00 60 -3 -2000 -3000 0 -1 60 00 0 -2 00 OCÉANO PACÍFICO -1 00 -16 -16 -1 6 VALLE -3000 -10 00 -22 0 00 - 0 00 2 -20 - 00 20 -2000 00 0 -20 00 -1 0 -2000 0 0 0 00 00 2 -16 - 16 0 Malpelo 0 -20 0 - - 3 00 0 -2000 -3 0 -10 10 0 00 -1 0 -1600 -1600 00 6 00 -3000 -1 -2 -2 - Uribia 00 -5 0 -1 - -1 600 00 -160 0 -1000 -200 -200 -2 0 -160 0 0 0 Cove Bay Play a Bow ie's Punta South End 00 -4 00 - 0 - 20 -10 -10 San Andrés 0 -4 00 10 -3 0 00 -1 60 16 00 0 -2 0 -2 - 00 -1 0 -4 0 00 0 -10 - 40 -2000 -1 Punta Norte Bahía Honda 00 0 -5-5-5 -5 -40 ARCHIPIÉLAGO DE SAN ANDRÉS, PROVIDENCIA Y SANTA CATALINA 0 0 -4000 -5 -5 -5 -5 -5 -5-5 -5 -5 -5 -10 -5 -5 -5 -10 -5 -5-5 -5 -5 -5 -5 -5 -5 -5 -2000 -1000 --5 5 0 -1 -300 0 -4 00 0 0 0 -40 0 0 0 0 -4 0 0 00 -4 0 0 -4 0 00 -10 0 -1 -5-5-5 -5 -5 0 - 40 0 -4 0 - 40 0 - 4 00 -20 0 -1000 - 20 -5 0 -1 - 10 0 -5 -5 -5 -5 -5-5-5 -5 -5 -5-5 -5 - 1 0 00 00 00 -16 00 -4000 0 0 -4 0 0 -30 -5 - 2 0 00 -1 6 0 0 0 -5 0-5-5 -5 -5-5 -5 -10 -10 0 -5 -5 -5 -5 -1 -5-5 -5 -5 -5 -1 0 0 0 -1 0 -4 0 -1000 00 ISLA DE PROVIDENCIA Y SANTA CATALINA -10 -4 00 - -1000 -1 0 -1000 0 00 -2 -5 -1 0 -5 00 00 00 0 -20 -5-5 -5 0 -1 - 40 - 40 00 0 00 -1 -1 00 0 -5 -5 -5 -5 -5-5 00 00 -16 00 -16 00 -10 -5 -5 0 0 00 -4 0 -10 00 -4 -10 -100 -1 -1-1 00 -10-5-5-5-5 -5 -500 -1 -5 -10 -50 -1-50 -1 -5 - 1 -1 -1 -1-50 0 0-5-5 -5-5-5 -5-10 -10 -5-5-10 -10 -5 -10-10 0 -300 0 -10 -10-1 0 0 -5 - 10 0 -1 - 00 -10 -10 00 -10 -10 -10 -5-5-5 -10 -10 -10 0-5 -1-10 -10 -10 -5-5 -10 -5 -5 -10 0 -10 -10 -1-10 -10 -10 -1-10 -100 -1 -1 -1 000 -5 0 Banco Serrana - Serrana -10 Bank -10 -10 -5 -10 -10 -10 -10 -10 00 -2 -1 00 0 -1 -10 0 0 -10 0 -1-5 -10 -5 -10 -1 0 -1 Banco Quita Sueño - Quitasueño Bank 0 -3 00 -3 0 00 Tumaco NARIÑO CAUCA EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA -200 - -1 0 00 -10 -20 0 00 -1600 -2 0 00 -2000 -1000 -2 00 0 00 -16 - 20 - 20 0 0 00 -1600 -2000 -2 0 -1 6 0 0 00 -2000 -3 0 0 0 -2000 -1600 -2 00 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 0 -3 000 -2 -5 -10 1 00 40 -4 -4 0 00 -40 - -4 -5 -5 -5 -5 -5 -5-5 -5 -5 -5 -10 -5 -5 -5 -10 -5 -5-5 -5 -5 -5 -5 -5 -5 -4000 00 0 0 -2 - -300 0 0 -4 00 -5 -10 -10 -10 -5 0 00 -3 - 0 0 -3 0 0 4 -4000 00 -3000 - 3 000 0 00 -3 Bahía Solano CHOCÓ - 30 Nuquí 00 -3 0 0 0 -3000 - -3 0 0 0 -1 0 0 0 -1600 -1 6 -3 0 -1 6 0 0 10 5 00 - -3 0 0 0 0 -1 0 00 00 -1000 -1600 -1600 -1600 -1600 -1600 -2000 -16 0 0 -3 0 -1600 10 00 -30 0 0 -3 0 -2 0 Marlin blanco (Kajikia albida) 3. Dorado (Coryphaena hippurus) 4. Pez vela (Istiophorus albicans) 5. Marlin picudo (Tetrapturus pfluegeri) 6. Marlin azul (Makaira nigricans) -1600 -2000 0 00 00 -2 0 -2000 - -2000 00 -2 -2000 00 20 -2 VALLE -3000 -2000 -3000 Buenaventura -1600 0 -3000 6 -1600 0 -1 OCÉANO PACÍFICO 00 0 00 -10 López de Micay 00 -2000 -30 -2000 -4 0 Timbiquí Santa BárbaraGuapi 00 -3000 -1600 - 1 000 -200 Banco de Tumaco -3000 00 0 -4000 0 -16 0 CAUCA Mosquera El Charco 00 Francisco Pizarro -2000 6 0 -1 -1 6 -3 -2000 -3000 00 -1 6 00 -1600 00 0 -2 -1600 - 20 -3000 -3 0 2. Bajo San Juan 0 00 -2000 2 0 0 0 -2000 -1 0 Atún aleta amarilla (Thunnus albacares) 00 - 1 60 0 0 00 1. -2000 -3 0 00 -3000 -1600 -1600 0 00 -1 0 Malpelo -200 0 - Bajo Baudó -1600 00 00 -2 -2000 00 - 3 00 0 COLOMBIA 00 2 00 0 -1 6 0 0 - 4 00 0 -1600 -4 00 -3000 -1000 -3 0 0 0 -2000 - 20 00 -2000 0 -4000 -4000 -4000 - 00 -10 -10 -5 0-5-5 -5-5-5 -5-10 -10 -5-5-10 -10 -5 -10-10 -1000 -1 -2000 - ANTIOQUIA -4 -3000 0 -3 0 -3000 -10 -10 -10-10 -10 -200 -1 0 0 -1 00 -1000 -1 -1000 -1000 -2000 0 Apartadó Juradó -1000 00 - 30 SUCRE Turbo Unguía -1600 0 -3000 -5 -10 -10 -10 -10 --5 5 0 -1 - 2 0 00 -1 6 0 0 -1000 - 20 -5 0 -10 - 10 -5 -5-5-5 -5 -5 00 - 10 -2000 - - 16 0 0 -200 -5 -5 -5-5-5-10 -5-5-5-1 -10 -10 -10 -10 1-5 -5 -5-10 -5 -10 -10 -5 -5 -5 -50-10 -5 -5 -5 -5 -5 -5-5 -5 Puerto Escondido Los Córdobas 00 -3000 -3000 -3000 00 -4000 00 -3000 -3000 -3000 -3000 -3 00 0 -3000 -3000 CESÁR ArboletesCÓRDOBA Necoclí -1600 -3000 -3000 -3 0 0 00 -3 0 0 0 - -3000 -3000 MAGDALENA BOLÍVAR -1000 -3000 -3000 Cartagena de Indias Acandí -2000 00 00 -1 0 0 -200 -200 -1000 -1600 -3000 -3000 ATLÁNTICO -5 3 00 -3000 LA GUAJIRA -3000 40 00 Riohacha Dibulla -10 PuertoSitioPueblo Ciénaga NuevoViejo Colombia Santiago de Tolú San Antero Moñitos -3 0 -100 -50 0 -3 00 60 San Onofre -3000 - 40 -1 Piojó - - 3 0 00 -500 -200 Uribia -5 -1000 - 0 -200 -500 -1600 00 -100 -5 0 Manaure -1000 - -4 0 0 - 4 00 -20 -1600 -2000 1 60 0 Santa Marta D.T.C.H. -5 6 -3000 -4 0 1 -50 2 0 00 -3000 -3000 -10 -10 00 -4 -2000 -1600 00 0 MAR CARIBE -1 -1 0 - 30 -3000 -1 00 0 -16 0 -2 0 -3000 600 - 16 -3 -1000 -3000 - 3 00 - -4000 -1 -2000 00 -4000 -5-5 -5 -10 -5 -5 -5-5 -10 -10 -5--5 10 -3 00 0 -4 0 -40 -3000 - 3 00 0 -3000 00 -4000 00 0 00 0 -4000 - 40 4 000 -3 0 0 0 - 30 - 3 00 -3000 -3000 -3000 -3000 30 -200 -3000 -5 -5 -5 -5 -5-5-5 -5 -5 -5-5 -5 - 1 -3000 0 -1000 -3 0 0 0 -1000 00 0 -2 -4 0 0 0 - 40 0 0 -1 60 00 -1600 -3000 -3000 -30 0 -3 00 -3000 -4 00 00 16 00 00 0 - 40 0 -4 0 - 40 0 0 -1600 30 0 00 -4 0 00 ARCHIPIÉLAGO DE SAN ANDRÉS, PROVIDENCIA Y SANTA CATALINA -5 5 -5 -5--5-5 -5 -10 -1000 -3000 - Banco Colombia 6 00 Tumaco NARIÑO -2000 -1 -3 0 00 -200 10 0 0 - -1 0 - 10 00 00 -10 -20 0 00 -1600 - 1 0 00 -2 0 00 -2000 -1000 -2 00 0 00 -16 - 20 - 20 0 0 00 -1600 -2000 -2 0 -1 6 0 0 00 -2000 -3 0 0 0 -2000 -1600 -2 00 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 0 -3 000 -2 -1000 -5 -10 1 -1 00 40 -10 -10 -5 0-5-5 -5-5-5 -5-10 -10 -5-5-10 -10 -5 -10-10 -10 -10 -200 -5 -1 0 -4 0 -1 00 -40 - -1 -1000 00 -4 0 -1000 -5 -5 -5 -5 -5 -5-5 -5 -5 -5 -10 -5 -5 -5 -10 -5 -5-5 -5 -5 -5 -5 -5 -5 -4 -4000 -1000 00 0 0 -2000 -2 - -2000 - -300 0 -10 -10 -10-10 -10 -10 -10 --5 5 0 -1 - 2 0 00 -1 6 0 0 -1000 - 20 -5 0 -10 - 10 0 -4 00 - -5 -10 -10 0 00 -10 -5 -3 0 CHOCÓ 0 -3000 0 00 -3 - 30 -3 0 0 0 11. Nuquí 00 -1 6 - 00 -1 0 -2000 00 00 -2 0 00 0 2 0 00 -2 0 -2000 00 0 00 -2000 00 0 -2000 -1 0 -2000 0 0 00 Bajo San Juan 00 -2000 Malpelo -200 0 - -2000 -3 0 10 -30 0 0 -2 -2000 - 0 -1600 -1600 -3 0 00 -1000 -1600 -1600 -1600 00 00 -1 0 - 1 60 0 - 3 00 0 00 20 00 -16 0 0 -1600 0 -2000 -2000 -4 00 -3000 Bajo Baudó 10 -3 0 0 0 -2 -2 VALLE -3000 -1600 00 -2000 -1600 -1 -1600 Buenaventura 0 00 -3000 -10 00 -2000 -30 Timbiquí Santa BárbaraGuapi 00 - 1 000 -200 -3000 0 -3 0 López de Micay -4 0 Banco de Tumaco Banco Colombia 6 00 -2000 -1 -3 0 00 CAUCA Mosquera El Charco Francisco Pizarro Ejemplar de atún aleta amarilla Thunnus albacares capturado por la flota industrial con palangre industrial o longline que opera en el Caribe colombiano 00 13. -3000 Larva de atún aleta amarilla Thunnus albacares -2000 12. 0 00 -4000 00 -2000 -1600 -16 0 -3 -2000 OCÉANO PACÍFICO -1 6 -3000 -1600 6 0 0 00 -1600 00 0 -2 -1600 00 -1 6 -1 COLOMBIA Microfotografía (10 X) de ovario de atún aleta amarilla Thunnus albacares en estado de madurez V (Desovando). 2 00 0 -1 6 0 0 - 4 00 0 00 00 -3000 -1600 0 -4000 -4000 -4000 -3 0 0 0 -3 0 -2000 - 20 00 -2000 -1000 -1 6 0 0 -3 0 0 -1600 -1 0 0 -1600 -1600 -3 0 0 0 -1600 -1600 - 0 00 Bahía Solano -4000 - 3 000 - 00 0 00 -3000 0 -3 0 -3000 00 - 30 00 0 -3000 -3 00 0 -3000 -5 -5 -5-5-5 -5 00 - 10 -2000 - -3 0 -1000 -4000 -3000 -3000 0 ANTIOQUIA -4 -3000 00 -3000 -3000 -3000 00 -3 -3000 -3000 -3000 - 16 0 10. Muestreo oceanográfico durante cruceros de prospección pesquera -1000 -3000 Apartadó Juradó -3000 -3000 00 -3000 -3000 -3 0 0 00 0 0 0 - -3000 -3000 40 00 00 Operaciones de pesca con palangre industrial o longline 00 - 40 0 -1600 9. -3 -3000 -3000 -200 -5 -5 -5-5-5-10 -5-5-5-1 -10 -10 -10 -10 1-5 -5 -5-10 -5 -10 -10 -5 -5 -5 -50-10 -5 -5 -5 -5 -5 -5-5 -5 Unguía -3 - SUCRE Turbo -3000 -3000 -1 0 0 -200 -200 -1000 Puerto Escondido Los Córdobas ArboletesCÓRDOBA Necoclí -3000 0 -1 -3000 CESÁR Acandí - 20 - 0 -5 MAGDALENA BOLÍVAR Santiago de Tolú San Antero Moñitos -3000 -3000 -4 0 0 - 4 00 Cartagena de Indias 7. y 8.Embarcaciones de pesca industrial de recursos grandes pelágicos - 0 -4 0 Riohacha LA GUAJIRA San Onofre -1000 - 3 0 00 -10 -10 00 -4 -2000 -1600 Ciénaga ATLÁNTICO -1000 - - 30 -3000 -1 -2 -3000 600 - 16 -3000 -4000 -1 -2000 00 Dibulla -10 PuertoSitioPueblo NuevoViejo Colombia -5 6 13 -2 Manaure Santa Marta D.T.C.H. -5 Piojó -1 00 -50 0 -2 -3 0 00 -3000 -3 0 -100 60 00 -3000 -3000 -3 -200 - 30 -3000 -3 0 0 0 00 -500 -500 -200 Uribia -100 -5 0 -3 -3 0 -3000 -200 0 00 00 -50 1 60 0 -3000 - 3 00 -3000 30 -20 -1600 -2000 0 00 -3000 - 3 00 0 -3000 00 -5 -5 -5 -5-5 -5 -5 -5 -5 -5 -5-5 -5 - 1 0 -3000 - 30 -1000 - 3 00 -3000 -3 0 00 -3 00 0 0 -10 -10 0 -3000 -10 00 0 -3000 -10 -10 -5 -10 -10 -10 -5-5 -10 -5 -5 -10 -3000 -5 -10 -10 -5 -3 -3000 -3000 -3000 -30 0 00 -1600 0 MAR CARIBE -1000 00 00 00 0 -16 0 -1600 - -3000 -3000 -3000 00 0 30 - 30 -40 -3 -1000 - -3000 -3 -3000 40 0 0 00 -4000 -4000 -5-5 -5 -10 -5 -5 -5-5 -10 -10 -5--5 10 00 -3 0 -3000 3 000 -3000 00 -3000 00 -30 -3 0 00 0 0 -4 0 -4000 -1600 0 30 -3000 -3 0 - 3 00 -4 00 0 0 00 4 000 -10 -1000 - 30 0 00 -3000 - -3 0 00 -30 -4 0 0 - 40 -5 5 -5 -5--5-5 -5 -10 -1000 00 -3000 -30 0 0 -4 0 00 0 -40 0 -1 -1 6 00 -3 0 -3000 -3000 - 3 00 0 -3 0 -3000 -3000 -3000 -3000 0 -3000 -3000 -3 0 0 0 -2 0 0 - 20 -10 -10 -3000 -3000 - -3 0 0 -3 0 00 0 00 0 00 00 00 0 -3000 10 -4000 0 - 40 0 -4 0 - 40 0 - 40 0 0 -1 60 -1000 -3 0 -1600 -2000 Punta Norte Bahía Honda 0 0 00 00 16 Cove Bay Play a Bow ie's Punta South End -10 -5 0-5-5 -5 -5-5 -10 -10 -5 -5-5 -10 -10 -5 -5 -5 -5-5 --10 -5 5 -10 -10 - 5-5-5-5 0 00 0 0 -4 0 0 ARCHIPIÉLAGO DE SAN ANDRÉS, PROVIDENCIA Y SANTA CATALINA -2000 0 -3 -3 0 0 0 -5 0-5-5 -5 -5-5 -5 -10 -10 -5 -5 -5 -5 -10 -5-5 -5 -5 -5 -1600 San Andrés -4 00 4 00 0 00 -1000 0 00 -2 0 -1 0 -1000 - 40 - 40 00 00 -1 0 0 0 -1 00 0 -3 00 0 000 00 0 -1000 -3000 00 -3 0 - 30 0 -1 0 -1000 - 16 0 -3000 0 00 -3 0 00 -1 -2000 00 -3000 0 00 -3 -3000 - 30 0 0 00 0 -5-5 -5 -10 -5 -5 -4000 0 -3000 00 -5 -10 -5 00 - 40 00 - 4 00 0 00 - 2 00 0 -1 0 -30 -3000-3000 -3 -3000 0 00 -3000 0 00 -2 0 0 0 -2 0 0 0 -5 -5 -5 -5 -5-5 -5-5-5 -5 -3000 -3000 -3 -3 0 00 -3 0 30 -3000 6 00 0 00 ISLA DE PROVIDENCIA Y SANTA CATALINA 00 30 -3000 -5 -1600 00 -3000 0 -10 00 -10 -10 - 30 0 -3 0 0 -10-10 -10-5-5-5-5 -5 -5 -10 -5 -5 -10 -5-1-10 0 - 1 -10 -5 0 -3 0 - 30 0 0 - 0 -5 - 10 0 -1000 -1000 0 0 -10 -3 - 300 0 00 -3000 0 00 - -2 0 0 0 -10 -10 -10 -5-5-5 -10 -10 -1600 -3000 -3000 0 -50 00 - 30 -10 10 12 -10 -10 -10 -10 -10 -10 -100 11 -10 -10 -10 -10 -10 00 Banco Serrana - Serrana -10 Bank -10 -10 -5 -10 0-10 -10 1-10 -10 --10 -10 -10-10 10 -10 Banco Quita Sueño - Quitasueño Bank -1 00 0 -10 -10 -10 9 -10 -10 -5-5 -5 -5 -10 -10 -5 -10 -5 -10 -5 -5-5 -5 -5-10 -5 -5-5-5 -5-10-5-5 -10 -10 -5-10 -10 8 00 -10 -10 -10 -10 -10 7 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA -3 -3000 -3000 00 00 0 0 -3 -3000 -3000 -3 0 -3000 00 -3000 - 40 0 0 00 0 0 -4 0 0 -3000 -10 Cove Bay Play a Bow ie's Punta South End -10 -5 0-5-5 -5 -5-5 -10 -10 -5 -5-5 -10 -10 -5 -5 -5 -5-5 --10 -5 5 -10 -10 - 5-5-5-5 0 00 -4000 -2 0 0 - 20 -10 -10 San Andrés - 40 - 40 00 0 00 -30 -3000 3 000 -3000 00 - 30 0 -5 0-5-5 -5 -5-5 -5 -10 -10 -5 -5 -5 -5 -10 -5-5 -5 -5 -5 -1600 0 -4 00 4 00 0 0 -40 0 0 -1 0 -1000 -3000 -30 0 0 -3000 -3000 -3000 -3000 30 000 00 00 -1 0 0 0 10 -4000 -3000 0 -5 -10 -5 00 -2000 00 -3000 -2 -10 -10 -5 -5-5 -1000 -1000 -1 6 00 -3 0 -3000 - 30 0 0 - -3 0 0 -3000 00 -1 00 -1000 0 00 -2 -2000 -3 0 00 -1 0 ISLA DE PROVIDENCIA Y SANTA CATALINA Punta Norte Bahía Honda - 40 00 0 00 -2000 -5-5 -5 -10 -5 -5 -1 -3000 -1 0 00 0 -1000 -3000 -3000 0 -1000 -5 -5 -5 -5-5-5 -5 -3000 -3 0 30 -3000 00 - 2 00 0 -10 -10 00 0 -3000 -2 0 0 0 -5 -3 0 00 -2 0 0 0 -1000 0 -3 -3 0 0 -10 -10 00 0 -3 00 0 00 - 4 00 0 -100 00 -3 0 - 30 0 -10 -10 -5 -10 -10 -10 -5-5 -10 -5 -5 -10 -10 0-10 -10 1-10 -10 --10 -10 -10-10 -3000 0 00 6 -1600 - 16 0 -3000 -3 -10-10 -10-5-5-5-5 -5 -5 -10 -5 -5 -10 -5 -10 0 -10 -5 - 1 -1 0 00 00 -3000 0 00 -3000 0 00 0 -30 -3000-3000 -3 -3000 -3 -3 0 00 -5 -10 -10 -5 -10 -3000 -3000 0 -3 0 0 0 -5 - 10 0 -1600 00 30 -30 0 0 - 00 00 -3000 -10 -10 -10 -5-5-5 -10 -10 - - 30 0 -10 -50 00 0 -3 0 -10 -10 -10 -10 -10 -10 00 Banco Serrana - Serrana -10 Bank -10 -10 -5 -10 -10 -10 -10 -10 -10 10 -3 - 300 0 00 -3000 -1 00 0 -10 -10 -10 -10 -10 -10 -10 -3000 -3000 0 00 - -2 0 0 0 -1600 -10 -10 -10 Banco Quita Sueño - Quitasueño Bank -5-5 -5 -5 -10 -10 -5 -10 -5 -10 -5 -5-5 -5 -5-10 -5 -5-5-5 -5-10-5-5 -10 -10 -5-10 -10 0 0 00 -1000 - 30 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA -3 3000 30 10 0 0 - 10 00 - 1 0 00 Tumaco NARIÑO Francisco Armando Arias Isaza Director General Otto Polanco Rengifo Director General Jesús Antonio Garay Tinoco Subdirector de Coordinación Científica José Duarte Carreño Secretario General Sandra Rincón Cabal Subdirectora Administrativa Sergio Manuel Gómez Flórez Jefe Oficina de Generación del Conocimiento y la Información Paula Cristina Sierra Correa Coordinadora de Investigación e Información para la Gestión Marina y Costera David A. Alonso Carvajal Coordinador Programa Biodiversidad y Ecosistemas Marinos Mario Enrique Rueda Hernández Coordinador Programa de Valoración y Aprovechamiento de Recursos Marinos Luisa Fernanda Espinosa Díaz Coordinadora Programa de Calidad Ambiental Marina Constanza Ricaurte Villota Coordinadora Programa de Geociencias Marinas Erick Serge Firtion Esquiaqui Director Técnico de Administración y Fomento Lázaro de Jesús Salcedo Caballero Director Técnico de Inspección y Vigilancia Neil Gallardo García Director Regional Barranquilla Jaime Albornoz Rivas Director Regional Cali Carlos Augusto Borda Rodríguez Supervisión Técnica y Administrativa Julián Mauricio Betancourt Portela Coordinador de Servicios Científicos Calle 40A # 13 - 09, Edificio Ugi. Pisos 6, 14 y 15 Bogotá D.C., Colombia Tel: (571) 377 5500 www.aunap.gov.co Calle 25 # 2 - 55 - Playa Salguero Rodadero, Santa Marta (Colombia) PBX: (575) 432 8600 www.invemar.org.co Convenio # 148 de 2015 Citar la obra completa así: Rodríguez, A., M. Rueda y F. Escobar. (Eds). 2015. Evaluación directa de las poblaciones de peces grandes pelágicos del Pacífico y Caribe continental de Colombia. INVEMAR y AUNAP. Serie de Publicaciones Generales de INVEMAR Nº 87. Serie Recursos Pesqueros de Colombia - AUNAP 2015. Santa Marta, Colombia. 120p. Citar los capítulos así: Autores. 2015. Título del capítulo. Pp. (intervalo de páginas), en Rodríguez, A., M. Rueda y F. Escobar. (Ed). 2015. Evaluación directa de las poblaciones de peces grandes pelágicos del Pacífico y Caribe continental de Colombia. INVEMAR y AUNAP. Serie de Publicaciones Generales de INVEMAR Nº 87. Serie Recursos Pesqueros de Colombia - AUNAP 2015. Santa Marta, Colombia. 120p. Palabras clave: grandes pelágicos, prospección pesquera, procesos poblacionales, evaluación y manejo. ISBN de la obra impresa: 978-958-8935-11-9 ISBN de la obra digital: 978-958-8935-12-6 © Derechos reservados según la ley, los textos pueden ser reproducidos total o parcialmente citando la fuente Fotografías de portada, contraportada y páginas internas: Archivo fotográfico Programa VAR - INVEMAR Cartografía: Felipe Valencia. LABSIS - INVEMAR Diseño: Carlos González/John Khatib (ediprint.com.co) Impresión: Ediprint S.A.S. 500 ejemplares Diciembre de 2015 Bogotá D.C., Colombia • Las líneas de delimitación fronteriza presentadas en este documento son una representación gráfica aproximada, con fines ilustrativos solamente. 4 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA ABREVIATURAS Y SIGLAS ANM Altura del Nivel del Mar AUNAP Autoridad Nacional de Acuicultura y Pesca CCPC Contracorriente Panamá-Colombia CGSM Ciénaga Grande de Santa Marta CIATComisión Interamericana del Atún Tropical CLAClorofila-a GPTGeneral Purpose Transceiver (Transceptor de Propósito General) IDV Integrated Data View (Visor de Datos Integrados) LABSIS Laboratorio de Sistemas de Información LH Longitud Horquilla LT Longitud Total CO Captura Objetivo NASC Nautical Area Scattering Coeficient (Coeficiente de Dispersión del Área Náutica) CPUA Captura por Unidad de Área ODVOcean Data View (Visor de Datos Oceánicos) CPUE Captura por Unidad de Esfuerzo SIG Sistema de Información Geográfica EEPEnfoque Ecosistémico de la Pesca TMCTalla Media de Captura FA Fauna Acompañante TMM Talla Media de Madurez FAOOrganización de las Naciones Unidas para la Alimentación y la Agricultura TS Target Strength (Fuerza de Blanco) GAMGeneralized Additive Models (Modelos Aditivos Generalizados) GCPGiro Ciclónico de Panamá GCVGeneralized Cross Validation Score (Criterio de Validación Cruzada Generalizado) TSMTemperatura Superficial del Mar UBMUnidades Básicas de Muestreo ZCITZona de Convergencia Intertropical EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 5 UNIDADES DE MEDIDAS USADAS % Porcentaje m Metro °C Grados centígrados m2 Metro cuadrado µg Microgramo m3 Metro cúbico cm Centímetro mg Miligramo h Hora ml Mililitro kg Kilogramo mm Milímetro kHzKilohertz mn Milla náutica km Kilómetro mn2 Milla náutica cuadrada km2 Kilómetro cuadrado s Segundo l Litro t Tonelada 6 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA AGRADECIMIENTOS Los autores desean agradecer a las siguientes instituciones, empresas y personas que contribuyeron a la publicación de este libro: A la Autoridad Nacional de Acuicultura y Pesca –AUNAP– por su gestión en ejecución de la política pesquera y retomar las prospecciones pesqueras como fuente de información para la evaluación y manejo de pesquerías en el país. Inicialmente a la actual directiva de a AUNAP en cabeza del Dr. Otto Polanco Rengifo, Director General, y apoyada por el actual jefe de la OGCI, Dr. Sergio Manuel Gómez Flórez. Así mismo, agradecer todo el apoyo de los doctores Julián Botero Arango, Vladimir Puentes Granada y José Ricardo Fonseca durante el tiempo que permanecieron en la entidad. En este mismo sentido, los autores agradecen todo el apoyo brindado por los diferentes profesionales de la AUNAP, Carlos Augusto Borda, Jhon Gabriel Ramírez, Carlos Polo Silva y Wilberto Angulo Viveros. Adicionalmente se agradece a las Direcciones Regionales del Caribe y Pacífico Dr. Neil Gallardo García y Dra. Juana de Dios Murillo (directora encargada durante 2014), respectivamente. Al CN (R) Francisco A. Arias Isaza director general del INVEMAR y los subdirectores Dr. Jesús A. Garay Tinoco y Sandra Rincón Cabal y demás dependencias del INVEMAR, por su valioso soporte administrativo y logístico. Al equipo administrativo y técnico del Programa de Valoración y Aprovechamiento de Recursos Marinos (VAR) y la Sede Pacífico del INVEMAR (Stephannie Chávez, Germán Angulo, Jorge Sierra, Jessica Eraso, Víctor Webber, Carlos Puentes, Marlon Montero, Hernando Hurtado y José Sánchez), por su invaluable labor en el desarrollo de esta investigación. Al Laboratorio de Sistemas de Información del INVEMAR, por su labor en la producción cartográfica del libro. A las empresas de la flota industrial de grandes pelágicos que operan con palangre (longline) en el Caribe y Pacífico colombiano, Imanely SAS y Sepúlveda Rodgers & Cia. Ltda, respectivamente. Estas empresas brindaron su apoyo y servicios a la investigación mediante el uso de las embarcaciones pesqueras Haleluya (en Caribe) e Hifer (en Pacífico), como plataformas de investigación. Sus tripulaciones (capitanes y pescadores) compartieron su conocimiento tradicional de la pesca de grandes pelágicos para las faenas de pesca exploratoria realizadas durante los cruceros de prospección. A las capitanías de los puertos de Cartagena y Buenaventura por facilitar la gestión en la realización de los cruceros de prospección pesquera. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 7 PRÓLOGO Como parte de sus funciones misionales, la Autoridad Nacional de Acuicultura y Pesca –AUNAP– realiza esfuerzos con socios estratégicos a través de convenios de cooperación técnica, cuya finalidad es la de investigar el potencial y estado de los recursos pesqueros del país. En este caso la AUNAP suscribió el convenio No. 148-2015 con el Instituto de Investigaciones Marinas y Costeras – INVEMAR, centro de investigación de amplio reconocimiento nacional e internacional que ha sido apoyo de la institucionalidad pesquera por más de quince años en los temas de evaluación de recursos marinos de interés para la pesca. Con los resultados de este convenio, la AUNAP propende por fundamentar sus decisiones de manejo con base en la mejor evidencia científica disponible. En consecuencia, un producto del convenio antes mencionado es este libro titulado “EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA”, el cual busca actualizar la poca información existente que soporte un conocimiento de los recursos de peces grandes pelágicos, recurso clave de la pesca de nuestro país que puede ser desarrollado de manera responsable a la luz de los resultados obtenidos en esta investigación conjunta. Este libro extrae los principales tópicos derivados de los convenios que se han adelantado en investigaciones sobre este tipo de recursos (Convenio No. 038-2013 y Convenio No. 140-2014), los cuales se basaron en prospecciones pesqueras directas, usando como plataformas de investigación a barcos de la flota industrial de palangre en el Caribe continental y Pacífico. Esta estrategia permitió combinar tecnología de investigación pesquera y ambiental de punta con el conocimiento tradicional del pescador, lo cual confiere mayor robustez a los resultados obtenidos y a las medidas de manejo que a partir de ellos puedan formularse. El libro sintetiza el trabajo de prospección pesquera en ambas costas del país, abordando los diferentes procesos poblacionales clave de los recursos pelágicos, así como el efecto de variables ambientales y las implicaciones de manejo con fines de un aprovechamiento sostenible. OTTO POLANCO RENGIFO Director General Autoridad Nacional de Acuicultura y Pesca -AUNAP- 8 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 3 5 Tabla 3-1 Muestreo hidroacústico durante los cruceros de investigación de grandes pelágicos en el Caribe y Pacífico colombiano. [25] Tabla 3-6 Muestreo biológicopesquero durante los cruceros de investigación de grandes pelágicos en el Caribe y Pacífico colombiano. [28] Tabla 5-1 Función de fuerza de blanco o Target Streght (TS) del atún aleta amarilla (Thunnus albacares) y dorado (Coryphaena hippurus). [44] Tabla 3-2 Coordenadas geográficas de las estaciones de pesca exploratoria (EP) con palangre superficial horizontal durante el crucero de investigación en peces grandes pelágicos en el Caribe colombiano. [26] Tabla 3-7 Escala de madurez sexual para hembras de túnidos propuesta por Figueiredo et al. (2008). [28] Tabla 5-2 Resumen del análisis geostadístico para la estimación de biomasa del atún aleta amarilla (Thunnus albacares) y el dorado (Coryphaena hippurus) en el Caribe continental colombiano. Co: nugget o efecto pepita; C+Co: Sill o varianza; Ao: Rango o distancia de autocorrelación (km); CME: Cuadrado medio del error. [45] Tabla 3-3 Coordenadas geográficas de las estaciones de pesca exploratoria (EP) con palangre horizontal de superficie durante el crucero de investigación de peces grandes pelágicos en el Pacífico colombiano. [26] Tabla 3-4 Características técnicas del palangre o longline empleado durante el crucero de grandes pelágicos en el Caribe colombiano. [27] Tabla 3-5 Características técnicas del palangre o longline empleado durante el crucero de grandes pelágicos en el Pacífico colombiano. [27] Tabla 3-8 Escala de madurez sexual general para desovadores parciales propuesta por Holden y Raitt, (1975). [29] Tabla 3-9 Registro fotográfico de las gónadas durante los cruceros de investigación de grandes pelágicos en el Caribe y Pacífico colombiano. [29] Tabla 3-10 Muestreo oceanográfico durante los cruceros de investigación de grandes pelágicos en el Caribe y Pacífico colombiano. [30] 4 Tabla 4-1 Número de anzuelos y tiempo de reposo empleado durante los cruceros de investigación en peces grandes pelágicos en el Caribe y Pacífico colombiano. [34] Tabla 5-3 Resumen del análisis geostadístico de la densidad del dorado (Coryphaena hippurus) en el Pacífico colombiano. Co: nugget o efecto pepita; C+Co: Sill o varianza; Ao: Rango o distancia de autocorrelación (km); CME: Cuadrado medio del error. [48] 6 Tabla 6-1 Tiempos para la tinción con la técnica de hematoxilinaeosina (Gómez-León, 2005). [54] Tabla 6-2 Sexo y estados de madurez macroscópicos de peces capturados en los cruceros de investigación de grandes pelágicos realizados durante el 2013 y 2014 en el Caribe colombiano. [55] Tabla 6-3 Porcentaje de diferencia relacionando el estado definido a nivel macroscópico y el determinado en el análisis histológico para Thunnus albacares (atún aleta amarilla). N: número de individuos que presentan diferencias en la clasificación, %D: porcentaje que representa cada diferencia. HI: hembra uno, HII: hembra dos, HIII: hembra tres, HIV: hembra cuatro, HV: hembra cinco, H?: hembra sin identificar estado de madurez, MI: macho uno, MII: macho dos, MIII: macho tres, MIV: macho cuatro, MIV: macho cinco. [57] Tabla 6-4 Escala de madurez gonadal macroscópica y microscópica propuesta para Thunnus albacares (atún aleta amarilla). Se incluye la descripción macro y microestructural de los estados de madurez gonadal. [58] EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 9 ÍNDICE DE TABLAS Tabla 6-5 Sexo y estados de madurez macroscópica de peces capturados en la campaña de grandes pelágicos en el Pacífico colombiano. [63] Tabla 6-6 Porcentaje de diferencia relacionando el estado definido a nivel macroscópico y el estado determinado en el análisis histológico. N: número de individuos que presentan diferencias en la clasificación, %D: porcentaje que representa cada diferencia. HII: hembra dos, HIII: hembra tres, HIV: hembra cuatro, HV: hembra cinco, M?: macho sin identificar estado de madurez, MII: macho dos, MIII: macho tres, MIV: macho cuatro, MIV: macho cinco. [65] Tabla 6-7 Escala de madurez gonadal macroscópica y microscópica propuesta para el dorado Coryphaena hippurus, se incluye la descripción macro y microestructural de los estados de madurez gonadal. [66] 7 Tabla 7-1 Valores de densidad (N/10 m2) de huevos y larvas de peces durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. [74] Tabla 7-2 Composición, porcentaje de abundancia (%A) y frecuencia de ocurrencia en porcentaje (%FO) de larvas de peces encontradas durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. NF: identificados a nivel de familia. [76] Tabla 7-3 Valores de abundancia (N/10 m2) de huevos y larvas de peces durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. [84] Tabla 7-4 Composición, porcentaje de abundancia (%A) y frecuencia de ocurrencia en porcentaje (%FO) de larvas de peces encontradas durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. NF: identificados a nivel de familia. [86] 8 Tabla 8-1 Modelación GAM usada para la relación entre la captura por unidad de esfuerzo -CPUE (Número de individuos/h*100 anzuelos) y los predictores [temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m) y longitud (°)], en el Caribe colombiano. Dev.Exp: Devianza explicada (%). GCV: criterio de validación cruzada generalizado. [101] Tabla 8-2 Modelación GAM usada para la relación entre la captura por unidad de área -CPUA (Número de individuos /m2) del ictioplancton y los predictores [temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m) y longitud (°)], en el Caribe colombiano. Dev.Exp: Devianza explicada (%). GCV: criterio de validación cruzada generalizado. [102] Tabla 8-3 Modelación GAM usada para la relación entre la captura por unidad de esfuerzo -CPUE (Número de individuos/h*100 anzuelos) de grandes pelágicos y los predictores: temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m), latitud (°) y longitud (°), en el Pacífico colombiano. Dev. Exp: Devianza explicada (%). GCV: criterio de validación cruzada generalizado. [106] Tabla 8-4 Modelación GAM usada para la relación entre la captura por unidad de área -CPUA (Número de individuos /m2) del ictioplancton y los predictores: temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m), latitud (°) y longitud (°), en el Pacífico colombiano. Dev. Exp: Devianza explicada (%). GCV: criterio de validación cruzada generalizado. [106] 9 Tabla 9-1 Indicadores y medidas de manejo y conservación para recursos grandes pelágicos en Colombia. [111] 10 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 3 Figura 3-1 Mapa del área de estudio, ubicando las estaciones de muestreo divididas en tres zonas (norte, centro y sur) durante el crucero de investigación en peces grandes pelágicos entre mayo y junio de 2014 en el Caribe colombiano. [23] Figura 4-3 Comparación de la captura total en a) número de individuos y b) peso por zona de muestreo de grandes pelágicos en el Caribe colombiano. [36] Figura 4-4 Comportamiento de la captura (kg) y la captura por unidad de esfuerzo (CPUE: N/100 anzuelos*h) en cada estación (EP) y zona de muestreo en el Caribe colombiano. [36] Figura 3-2 Mapa del área de estudio, ubicando las estaciones de muestreo divididas en tres zonas (costera, intermedia y oceánica) Figura 4-5 Comparación de la durante el crucero de investigación captura por unidad de esfuerzo en peces grandes pelágicos entre -CPUA (Número de individuos/100 noviembre y diciembre de 2014 en el Pacífico colombiano. [24] anzuelos*h) por zona de muestreo de grandes pelágicos en el Caribe colombiano. [37] 4 Figura 4-1 Composición porcentual de la captura en peso a nivel de familias durante el crucero de investigación de peces grandes pelágicos en el Caribe colombiano. [34] Figura 4-2 Composición de la captura en número de individuos (N) y peso (kg) para las zonas de muestreo: a) norte, b) centro y c) sur, en el Caribe colombiano. [35] Figura 4-8 Composición de la captura en peso (kg) y número de individuos (N) para las zonas de muestreo: a) costera, b) intermedia y c) oceánica, en el Pacífico colombiano. [39] Figura 4-9 Comparación de la captura total en peso (kg) y b) Número de individuos, por zona de muestreo de peces grandes pelágicos en el Pacífico colombiano. [38] Figura 4-10 Comportamiento de la captura (kg) y la captura por unidad de esfuerzo (CPUE: N/100 anzuelos*h) en cada estación (EP) y zona de muestreo en el Pacífico colombiano. [40] Figura 4-6 Análisis de clasificación de las especies calculado con Figura 4-11 Comparación de la el coeficiente de similitud de captura por unidad de esfuerzo Morisita-Horn, utilizando la -CPUE (N/100 anzuelos*h) por zona abundancia a partir de la CPUE de muestreo de grandes pelágicos (N/100 anzuelos*h) para los en el Pacífico colombiano. [40] grandes pelágicos en el Caribe colombiano. [37] Figura 4-12 Análisis de clasificación de las especies Figura 4-7 Composición porcentual calculado con el coeficiente de de la captura en peso a nivel similitud de Morisita-Horn utlizando de familias durante el crucero la abundancia a partir de la CPUE de investigación de peces (N/100 anzuelos*h) para los grandes pelágicos en el Pacífico grandes pelágicos en el Pacífico colombiano. [38] colombiano. [40] 5 Figura 5-1 a) Montaje del transductor en la parte lateral del barco para la detección instantánea de peces. b) Diagrama del sistema de información hidroacústica para visualización de las ecodetecciones (ecograma). [43] Figura 5-2 Esquema de calibración de la ecosonda mediante una esfera con fuerza de blanco conocida y suspendida bajo el eje del haz acústico del transductor. [44] Figura 5-3 Representación gráfica de la función de variograma. [45] Figura 5-4 Variograma omnidireccional de la densidad de a) atún aleta amarilla (Thunnus albacares) y b) dorado (Coryphaena hippurus), en el Caribe continental colombiano. [45] Figura 5-5 Distribución espacial de la densidad (t/mn2) del atún aleta amarilla (Thunnus albacares) en el Caribe continental colombiano. [46] Figura 5-6 Distribución espacial de la densidad (t/mn2) del dorado (Coryphaena hippurus) en el Caribe continental colombiano. [46] EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 11 ÍNDICE DE FIGURAS Figura 5-7 Distribución espacial de las tallas (cm de longitud total) del atún aleta amarilla (Thunnus albacares) en el Caribe continental colombiano. [47] Figura 5-8 Distribución espacial de las tallas (cm de longitud total) del dorado (Coryphaena hippurus) en el Caribe continental colombiano. [47] Figura 5-9 Variograma omnidireccional de la densidad del dorado (Coryphaena hippurus) en el Pacífico colombiano. [48] Figura 5-10 Distribución espacial de la densidad (t/mn2) del dorado (Coryphaena hippurus) en el Pacífico colombiano. [48] Figura 5-11 Distribución espacial de las tallas (cm de longitud total) del dorado (Coryphaena hippurus) en el Pacífico colombiano. [48] 6 Figura 6-1 Proceso metodológico para la obtención de cortes histológicos de una fracción de gónada. a) ubicación de las gónadas en bandeja plástica para su posterior corte, b) herramientas de trabajo para el procedimiento (bisturí, pinza y rejilla), c) corte longitudinal de la parte media de la gónada seleccionada para el análisis histológico, y d) gónada recolectada de gran tamaño. [52] Figura 6-2 Proceso metodológico para la obtención de cortes histológicos de una fracción de gónada. a) realización de cubos de parafina con la gónada inmersa, b) organización de los cubos para su posterior corte, c) micrótomo utilizado para los cortes, y d) corte de los cubos de parafina mediante el micrótomo para la obtención de las placas histológicas. [53] Figura 6-3 Ejemplos de placas histológicas obtenidas, para su posterior análisis descriptivo y comparativo. [54] Figura 6-4 Histograma de frecuencia de tallas de los ejemplares de Thunnus albacares (atún aleta amarilla) capturados durante los cruceros de grandes pelágicos en el Caribe colombiano (2013 y 2014). La TMM indicada fue estimada por Cayré et al. (1993). [56] Figura 6-5 Distribución espacial de los individuos maduros del atún aleta amarilla (T. albacares) en el Caribe colombiano durante la campaña realizada entre mayojunio de 2014. [56] Figura 6-6 Distribución espacial de los individuos inmaduros del atún aleta amarilla (T. albacares) en el Caribe c olombiano durante la campaña realizada entre mayojunio de 2014. [57] Figura 6-7 Histograma de frecuencia de tallas de los ejemplares de dorado Coryphaena hippurus capturados durante la campaña de grandes pelágicos en el Pacífico colombiano. La TMM indicada fue estimada por SolanoSare et al. (2008) para el Pacífico peruano. [64] Figura 6-8 Distribución espacial de los individuos maduros de dorado (Coryphaena hippurus) en el Pacífico colombiano durante la campaña realizada en 2014. [64] Figura 6-9 Distribución espacial de los individuos inmaduros de dorado (Coryphaena hippurus) en el Pacífico colombiano durante la campaña realizada en 2014. [65] 7 Figura 7-1 Morfotipos de huevos encontrados durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. a) Morfotipo I; b) Morfotipo II; c) Morfotipo III y d) En estado de división. [75] Figura 7-2 Distribución espacial de la densidad (N/ 10 m2) de a) huevos y b) larvas, durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. [75] Figura 7-3 Porcentaje de abundancia de las familias de larvas de peces durante el crucero de investigación de grandes pelágicos en el Caribe continental colombiano. [76] Figura 7-4 Larvas de peces encontrados durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. a) Sardina (Clupeidae); b) Pez Linterna (Diaphus sp.); c) Merlucita (Bregmaceros atlanticus); d) Melamphaes sp; e) Cherna (Serranus spp.); f) Jurel (Caranx spp.); g) Gobiidae; h) Pez cirujano (Achanthurus sp.); i) Escolar (Gemphylidae); j) Lenguado (Bothus ocellatus); k) Globo (Sphoeroides spp); l) Balistes sp. [78] 12 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Figura 7-5 Distribución espacial de la abundancia de larvas (larvas/10 m2) de peces pelágicos de importancia comercial durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. [80] Figura 7-6 Morfotipos de huevos encontrados durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. a) y b) Morfotipo III. [84] Figura 7-7 Distribución espacial de la densidad (N/ 10 m2) de a) huevos y b) larvas durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. [85] Figura 7-8 Porcentaje de abundancia de las familias de larvas de peces durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. [86] Figura 7-9 Diversidad de larvas de peces encontrada durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. a) Anchoa (Cetengraulis mysticetus); b) Diaphus pacificus; c) Diogenichthys. Laternatus; d) Merlucita (Bregmaceros sp.); e) Merluza (Brotula sp.); f) Lisa (Mugil cephalus); g) Naucrates ductor ; h) Thalassoma sp.; i) Euthynus lineatum; j) Cubiceps pauciradiatus; k) Syacium sp.; l) Symphurus sp. [88] Figura 7-10 Distribución espacial de la abundancia de larvas (larvas/10 m2) del dorado (Coryphaena hippurus) durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. [90] 8 Figura 8-1 Distribución superficial de la temperatura (°C) durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. [94] Figura 8-2 Distribución superficial del oxígeno disuelto (%) durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. [95] Figura 8-3 Distribución superficial de la salinidad durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. [95] Figura 8-4 Distribución superficial del pH durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. [96] Figura 8-5 Distribución superficial de la clorofila-a (µg/l) durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. [96] Figura 8-6 Comportamiento a profundidad de a) temperatura (°C), b) salinidad, c) oxígeno disuelto (%), d) pH y e) clorofila-a (µg/l) durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. [97] Figura 8-7 Comportamiento a profundidad de la temperatura (°C) de a) salinidad, b) oxígeno disuelto (%) y c) clorofila-a (µg/l) durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. [98] Figura 8-8 Comportamiento promedio de la altura del nivel del mar (m) y corrientes superficiales (m/s) entre el 22 de mayo y el 12 de junio de 2014 en el Caribe colombiano. CGSM: Ciénaga Grande de Santa Marta. [99] Figura 8-9 Comportamiento promedio de la temperatura superficial del mar (TSM; °C) y corrientes (m/s) entre el 22 de mayo y el 12 de junio de 2014 en el Caribe colombiano. CGSM: Ciénaga Grande de Santa Marta. [99] Figura 8-10 Comportamiento promedio de la salinidad y corrientes superficiales (m/s) entre el 22 de mayo y el 12 de junio de 2014 en el Caribe colombiano. CGSM: Ciénaga Grande de Santa Marta. [100] Figura 8-11 Comportamiento promedio de la clorofila-a (mg/ m3) y corrientes superficiales (m/s) entre el 22 de mayo y el 12 de junio de 2014 en el Caribe colombiano. CGSM: Ciénaga Grande de Santa Marta. [100] Figura 8-12 Modelación GAM de relación entre la captura por unidad de esfuerzo -CPUE (Número de individuos/h*100 anzuelos) y los predictores [temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m) y longitud (°)], en el Caribe colombiano. Se muestran en color gris los intervalos de confianza. (+): Abundancia relativa o CPUE integrada de todos los peces grandes pelágicos. [102] Figura 8-13 Modelación GAM de relación entre la captura por unidad de área -CPUA (número de individuos/m2) del ictioplancton y los predictores [temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m) y longitud (°)], en el Caribe colombiano. Se muestran en color gris los intervalos de confianza. [103] Figura 8-14 Comportamiento a profundidad de a) salinidad, b) oxígeno disuelto (%) y c) pH, en el Pacífico colombiano. [103] Figura 8-15 Comportamiento promedio de la altura del nivel del mar (m) y corrientes superficiales (m/s) entre el 20 de noviembre y el 17 de diciembre de 2014 en el Pacífico colombiano. [104] Figura 8-16 Comportamiento promedio de la temperatura superficial del mar (TSM; °C) y corrientes superficiales (m/s) entre el 20 de noviembre y el 17 de diciembre de 2014 en el Pacífico colombiano. [104] Figura 8-17 Comportamiento promedio de la salinidad superficial del mar y corrientes superficiales (m/s) entre el 20 de noviembre y el 17 de diciembre de 2014 en el Pacífico colombiano. [105] Figura 8-18 Comportamiento promedio de la clorofila-a (mg/ m3) y corrientes superficiales (m/s) entre el 20 de noviembre y el 17 de diciembre de 2014 en el Pacífico colombiano. [105] Figura 8-19 Modelación GAM de relación entre la captura por unidad de esfuerzo -CPUE (Número de individuos/h*100 anzuelos) de grandes pelágicos y los predictores: temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m), latitud (°) y longitud (°), en el Pacífico colombiano. Se muestran en color gris los intervalos de confianza. [107] Figura 8-20 Modelación GAM de relación entre la captura por unidad de área -CPUA (Número de individuos/m2) del ictioplancton y los predictores: temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m), latitud (°) y longitud (°), en el Pacífico colombiano. Se muestran en color gris los intervalos de confianza. [107] 9 Figura 9-1 Modelo conceptual del sistema de manejo pesquero para el recurso grandes pelágicos en Colombia. [110] 13 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA CONTENIDO 1 2 3 4 5 6 7 8 9 RESUMEN 10 11 12 13 INTRODUCCIÓN GENERAL 18 19 14 15 16 17 24 25 ENFOQUE METODOLÓGICO 20 21 22 23 COMPOSICIÓN DE LA CAPTURA Y ABUNDANCIA RELATIVA 26 27 28 29 30 31 32 33 14 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 34 35 36 37 38 39 40 41 DISTRIBUCIÓN ESPACIAL DE LA ABUNDANCIA, ESTRUCTURA DE TALLAS Y ESTIMACIÓN DE BIOMASA 42 43 44 45 46 47 48 49 57 ASPECTOS REPRODUCTIVOS DE RECURSOS GRANDES PELÁGICOS 50 51 52 53 54 55 56 58 59 60 61 62 63 64 65 COMPOSICIÓN, ABUNDANCIA Y DISTRIBUCIÓN ESPACIAL DEL ICTIOPLANCTON 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 15 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 82 83 84 85 86 87 88 89 VARIABLES OCEANOGRÁFICAS Y SU RELACIÓN CON LA ABUNDANCIA DEL RECURSO 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 ESTRATEGIAS DE MANEJO PARA LA PESQUERÍA DE GRANDES PELÁGICOS BASADA EN EL ENFOQUE ECOSISTÉMICO 106 CONCLUSIONES 107 108 109 110 111 112 115 116 117 118 119 120 BIBLIOGRAFÍA 114 113 16 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA PRESENTACIÓN El Instituto de Investigaciones Marinas y Costeras –INVEMAR– como parte de su misión en realizar investigación básica y aplicada de los recursos naturales renovables y ecosistemas marinos y oceánicos de interés nacional, dirige esfuerzos para la generación de conocimiento sobre el estado de los recursos marinos y costeros como una herramienta fundamental para soportar la toma de decisiones, mejorar en la gestión y regulación de los recursos. En los últimos años, parte de las acciones de investigación en el tema de pesca han sido requeridas y desarrolladas en cooperación con la Autoridad Nacional de Acuicultura y Pesca –AUNAP–, desde su misión de ejecutar la política pesquera y acuícola en el territorio colombiano con fines de investigación, ordenamiento, administración, control y vigilancia de los recursos pesqueros como respuesta a necesidades de los usuarios de la pesca en las costas del país. Una parte de esta investigación conjunta se ha centrado en la evaluación de los recursos de peces grandes pelágicos (e.g. atunes, dorados y especies afines), motivada por la escasa información disponible y la alta demanda nacional e internacional de este recurso medianamente aprovechado tanto en la región costera como la oceánica del Caribe y Pacífico de Colombia. Este trabajo conjunto dio alcance a la publicación de este libro como un instrumento de amplia divulgación del conocimiento en recursos grandes pelágicos, aportando información bajo un enfoque ecosistémico de aspectos biológico-pesqueros y ecológicos claves para el manejo y conservación de los peces grandes pelágicos por parte del sector ambiental y pesquero del país. Específicamente se hace un recorrido por la composición de especies objeto de pesca de un arte selectivo para este recurso y usado en el país comercialmente como es el palangre horizontal o espinel. Esta información colectada con riguroso diseño muestral y aplicando tecnologías de punta de hidroacústica pesquera y oceanografía pesquera satelital, permitió determinar niveles de abundancia, su distribución espacial e identificar el efecto del ambiente sobre la disponibilidad del recurso. Así mismo, se obtuvo información reciente para el país sobre la estructura de tallas de las poblaciones, su estado reproductivo objeto de pesca y las fases iniciales de historia de vida (composición, densidades y distribución de huevos y larvas de peces). La anterior información fundamenta la naturaleza de esta publicación que aborda conceptos y tópicos de interés, presentan indicadores de estado de aprovechamiento y sugiere medidas operacionales e institucionales de manejo y conservación. FRANCISCO ARMANDO ARIAS ISAZA Director General Instituto de Investigaciones Marinas y Costeras –INVEMAR– EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 1 17 RESUMEN Esta publicación es producto de investigación conjunta entre la AUNAP y el INVEMAR para generar conocimiento de recursos grandes pelágicos, como una herramienta fundamental para el manejo y conservación de los recursos pesqueros en aguas marinas jurisdiccionales en Colombia. Los resultados de la investigación están basados en información independiente de la pesquería, es decir, cruceros de prospección directa en el Caribe continental (mayo-junio de 2014) y Pacífico (noviembre-diciembre de 2014) haciendo uso de embarcaciones pesqueras como plataforma de investigación. Adicionalmente, también fue empleada información procedente de cruceros llevados a cabo en 2013 para la misma área de estudio y similar periodo de prospección. Los alcances y resultados obtenidos en el Caribe y Pacífico se presentan en seis secciones orientadas en tópicos claves de evaluación y manejo integral de pesquerías: I) Composición de la captura y abundancia relativa; II) Distribución espacial de la abundancia, estructura de tallas y estimación de biomasa; III) Aspectos reproductivos; IV) Composición, abundancia y distribución espacial del ictioplancton (huevos y larvas de peces); V) Variables oceanográficas y su relación con la abundancia del recurso y VI) Estrategias de manejo para la pesquería de grandes pelágicos basada en el enfoque ecosistémico. Con base en lo anterior, este libro reúne y consolida los principales aportes de la investigación, presentando así un modelo conceptual de manejo, indicadores pesqueros y estrategias de manejo y conservación que abordan aspectos biológico-pesqueros, ecológicos y tecnológicos, brindando así al sector pesquero y ambiental herramientas de uso para la toma de decisiones y el aprovechamiento sustentable de los recursos grandes pelágicos en Colombia. CARIBE La captura total obtenida fue 8.385 kg, correspondiente a 311 individuos identificados dentro de 16 especies agrupadas en diez familias, siendo la más abundante en peso Scombridae (33,4 %). Las especies más abundantes en peso y número fueron el atún aleta amarilla (Thunnus albacares; 2.635 kg y 61 individuos), el tiburón sedoso (Carcharhinus falciformis; 1.211 kg y 70 individuos), el tiburón azul (Prionace glauca; 1.201 kg y 19 individuos), el marlín azul (Makaira nigricans; 755 kg y 8 individuos), el dorado (Coryphaena hippurus; 690 kg y 58 individuos), entre otros. El análisis de distribución espacial del atún aleta amarilla mostró un parche con densidad alta en la zona sur frente al golfo de Morrosquillo (>171<334 t/mn2) y otro parche de menor tamaño y densidad intermedia (>44<85 t/mn2) frente a Barranquilla en la zona centro. Para esta especie la biomasa instantánea estimada fue 9.200 t, con una densidad media igual a 7,44 t/mn2 y un área efectiva de ocurrencia del recurso de 1.236 mn2. El 90 % de los individuos del atún aleta amarilla fueron adultos, los cuales se encontraron agregados principalmente en la zona norte, exhibiendo tallas de entre 164 y 191 cm de longitud total (LT), mientras que la mayoría de individuos con menores tallas se localizaron en la zona centro y sur. El dorado presentó parches con densidad alta (>61<113 t/ mn2), principalmente en la zona centro frente a Barranquilla. La biomasa estimada fue 1.700 t, con una densidad media igual a 3,21 t/mn2 y un área efectiva de 530 mn2. El dorado exhibió los individuos más grandes principalmente hacia las zonas centro y sur con tallas superiores entre 136 y 152 cm de LT. Respecto a los aspectos reproductivos del atún aleta amarilla, se analizaron 115 individuos, 55 en 2013 y 60 en 2014, con prevalencia en ambos años de peces maduros (estados de madurez IV y V) y capturas similares de hembras y machos. La distribución espacial de la madurez del atún aleta amarilla entre mayo y junio de 2014, mostró la mayoría de los individuos maduros agregados frente de Santa Marta y frente al golfo de Morrosquillo. Para esta especie se propone una escala de madurez gonadal 18 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA macroscópica y microscópica ilustrada, como un valioso aporte al conocimiento de la biología de esta especie e insumo para las medidas de conservación y manejo. En cuanto al ictioplancton, cerca del 50 % de los huevos colectados se encontraron en estado de división o no fecundados, el resto fueron discriminados en tres morfotipos (M): MI (vitelo y/o larva pigmentada), MII (huevo sin pigmento) y MIII (huevo con ornamentación del corión). La distribución espacial de los huevos fue relativamente homogénea en el Caribe, sin embargo, las estaciones que presentaron un mayor valor de densidad estuvieron hacia la zona norte (>90 huevos/10 m2) entre La Guajira y Santa Marta. En general, larvas de peces presentaron un patrón homogéneo en su distribución espacial con una alta densidad en la zona norte (>178 larvas/10 m2) debido la alta abundancia de la familia Clupeidae (20%; 300 larvas/10 m2). Se registraron un total de 42 familias y 87 taxones a nivel de género y especie. Aunque con baja abundancia (2,4%) se encontraron larvas de grandes pelágicos tales como la bacoreta (Euthynnus alleteratus), barrilete (Katsuwonus pelamis), sierra wahoo (Acanthocybium solandri), atún aleta amarilla (T. albacares) y ojo gordo (T. obesus). El análisis oceanográfico evidenció una variabilidad ambiental en el Caribe continental, permitiendo establecer que zonas de alta productividad asociadas a termoclinas estuvieron entre los 50 m y 75 m de profundidad. Se determinó que la temperatura superficial del mar y la clorofila-a fueron predictores significativos de la abundancia relativa de peces grandes pelágicos. PACÍFICO La captura total obtenida fue 5.804 kg, correspondiente a 282 individuos identificados dentro de 11 especies agrupadas en siete familias, entre las cuales destacó la familia Istiophoridae (43,0 %). El análisis en la composición de la captura por zona de muestreo mostró la mayor abundancia en peso y número de individuos en la zona costera, representada principalmente por el pez vela (Istiophorus platypterus; 1.224 kg y 41 individuos), el dorado (C. hippurus; 678 kg y 68 individuos) y tiburón sedoso (C. falciformis; 559 kg y 65 individuos), los cuales también fueron observados en la zona oceánica con alta abundancia en número de individuos, presentando solo el pez vela I. platypterus una alta abundancia en peso. El análisis espacial del dorado mostró solo un parche con densidad alta en la zona costera frente a Buenaventura (>66<125 t/mn2), mientras que en el resto del área evaluada las densidades fueron bajas (<16 t/ mn2). La biomasa estimada fue 3.900 t, con una densidad media igual a 3,6 t/mn2 y un área efectiva de 1.087 mn2. Respecto a la distribución espacial de las tallas de dorado, se encontró a los individuos más grandes al norte de la zona costera e intermedia, con tallas entre 127 y 145 cm de LT; mientras que se observó una tendencia de individuos de menores tallas (>103<117 cm de LT) hacia la zona oceánica. Respecto a los aspectos reproductivos de dorado se encontró la mayoría de peces maduros (estados IV y V) y además se capturó una mayor proporción de hembras que machos. La distribución espacial de la madurez del dorado mostró una agregación de individuos maduros hacia el norte de la zona costera entre Cabo Corrientes y Bahía Solano, mientras que la ocurrencia de individuos inmaduros fue casi nula. El 85,8 % de los individuos capturados se encontraron por encima de la talla media de madurez (TMM) reportada en la literatura de 89 cm de LT. Para el dorado se propone una escala de madurez gonadal macroscópica y microscópica ilustrada, como aporte a la biología reproductiva de la especie y apoyo al diseño de estrategias de manejo y conservación. En cuanto al ictioplancton, la densidad de huevos estuvo entre 75 y 903 huevos/10 m2, siendo los más característicos los morfotipos MI (larva o vitelo pigmentado) y MIII (corion ornamentado). Las mayores densidades de huevos de peces se encontraron en las zona intermedia y oceánica del área de estudio (>500 huevos/10 m2), sin embargo, también se encontraron densidades importantes en la zona costera, registrando huevos en el 100 % de las estaciones muestreadas. En larvas de peces se encontraron densidades entre 70 y 1.145 larvas/10 m2 y al igual que en los huevos, las mayores abundancias de larvas se concentraron en la zona oceánica (> 1.000 larvas/10 m2). Se recolectaron un total de 549 larvas representadas en 54 taxones (género y especie) y 38 familias. La familia más importante en abundancia fue Myctophidae (43,6 %). Tambien se destacaron larvas de importancia comercial como Paralichthydae (lenguados, 1,74 %), Cynoglossidae (lengüetas, 1,30 %), Coryphaenidae (dorado, 0,94 %), Mugilidae (lisas, 0,89 %); Scombridae (atunes y sierras, 0,61 %) y Engraulidae (carduma, 0,27 %). La especie más abundante y frecuente fue Diogenichthys laternatus (pez linterna), especie de hábitat batipelágico encontrada en todas las estaciones de muestreo representando el 18 % de la abundancia total. Se identificaron larvas de dorado C. hippurus con una frecuencia de aparición de 17,65 %, lo que sugiere la ocurrencia de eventos reproductivos para esta especie en el Pacífico. También se capturaron larvas de escómbridos Katsuwonus pelamis y Euthynnus lineatus (barrilete) que son especies pelágicas de interés comercial. En el Pacífico, la oceanografía se caracterizó por la ocurrencia de frentes térmicos asociados con celdas ciclónicas o anticiclónicas que ocurrieron en límites con Panamá demarcando áreas productivas de interés para la pesca. Específicamente, altas abundancias de grandes pelágicos se asociaron a mayores concentraciones de clorofila-a (~0,195 mg/m3) y altura del nivel del mar entre 0,360 y 0,365 m. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 2 19 INTRODUCCIÓN GENERAL Los recursos grandes pelágicos se categorizan como especies transzonales y altamente migratorias con desplazamientos que incluyen aguas costeras y oceánicas alrededor del mundo (FAO, 1994; Maguire et al., 2006). Este recurso es ampliamente aprovechado por pesca, debido al alto valor comercial de especies como los atunes, dorados y picudos, aunque otras especies afines capturadas incidentalmente como las sierras, barracudas, tiburones, entre otros, aportan a los ingresos de estas pesquerías con precios intermedios en los mercados locales y regionales (Maguire et al., 2006). La captura industrial de grandes pelágicos se realiza con diversos artes o aparejos de pesca, en los que destaca el palangre superficial horizontal o longline (línea principal de variada longitud provista de líneas secundarias con anzuelos y carnada) y la red de cerco (mallas que forman un bolso de encerramiento para bancos densos de peces) (Beverly et al., 2003; FAO, 2012). Otros artes y métodos de pesca usados también en pesca artesanal incluyen líneas de mano, redes de enmalle, caña, arpón y correteo. Los reportes globales recientes para este recurso indican una alta explotación y declive considerable en algunas poblaciones como por ejemplo en túnidos (e.g. atún rojo, atún aleta azul), los cuales presentan una tercera parte en estado de sobreexplotación, un 37,5 % en plena explotación y solo un 29 % sin explotar plenamente (FAO, 2012). Específicamente, para el océano Pacífico Oriental que incluye aguas colombianas, el más reciente informe de la Comisión Interamericana del Atún Tropical –CIAT– señala que el estado de explotación de las especies atún aleta amarilla, patudo, barrilete, albacora, aleta azul, pez espada, pez vela, marlín rayado, marlín azul y tiburón sedoso oscila entre moderadamente explotado y en máximo rendimiento sostenible (CIAT, 2015). No obstante, se conoce que la disponibilidad de estas especies no solo está condicionada al efecto de activi- dades antrópicas como la mortalidad por pesca, sino también a la influencia de procesos oceanográficos y bioecológicos de mediana y gran escala (Domokos et al., 2007; Langley et al., 2009; Fromentin et al., 2014). Dada la importancia de los grandes pelágicos como aporte significativo de alimento y generación de ingresos y divisas y, que durante los últimos años ha sido evidente la vulnerabilidad de las poblaciones por efecto de la pesca y el ambiente, en la actualidad son notorios los esfuerzos en materia de monitoreo, evaluación y ordenamiento del recurso llevados a cabo por organizaciones tales como, la CIAT, la Comisión Internacional para la Conservación del Atún del Atlántico y la Organización de las Naciones Unidas para la Alimentación y la Agricultura –FAO–, entre otras. En Colombia, los grandes pelágicos son aprovechados principalmente en el Pacífico, en primera instancia por la flota industrial de bandera extranjera y nacional que opera con redes de cerco, seguido de la flota industrial nacional que opera con espinel o palangre (denominada de pesca blanca) en aguas costa afuera y, en menor grado por embarcaciones de tipo artesanal que trabajan cerca de la costa con espineles más pequeños, líneas de mano y redes de enmalle (Rueda et al., 2012). En el Caribe colombiano, el recurso es aprovechado esencialmente por embarcaciones con palangre asociadas a otros países como el Japón, Corea e Indonesia, aunque en los últimos años han reducido en gran medida su actividad de pesca pasando de cinco embarcaciones en 2006 a solo una para los años 2013 y 2014 (Rueda et al., 2014; Rodríguez et al., 2015). Por su parte, la pesquería artesanal utiliza con menor actividad que en el Pacífico redes de enmalle, palangres pequeños y correteo para la captura de este recurso. Las principales especies objetivo de captura de la flota industrial de palangre son el atún aleta amarilla (Thunnus albacares) y el dorado (Coryphaena 20 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA hippurus); mientras que incidentalmente se capturan diferentes especies de marlín (Makaira nigricans, Kajikia albida, Tetrapturus spp.), pez vela (Istiophorus albicans), pez espada (Xiphias gladius), tiburón sedoso (Carcharhinus falciformis) y tiburón látigo (Alopias spp.), entre otros. Tanto para el Caribe continental como para el Pacífico existe gran incertidumbre sobre los niveles de abundancia, distribución y aspectos de procesos poblacionales de las especies de peces grandes pelágicos aprovechadas por las pesquerías nacionales. Bajo este panorama, en que existe recurso potencialmente aprovechado por pesca, se requiere direccionar esfuerzos de investigación para aumentar el estado de conocimiento como una herramienta fundamental para soportar la toma de decisiones en la pesquería. Por esta razón, desde el año 2013 la Autoridad Nacional de Acuicultura y Pesca –AUNAP– y el Instituto de Investigaciones Marinas y Costeras –INVEMAR–, con la acción participativa del sector pesquero, han venido realizando un trabajo conjunto para valorar las poblaciones objeto de pesca de los grandes pelágicos en ambas costas del país, basado en una evaluación directa a través de cruceros de investigación (datos independientes de la pesquería). Esta fuente de información, ha permitido colectar datos en sus componentes biológico-pesquero, reproductivo, hidroacústico y oceanográfico, de forma rápida, confiable y con una amplia cobertura espacial. Los resultados de estas investigaciones se presentan en esta publicación estructurada en seis secciones orientadas en tópicos claves de evaluación y manejo integral de pesquerías: I: Composición y abundancia relativa de especies, el cual describe y analiza la diversidad de especies que confor- maron la captura objetivo e incidental por zonas de pesca; II: Distribución espacial de la abundancia, estructura de tallas y estimación de biomasa, que establece un primer punto de referencia sobre el estado de la población del atún aleta amarilla (T. albacares) y el dorado (C. hippurus), determinando el potencial pesquero del recurso en términos de biomasa explotable; III: Aspectos reproductivos, como un aporte a la biología reproductiva con base en un análisis macroscópico y microestructural (histología) de las gónadas y la propuesta de una escala de madurez sexual para el atún aleta amarilla y el dorado; IV: Composición, abundancia y distribución espacial del ictioplancton, el cual describe los peces en sus primeras fases de vida (huevos y larvas) que son claves en la historia de vida de los grandes pelágicos; V: Variables oceanográficas y su relación con la abundancia del recurso, este capítulo hace una descripción de la oceanografía (temperatura, clorofila, salinidad, altura del nivel del mar, oxígeno disuelto, entre otras) y cómo estas afectan la disponibilidad de los grandes pelágicos y VI: Estrategias de manejo para la pesquería de grandes pelágicos basada en el enfoque ecosistémico, que presenta indicadores y un modelo conceptual de manejo a partir de la mejor evidencia científica disponible para la toma de decisiones. Esta publicación suministra información confiable y oportuna tanto a la institucionalidad pesquera y ambiental, como a la academia, investigación y en especial a los gremios, asociaciones y pescadores de ambas costas en su calidad de usuarios directos del recurso aumentando el conocimiento del recurso grandes pelágicos del país. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 21 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 3 23 ENFOQUE METODOLÓGICO ÁREA DE ESTUDIO CARIBE El área de estudio comprendió desde Cabo de La Vela, departamento de La Guajira (13° 3,825’ N – 72° 52,404’ W) hasta el frente del golfo de Urabá, departamento de Antioquia (09° 26,732’ N – 76° 51,412’ W), cubriendo la región oceánica entre 30 y 85 millas náuticas (mn) de distancia de la costa [Figura 3-1 ]. El área de muestreo fue categorizada siguiendo un patrón de influencia marina variable de Norte a Sur por influencia de ríos y áreas estuarinas, en las siguientes zonas: norte (estaciones ubicadas entre La Guajira y Santa Marta); centro (estaciones ubicadas entre Santa Marta y Cartagena) y sur (estaciones ubicadas entre Cartagena y el golfo de Urabá). El Caribe colombiano se caracteriza por el desarrollo de afloramientos estacionales o surgencias costeras, el giro ciclónico de Panamá (GCP) y la contracorriente Panamá-Colombia (CCPC) (Molinari et al., 1980; Andrade y Barton, 2005; Bernal et al., 2006). En La Guajira (zona norte), los vientos Alisios son más fuertes hacia el final e inicio del año, lo que induce la surgencia costera de aguas más frías y ricas en nutrientes que favorecen la oferta de alimento (Andrade, 1993; Andrade et al., 2003; Andrade y Barton, 2005; Franco-Herrera, 2005) y la disponibilidad de recursos pesqueros de importancia comercial (Páramo et al., 2011; Rodríguez y Páramo, 2012). Por el contrario, el área del Darién (zona sur) cerca a Panamá, está más resguardada de los vientos alisios, por lo que allí domina el régimen del GPC, siendo las aguas más cálidas y presentándose oscilaciones intraanuales menores, debido a las descargas de los ríos que generan una termoclina superficial que inhibe la mezcla vertical (Bernal et al., 2006; Ruiz-Ochoa, 2011). Figura 3-1 Mapa del área de estudio, ubicando las estaciones de muestreo divididas en tres zonas (norte, centro y sur) durante el crucero de investigación en peces grandes pelágicos entre mayo y junio de 2014 en el Caribe colombiano. Fuente INVEMAR – LabSIS, 2015. 24 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA PACÍFICO El área de estudio comprendió, en sentido latitudinal (Norte-Sur), desde el frente de Cabo Huina en el Chocó (05° 54,268’ N 78° 10,213’ W) hasta la isla Gorgona en el Valle del Cauca (03° 43,9932’ N - 78° 15,503’ W), mientras que en sentido longitudinal (Este-Oeste) desde las 30 mn de la costa hasta la isla de Malpelo [Figura 3-2 ]. El área de muestreo fue categorizada por las siguientes zonas: costera (estaciones ubicadas por lo menos a 30 mn la franja costera); intermedia (estaciones ubicadas a 100 mn costa afuera) y oceánica (estaciones ubicadas cerca de la isla Malpelo, superior a 250 mn costa afuera). El Pacífico colombiano presenta diferentes condiciones oceanográficas a lo largo de la costa que la delimitan geográficamente. La zona norte se extiende desde los límites con Panamá hasta Cabo Corrientes en el Chocó y se caracteriza por la presencia de acantilados, una plataforma profunda y estrecha de aproximadamente uno o dos millas náuticas con sustrato predominantemente rocoso, aunque también se identifican arenas carbonatadas biogénicas y arenas lodosas (Cantera y Contreras, 1993; Díaz et al., 1997). La zona centro y sur, entre Cabo Corrientes y Pasacaballos en Nariño, presenta una plataforma más extensa y rellena de sedimentos marinos, en su mayoría de origen fluvial provenientes de numerosos ríos que desembocan en la costa, lo cual determina que la costa sea baja con playas arenosas, así como con extensas formaciones de manglares en áreas estuarinas (Díaz et al., 1997). La Zona de Convergencia Intertropical (ZCIT) es la que regula el clima del Pacífico, con alta nubosidad Figura 3-2 Mapa del área de estudio, ubicando las estaciones de muestreo divididas en tres zonas (costera, intermedia y oceánica) durante el crucero de investigación en peces grandes pelágicos entre noviembre y diciembre de 2014 en el Pacífico colombiano. Fuente INVEMAR – LabSIS, 2015. e intensas lluvias generadas por la inestabilidad de las masas de aire húmedo y caliente que allí se presentan (Tchantsev y Cabrera, 1998; Poveda et al., 2001). Al inicio del año se crea la divergencia del agua costa afuera debido a los patrones de circulación atmosférica en el área oceánica y a la surgencia en el golfo de Panamá que incrementan la productividad (Rodríguez-Rubio y Stuardo, 2002; Rodríguez-Rubio et al., 2003). La salinidad muestra estar directamente afectada por la precipitación y el aporte de los ríos; mientras en enero aumenta, debido a la entrada de aguas oceánicas, en el periodo marzo-junio disminuye influenciada por la precipitación (marzo), y el aporte de los ríos (mayo-junio) registrando salinidades entre 25 y 28 (Tchantsev y Cabrera, 1998). La isla Malpelo, que está ubicada a 500 km al oeste de Buenaventura (Valle del Cauca) (04°0’00” N, 81°3’30” W), es la cúspide de una cordillera sumergida que se levanta paralela a la costa continental, desde una profundidad de 4.000 m. La isla es de origen volcánico y exhibe acantilados muy escarpados en casi todos sus márgenes y tiene 1,2 km2 de superficie emergida (Chase, 1968; Stead, 1975; Prahl, 1990). El clima que se encuentra bajo la influencia de la franja ecuatorial de la ZCIT, es predominantemente cálido, húmedo y con precipitaciones durante la mayor parte del año (Prahl, 1990; Brando et al., 1992), albergando, además, diversas y abundantes comunidades de especies de peces de alto valor pesquero (Rubio et al., 1992; Solano y Hernández, 1998). 25 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA DISEÑO DE MUESTREO DETECCIÓN HIDROACÚSTICA Se realizaron dos cruceros de investigación para evaluación de recursos grandes pelágicos, uno en el Caribe entre mayo y junio de 2014 y otro en el Pacífico entre noviembre y diciembre de 2014. Para los cruceros se utilizó como plataforma de investigación embarcaciones convencionales de la flota industrial con palangre de operación en cada área de estudio [Tabla 3-1 ]. Se empleó en cada crucero un diseño de muestreo sistemático en una grilla regular de 18 transectos en forma de zig-zag separados cada 30 mn aproximadamente. Durante la navegación en la grilla de muestreo se realizaron detecciones hidroacústicas de peces mediante una ecosonda SIMRAD ES70 (split beam) con transductor de frecuencia 38 kHz y además, se contó con el apoyo del sistema cartográfico y de navegación OLEX®. La ecosonda científica proporciona una tecnología de investigación pesquera muy utilizada para la evaluación directa de recursos marinos, la cual trabaja bajo los principios del sonido, es decir, que se transmiten pulsos eléctricos mediante una unidad transceptora GPT (General Purpose Transceiver, por su siglas en inglés), los cuales son transformados por el transductor en pulsos u ondas de sonido que se propagan a través de la columna del agua y que son recibidos en forma de ecos al chocar con los peces y el fondo marino. Todos los ecos recibidos se visualizaron en un monitor como imágenes digitales denominadas ecogramas, mostrando en tiempo real y con buena definición las detecciones hidroacústicas en la columna del agua [Tabla 3-1 ]. Tabla 3-1 Muestreo hidroacústico durante los cruceros de investigación de grandes pelágicos en el Caribe y Pacífico colombiano. Caribe Pacífico Plataforma de investigación: Embarcaciones convencionales de pesca con palangre horizontal industrial o longline. Fuente imagen: Programa VAR - Invemar. Monitor de la ecosonda SIMRAD ES70: Permite la visualización de ecogramas con información de las detecciones de peces y el fondo marino. Sistema Cartográfico y de Navegación OLEX: Este equipo facilita la adquisición de datos batimétricos y la navegación en crucero. Fuente imagen: Programa VAR - Invemar. Unidad Transceptora de frecuencia 38 kHz: Contiene los circuitos requeridos para transmitir y recibir señales eléctricas en la frecuencia de trabajo de la ecosonda. Fuente imagen: Manual SIMRAD (2003). Transductores Split Beam de frecuencia 38 kHz: Transforma la energía eléctrica en pulsos u ondas sonoras. Interpreta el tiempo transcurrido entre la emisión y recepción del pulso y la intensidad con que los recibe. Fuente imagen: Manual SIMRAD (2003). Esquema de señal hidroacústica: Transmisión de pulso y recepción del eco al chocar y rebotar con el fondo marino y los peces. Fuente imagen: Adaptada y modificada de Manual SIMRAD (2003). 26 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA PESCA EXPLORATORIA En total se realizaron 35 estaciones de pesca exploratoria, 18 estaciones en el Caribe [Tabla 3-2 ] y 17 en el Pacífico [Tabla 3-3 ]. En las estaciones de pesca se empleó un palangre superficial horizontal o longline con características técnicas propias de los artes o aparejos empleados por la flota pesquera de grandes pelágicos en cada área de estudio [Tabla 3-4 y Tabla 3-5 ]. Un palangre está definido como un aparejo compuesto por una línea principal de gran longitud, de la cual van unidas líneas secundarias con anzuelos y carnada (cebo) para atraer los peces. Posición inicial Para estandarizar el esfuerzo de pesca se empleó, en lo posible en todas las estaciones, igual número de anzuelos y tiempo de reposo del palangre en el agua, contando el tiempo a partir del último anzuelo lanzado al agua. En el crucero del Pacífico, el número de anzuelos en las últimas estaciones fue variable (entre 350 y 500 anzuelos), debido a la limitante en la disponibilidad de carnada a final del muestreo. No obstante, este factor de variabilidad fue controlado al estandarizar los cálculos de abundancia a un número de 100 anzuelos. Posición final Estación de pesca Latitud (N) Longitud (W) Latitud (N) Longitud (W) EP01 12°42,373 72°54,610 13°03,628 72°52,602 EP02 12°42,148 72°53,712 12°48,968 73°14,790 EP03 12°31,378 73°27,747 12°50,550 73°22,120 EP04 12°27,793 73°27,132 12°35,799 73°50,060 EP05 12°14,293 73°53,470 12°34,879 73°51,436 EP06 12°13,809 74°05,175 12°20,234 74°25,770 EP07 11°59,549 74°27,268 12°20,659 74°27,146 EP08 11°57,750 74°37,035 12°04,895 74°55,808 EP09 11°42,400 74°57,405 11°40,576 75°16,925 EP10 11°28,731 75°18,908 11°27,787 75°37,176 EP11 11°22,887 75°28,282 11°18,948 75°46,048 EP12 11°03,095 75°43,771 10°51,107 75°54,077 EP13 10°55,549 75°56,781 10°51,108 76°16,110 EP14 10°28,758 76°15,616 10°47,005 76°08,854 EP15 09°53,639 76°26,683 09°36,276 76°35,320 EP16 09°53,285 76°29,791 09°56,095 76°47,335 EP17 09°58,854 76°31,419 10°15,189 76°38,624 EP18 10°22,053 76°13,597 10°24,664 76°31,437 Estación de pesca Latitud (N) Longitud (W) Latitud (N) Longitud (W) EP01 04°34,610 80°30,704 04°47,338 80°23,657 EP02 04°16,442 80°24,340 04°17,093 80°33,891 EP03 03°46,525 80°17,716 03°50,417 80°28,413 EP04 03°26,407 80°20,555 03°37,772 80°28,475 EP05 03°10,182 80°34,648 02°59,761 80°28,252 EP06 02°56,484 82°29,638 03°04,724 82°42,961 EP07 03°28,026 82°30,824 03°38,928 82°41,280 EP08 04°02,973 82°37,828 03°55,580 82°28,788 EP09 04°16,773 82°23,055 04°21,011 82°32,537 EP10 04°34,632 82°34,184 04°46,753 82°32,883 Posición inicial Posición final EP11 05°54,268 78°10,213 06°05,014 78°16,215 EP12 05°49,803 78°13,048 05°42,156 78°22,828 EP13 05°36,655 78°30,182 05°29,067 78°22,001 EP14 05°28,399 78°22,977 05°20,813 78°12,462 EP15 04°51,638 78°30,477 04°43,517 78°25,084 EP16 04°07,591 78°29,936 04°03,222 78°26,092 EP17 03°43,932 78°15,503 03°38,202 78°18,285 Zona Norte Tabla 3-2 Coordenadas geográficas de las estaciones de pesca exploratoria (EP) con palangre superficial horizontal durante el crucero de investigación en peces grandes pelágicos en el Caribe colombiano. Centro Sur Zona Intermedia Oceánica Costera Tabla 3-3 Coordenadas geográficas de las estaciones de pesca exploratoria (EP) con palangre horizontal de superficie durante el crucero de investigación de peces grandes pelágicos en el Pacífico colombiano. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Características principales del palangre Palangre horizontal compuesto por una línea principal de 38,4 km de longitud (20,7 mn) de material poliamida monofilamento. Un total de 768 líneas secundarias del mismo material con 15 m de longitud y compuestas cada una por anzuelos circulares calibre 13/0 (tipo atunero) y espaciadas cada una a 50 m. La carnada empleada fue la cachorreta (Auxis rochei). El tiempo de reposo del palangre fue de cinco horas, contando el tiempo a partir del último anzuelo lanzado al agua. Para el seguimiento y rápida ubicación del palangre en el agua fueron dispuestas seis radioboyas a lo largo de la línea principal. La profundidad relativa de trabajo del palangre y acción de los anzuelos fue de 50 m aproximadamente (cerca de la termoclina). Tabla 3-4 Características técnicas del palangre o longline empleado durante el crucero de grandes pelágicos en el Caribe colombiano. Esquema general del palangre Características principales del palangre Palangre horizontal compuesto por una línea principal que varió entre estaciones de muestreo entre 7-10 km de longitud (3,8-5,4 millas náuticas) y de material poliamida monofilamento. Se emplearon entre 350 y 500 líneas secundarias del mismo material con 6 m de longitud y compuestas cada una por anzuelos circulares calibre 14/0 (tipo atunero) y espaciados cada uno a distancia de 20 m. La carnada empleada fue una mezcla calamar (Loligo sp) y peces vivos. El tiempo de reposo del palangre fue de cuatro horas, contando el tiempo a partir del último anzuelo lanzado al agua. Para el seguimiento y ubicación del palangre en el agua fueron dispuestos banderines a lo largo de la línea principal. La profundidad relativa de trabajo del palangre y acción de los anzuelos fue de 20 m aproximadamente. Esquema general del palangre Tabla 3-5 Características técnicas del palangre o longline empleado durante el crucero de grandes pelágicos en el Pacífico colombiano. 27 28 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA En general, la captura objetivo (atunes y dorados) y fauna acompañante (picudos, tiburones, entre otros) fue procesada a bordo determinando para cada especie identificada el peso, talla, sexo y madurez sexual [Tabla 3-6 ]. La identificación de las especies Caribe se realizó con base en guías especializadas en taxonomía de peces (e.g. Compagno, 2002; Collette, 2002) y fichas técnicas para mayor facilidad de identificación de especies y toma de medidas morfométricas en campo. Pacífico Tabla 3-6 Muestreo biológicopesquero durante los cruceros de investigación de grandes pelágicos en el Caribe y Pacífico colombiano. Maniobras de pesca con palangre superficial horizontal o longline. Fuente imagen: Programa VAR – Invemar Procesamiento de la captura a bordo para toma de información biológico-pesquera. Fuente imagen: Programa VAR – Invemar La determinación de la madurez sexual macroscópica para hembras de túnidos se realizó siguiendo la escala macroscópica propuesta por Figueiredo et al. (2008) [Tabla 3-7 ], mientras que en el dorado y demás especies se tuvo en cuenta la escala general de desovadores parciales propuesta por Holden y Raitt (1975) [Tabla 3-8 ]. Para posterior validación en laboratorio de los estados de madurez determinados macroscópicamente, se Tabla 3-7 Escala de madurez sexual para hembras de túnidos propuesta por Figueiredo et al. (2008). Estado de madurez efectuó un registro fotográfico de cada gónada detallando con rótulo la escala métrica, el número de la estación de muestreo y el código identificador de la gónada correspondiente a cada individuo [Tabla 3-9 ]. Cada gónada fue conservada y fijada en formalina al 4 %, con el fin de valorar la condición reproductiva mediante histología. Descripción Inmaduro (I) Las gónadas tienen un color rosado pálido y no se distinguen ovocitos. En Maduración (II) Las gónadas son más grandes respecto al Estado I y presentan un color rosado. Ovocitos pequeños y vasos sanguíneos muy delgados se podrían observar a simple vista. Desove (III) Las gónadas ocupan por lo menos 2/3 de la cavidad abdominal y pueden observarse muchos ovocitos traslúcidos a simple vista. Las gónadas son de color naranja-amarillo. Desovado (IV) Las gónadas son flácidas y presentan aspecto sanguinolento (vasos sanguíneos dilatados). 29 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Tabla 3-8 Escala de madurez sexual general para desovadores parciales propuesta por Holden y Raitt, (1975). Tabla 3-9 Registro fotográfico de las gónadas durante los cruceros de investigación de grandes pelágicos en el Caribe y Pacífico colombiano. Fuente imagen: Programa VAR - Invemar. Estado de madurez Descripción Inmaduro (I) Ovarios y testículos cerca de 1/3 de la cavidad abdominal. Ovarios rosáceos, traslúcidos y huevos invisibles a simple vista; testículos blancuzcos. Virgen madurando y recuperado (IIR) Ovarios y testículos cerca de 1/2 de la longitud de la cavidad abdominal. Ovarios rosáceos, traslucidos, huevos invisibles a simple vista; testículos blancuzcos, más o menos simétricos. Madurando (III) Ovarios y testículos cerca de 2/3 de la longitud de la cavidad abdominal. Ovarios de color rosáceo amarillo aspecto granular; testículos blancuzcos a crema. No hay huevos transparentes o traslucidos visibles. Maduro (IV) Ovarios y testículos ocupan de 2/3 a toda la longitud de la cavidad abdominal. Ovarios de color naranja con vasos sanguíneos superficiales visibles. Grandes huevos maduros y transparentes. Testículos blancuzcos crema y blandos. Desove (V) Ovarios y testículos contraídos cerca de la mitad de la longitud de la cavidad abdominal. Paredes flojas. Los ovarios pueden contener restos de huevos opacos, maduros, en desintegración, oscurecidos a traslúcidos. Testículos sanguinolentos y flácidos. Caribe Pacífico 30 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA OCEANOGRAFÍA Una parte de la información oceanográfica fue obtenida con mediciones in situ de la temperatura, salinidad, oxígeno disuelto y pH, usando una sonda multiparamétrica CTDO . Otra fuente de información oceanográfica se basó en datos superficiales de temperatura (TSM), salinidad, altura del nivel del mar (ANM), clorofila-a (CLA) y corrientes marinas (componente zonal U: sentido Este-Oeste y meridional V: sentido Norte-Sur), los cuales fueron adquiridos mediante sensoramiento remoto satelital y salidas de modelos. La información satelital y modelada se descargó del portal de Internet [MyOcean – Monitoring and Forecasting (www.myocean.eu)]. Para el muestreo de ictioplancton (huevos y larvas de peces) en cada estación de muestreo se realizaron arrastres oblicuos con una red tipo bongo (500 micras con flujómetro HidroBios previamente calibrado) durante un tiempo efectivo de 10 minutos y una velocidad entre 2 y 3 nudos (Smith y Richardson, 1979) . Durante el muestreo de ictioplancton también fue registrada información como la longitud del cable lascado, el ángulo de inclinación del cable (en lo posible debe estar cercano 45°), hora inicial y final del descenso y ascenso de la red, conteo inicial y final del flujómetro. Una vez colectada la muestra de ictioplancton, se almacenó en un frasco de 1 litro con solución de 100 ml de formol tamponado al 37 %, lavando el colector de la red sobre el mismo frasco para evitar pérdida de muestras, hasta completar un litro de volumen reduciendo así la concentración de formol al 4 % [Tabla 3-10 ]. Caribe Tabla 3-10 Muestreo oceanográfico durante los cruceros de investigación de grandes pelágicos en el Caribe y Pacífico colombiano. Pacífico Muestreo de variables oceanográficas con sonda multiparámétrica CTDO. Fuente imagen: Programa VAR-Invemar. Ángulo 45° de inclinación Red bongo Cable lascado Red bongo Profundidad máxima de arrastre Esquema general del muestreo de ictioplancton con red tipo bongo. Fuente imagen: Tomada y adaptada de Aceves y Saldierna, 2013. Muestreo de ictioplancton con red tipo bongo. Fuente imagen: Programa VAR-Invemar. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 4 33 COMPOSICIÓN DE LA CAPTURA Y ABUNDANCIA RELATIVA Beatriz Mejía-Mercado, José Correa, Harold Castillo, Fabián Escobar, Jorge Viaña, Alfredo Rodríguez y Mario Rueda INTRODUCCIÓN Históricamente, la pesca ha sido considerada una de las actividades de mayor tradición cultural y fuente importante de alimento y sustento económico para los asentamientos humanos, sobre todo las comunidades costeras y ribereñas (FAO, 2012). En Colombia, esta actividad ha sido realizada continuamente por estas comunidades mediante la utilización de diferentes artes de pesca (Rubio, 1986; Medina, 2002; Caldas, 2002). Sin embargo, han sido pocos los trabajos que continuamente reportan datos que ayuden a caracterizar las capturas para conocer su composición y abundancia en el tiempo y espacio, incluso mucha de esta información se encuentra en informes técnicos no publicados o con niveles de resolución taxonómica muy bajos. El análisis de la composición de la captura ayuda a referenciar las especies objetivo y aquellas que son capturadas de forma incidental en una pesquería dada. En Colombia, los pocos estudios que valoran la composición de la captura se han enfocado a pesquerías costeras y principalmente aquellas que describen la estructura de la comunidad demersal (e.g. Medina, 2002; Duarte et. al., 2006; Herazo et. al., 2006; Páramo et. al., 2009), pero no existe este tipo de estudio enfocado al recurso de grandes pelágicos. Por otro lado, la captura por unidad de esfuerzo (CPUE) como el índice de abundancia relativa más usado (Haggarty y King, 2006; King, 2013) puede aportar información relevante sobre cambios en el estado de la población en tiempo y espacio. Aunque la CPUE ha sido definida como un índice poco preciso debido que puede ser influenciado por otros factores tales como la capturabilidad y comportamiento del recurso, entre otros (Harley et. al., 2001; Haggarty y King, 2006), utilizar este índice estandarizado permite manejar de forma más adecuada las variaciones que no están relacionadas a la abundancia, posibilitando el uso efectivo de la CPUE para el monitoreo y evaluación de las comunidades de peces infiriendo cambios en las abudancias (Haggarty y King, 2006). Asimismo, el uso de datos de CPUE derivados de cruceros de investigación estandarizados provee una buena medida de la abundancia relativa, mientras que el uso de artes de líneas y anzuelo puede ser una herramienta efectiva y simple de obtener una medida de abundacia relativa de recursos grandes pelágicos (Haggarty y King, 2006). El propósito de este capítulo es entonces describir la composición y la abundancia relativa de peces grandes pelágicos en el Caribe continental y Pacífico colombiano con base en la información colectada en cruceros de investigación independientes de la pesca durante el año 2014. MÉTODOS ESPECÍFICOS La captura total de peces obtenida en las estaciones de muestreo (EP) fue analizada mediante gráficas de barras para describir su composición por familias y especies de grandes pelágicos. La abundancia relativa fue definida en términos de la captura por unidad de esfuerzo –CPUE–, usando una función que estandarizó la captura en número respecto al tiempo (t) de reposo del palangre en el agua y al número de anzuelos (a) empleados en cada crucero (Maunder y Punt, 2004; Haimovici y Ávila Da Silva, 2007): Ci CPUE= 100 t*a * 34 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Donde Ci es la captura en número de individuos (N), t es el tiempo promedio (h) de reposo del palangre en el agua, el cual se obtuvo a partir del tiempo transcurrido entre el último anzuelo lanzado el agua y el primer anzuelo recogido, y a es el número de anzuelos empleados [Tabla 4-1 ]. La CPUE se expresó como el número de individuos capturados por cada 100 anzuelos en una hora para el palangre horizontal o longline. Para observar tendencias espaciales, la captura fue analizada categorizando cada área de estudio por zonas con delimitación geográfica definida (para mayor detalle ver sección 3). Para determinar diferencias estadísticas de la captura multiespecies (peso y número) y la abundancia relativa (CPUE) entre zonas, se realizaron análisis de varianza no paramétricos (test de Kruskal-Wallis) y paramétricos (ANOVA a una vía) teniendo en cuenta el cumplimiento de los supuestos de homogenidad de varianza y normalidad de los residuos con nivel de significancia estadística p<0,05. Todas las estimaciones se realizaron utilizando el software R (Development Core Team, 2015). Con el objeto de describir asociaciones entre especies o agupaciones con alguna medida de afinidad, se aplicó el índice de Morisita-Horn (Magurran, 2013). Para esto se construyeron matrices de abundancia en número (CPUE), excluyendo los grupos que aportaron menos del 2 % de las densidades totales para eliminar especies raras (Field et al., 1982). Posteriormente, la matriz de similitud fue usada para realizar un análisis de conglomerados mediante agrupamiento jerárquico utilizando el promedio ponderado de los grupos, ya que produce poca distorsión a las afinidades originales (Herrera-Moreno, 2000). RESULTADOS Y DISCUSIÓN CARIBE La captura total obtenida fue 8.385 kg, correspondiente a 311 individuos identificados dentro de 16 especies agrupadas en diez familias, siendo las más abundantes en peso Scombridae (33,4 %) y Carcharhinidae (32,6 %) [Figura 4-1 ]. Esto concuerda con lo encontrado por Rueda et al. (2014) en el área de estudio, donde las familias más abundantes fueron Scombridae (46,6 %) y Carcharhinidae (21,9 %) para una captura total de 3.175 kg. Las especies más abundantes en peso y número fueron el atún aleta amarilla (Thunnus albacares; 2.635 kg y 61 individuos), el tiburón sedoso (Carcharhinus falciformis; 1.211 kg y 70 individuos), el tiburón azul (Prionace glauca; 1.201 kg y 19 individuos), el marlín azul (Makaira nigricans; 755 kg y 8 individuos), el dorado (Coryphaena hippurus; 690 kg y 58 individuos), el pez espada (Xiphias gladius; 573 kg y 26 individuos) y el pez vela (Istiophorus albicans; 563 kg y 34 individuos). Rueda et al. (2014), también han reportado para la zona el atún aleta amarilla como la especie más abundante de la captura obtenida durante 2013. La composición de la captura por zona de muestreo, mostró la mayor abundancia en número en la zona sur (N=130), representada principalmente por el tiburón sedoso (N=35) y el dorado (N=24), mientras que la zona norte presentó la menor abundancia (N=87), destacando al atún aleta amarilla (N=19) y el dorado (N=14) [Figura 4-2 ]. Área de estudio Caribe Pacífico* El análisis estadístico mostró diferencias significativas en la captura en número en al menos una de las zonas (F 2,15= 4,3, p-valor <0,05), siendo específica la diferencia entre las zonas sur y norte [Figura 4-3a ]. La captura en peso no mostró diferencias significativas entre zonas (F2,15= 0,6, p-valor >0,05), aunque se observó una tendencia de encontrar menor captura en la zona centro (2.380 kg) y mayor captura en la zona sur (3.037 kg) [Figura 4-3b ], aportada principalmente por las especies atún aleta amarilla (789 kg) y el marlín azul (605 kg). 2,0% 1,0% Scombridae Carcharhinidae 6,8% Istiophoridae 8,2% 33,4% Coryphaenidae Xiphiidae 8.385 kg 15,7% Sphyrnidae Sphyraenidae Lamnidae Dasyatidae 32,6% Alepisauridae Figura 4-1 Composición porcentual de la captura en peso a nivel de familias durante el crucero de investigación de peces grandes pelágicos en el Caribe colombiano. Tipo y calibre de anzuelo Circular 13/0 Distancia entre anzuelos (m) 50 Número de anzuelos 768 Tiempo de reposo (h) 5 Circular 14/0 20 350-500 4 * El número de anzuelos empleados varió de acuerdo con la disponibilidad de carnada para algunas estaciones de muestreo. Tabla 4-1 Número de anzuelos y tiempo de reposo empleado durante los cruceros de investigación en peces grandes pelágicos en el Caribe y Pacífico colombiano. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA a ZONA NORTE 141 150 127 12 85 70 156 55 2.968 kg 428 1.044 N=87 697 b ZONA CENTRO 26 142 164 96 25 45 256 2.380 kg 387 802 N=94 434 c ZONA SUR 294 311 396 14 80 70 3.037 kg 789 N=130 463 605 19 Thunnus albacares Atún aleta amarilla 11 Prionace glauca Tiburón azul 8 Carcharhinus falciformis Tiburón sedoso 10 Istiophorus albicans Pez vela 2 Makaira nigricans Marlin azul 14 Coryphaena hippurus Dorado 8 Acanthocybium solandri Sierra Wahoo 3 Xiphias gladius Pez espada 1 Galeocerdo cuvier Tiburón tigre 7 Sphyraena barracuda Barracuda 1 Kajikia albida Marlin blanco 1 Dasyatis violacea Raya 1 Katsuwonus pelamis Bonito rayado 1 Alepisaurus spp Pez dragón 21 Thunnus albacares Atún aleta amarilla 7 Prionace glauca Tiburón azul 27 Carcharhinus falciformis Tiburón sedoso 20 Coryphaena hippurus Dorado 1 Sphyrna mokarran Tiburón martillo 2 Galeocerdo cuvier Tiburón tigre 6 Istiophorus albicans Pez vela 5 Sphyraena barracuda Barracuda 2 Acanthocybium solandri Sierra Wahoo 2 Xiphias gladius Pez espada 1 Alepisaurus spp Pez dragón 21 Thunnus albacares Atún aleta amarilla 6 Makaira nigricans Marlin azul 21 Xiphias gladius Pez espada 35 Carcharhinus falciformis Tiburón sedoso 6 Istiophorus albicans Pez vela 24 Coryphaena hippurus Dorado 1 Galeocerdo cuvier Tiburón tigre 1 Prionace glauca Tiburón azul 2 Acanthocybium solandri Sierra Wahoo 1 Isurus paucus Tiburón mako aletón 1 Dasyatis violacea Raya Figura 4-2 Composición de la captura en número de individuos (N) y peso (kg) para las zonas de muestreo: a) norte, b) centro y c) sur, en el Caribe colombiano. 35 36 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA a b 700 600 20 PESO (Kg) 15 500 400 300 200 10 Figura 4-3 Comparación de la captura total en a) número de individuos y b) peso por zona de muestreo de grandes pelágicos en el Caribe colombiano. 100 ZONA NORTE ZONA CENTRO ZONA SUR ZONA NORTE ZONA CENTRO ZONA SUR ZONA DE MUESTREO La abundancia relativa o CPUE promedio (± DS) en toda el área de estudio fue 0,45 N/100 anzuelos*h (± 0,14). Las estaciones de pesca que presentaron mayor CPUE fueron en su orden la EP13 (0,76 N/100 anzuelos*h), EP16 (0,60 N/100 anzuelos*h), EP17 (0,57 N/100 anzuelos*h) y EP18 (0,55 N/100 anzuelos*h) ubicadas en la zona sur, seguidas de la EP11 y EP08 (0,55 N/100 anzuelos*h) ubicadas en la zona centro; mientras que las estaciones de menor CPUE fueron la EP10 (0,18 N/100 anzuelos*h) y EP09 (0,23 N/100 anzuelos*h), ubicadas también en la zona centro [Figura 4-4 ]. La abundancia relativa o CPUE mostró diferencias significativas en al menos una de las zonas de muestreo (F2,15= 4,2, p-valor <0,05). Estas diferencias se encontraron entre la zona sur-norte y sur-centro [Figura 4-5 ], aportadas principalmente por el tiburón sedoso, el dorado y el pez espada con una contribución del 50,31 % y con los valores de CPUE más altos para la zona sur. Rueda et al. (2014) reportaron para el mes de mayo de 2013 que la mayor abundancia de grandes pelágicos se localizó hacia la zona sur (1.382 kg), lo cual coincide con los resultados de este estudio (mayo-junio/2014), donde la mayor abundancia se encontró principalmente en la zona sur del Caribe (3.037 kg). Sin embargo, en ambos estudios también se evidenció alta abundancia en la zona norte, donde las condiciones oceanográficas (e.g. eventos de surgencia) juegan un rol importante en la disponibilidad del recurso pesquero (Bula-Meyer, 1985; Andrade y Barton, 2005; Manjarrés-Martínez et al., 2010). Lo anterior confirma que la zona norte y sur del Caribe son áreas con potencial de pesca de grandes pelágicos, cuyas especies CAPTURA (Kg) CPUE (N/100 anzuelos*h) 0,76 800 0,8 700 0,60 CAPTURA (Kg) 600 0,55 0,47 500 400 0,39 0,34 0,42 0,55 0,47 0,47 0,47 0,7 0,57 0,55 0,5 0,44 0,36 0,4 0,29 300 0,3 0,23 0,18 200 0,2 0,1 100 0 0,6 CPUE (N/100 anzuelos*h) NÚMERO DE INDIVIDUOS 25 EP01 EP02 EP03 EP04 ZONA NORTE EP05 EP06 EP07 EP08 EP09 EP10 EP11 EP12 ZONA CENTRO EP13 EP14 EP15 EP16 EP17 EP18 0,0 ZONA SUR ESTACIONES POR ZONAS DE MUESTREO Figura 4-4 Comportamiento de la captura (kg) y la captura por unidad de esfuerzo (CPUE: N/100 anzuelos*h) en cada estación (EP) y zona de muestreo en el Caribe colombiano. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 37 CPUE (N/100 ANZUELOS*h) 0,7 0,6 0,5 0,4 0,3 Figura 4-5 Comparación de la captura por unidad de esfuerzo -CPUA (Número de individuos/100 anzuelos*h) por zona de muestreo de grandes pelágicos en el Caribe colombiano. 0,2 ZONA NORTE ZONA CENTRO ZONA SUR ZONA DE MUESTREO son altamente migratorias y por tanto, tienen preferencias por hábitats con condiciones oceanográficas particulares que favorecen los eventos reproductivos y/o de reclutamiento (Manjarrés-Martínez et al., 2010). El análisis de clasificación mediante el coeficiente de similitud de Morisita-Horn evidenció la formación de tres asociaciones de peces partir del umbral de similitud del 50 % aproximadamente: (I) las especies P. glauca (tiburón azul), A. solandri (sierra wahoo) y S. barracuda (barracuda); (II) las especies I. albicans (pez vela), T. albacares (atún aleta amarilla) y X. gladius (pez espada) y (III) las especies C. falciformis (tiburón sedoso), C. hippurus (dorado) y M. nigricans (marlín azul) [Figura 4-6 ]. Asimismo, se destaca la mayor asociatividad de las especies del grupo II con las del grupo III [Figura 4-6 ]. SIMILITUD 0.96 0.84 0.72 0.60 0.48 0.36 0.24 Tiburón azul (P. glauca) Sierra Wahoo (A. solandri) Barracuda (S. barracuda) I Pez vela (I. albicans) Atún aleta amarilla (T. albacares) Pez espada (X. gladius) II Tiburón sedoso (C. falciformis) Dorado (C. hippurus) Marlin (M. nigricans) III Figura 4-6 Análisis de clasificación de las especies calculado con el coeficiente de similitud de Morisita-Horn, utilizando la abundancia a partir de la CPUE (N/100 anzuelos*h) para los grandes pelágicos en el Caribe colombiano. 38 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA PACÍFICO 0,2% La captura total obtenida fue 5.804 kg, correspondiente a 282 individuos identificados dentro de 11 especies agrupadas en siete familias, entre las cuales se destacaron las familias Istiophoridae (43,0 %) y Carcharhinidae (23,8 %) [Figura 4-7 ]. De acuerdo con estudios recientes en el área, Rueda et al. (2014) reportan a la familia Coryphaenidae (dorados) como la más abundante (48,4 %) en 2013, seguida por la familia Carcharhinidae (tiburones) (24,7 %). El análisis en la composición de la captura por zona de muestreo mostró la mayor abundancia en peso y número de individuos en la zona costera, representada principalmente por el pez vela I. platypterus, dorado C. hippurus y tiburón sedoso C. falciformis, los cuales también fueron observados en la zona oceánica con alta abundancia en número de individuos, presentando solo I. platypterus una alta abundancia en peso [Figura 4-8 ]. Rueda et al. (2014) realizaron un muestreo en la zona costera en 2013, reportando las especies C. hippurus y C. falciformis como las más abundantes. La alta variabilidad de la captura en peso y número de individuos, limitó el uso de estadística paramétrica para evaluar diferencias. Por lo tanto, se realizó un análisis no paramétrico al comparar las medianas de la captura en peso mediante el test de Kruskall-Wallis, el cual evidenció que no existen diferencias significativas entre la captura por zona de muestreo (X2=1,07, X2=1,31, p-valor > 0,05) [Figura 4-9 ]. La abundancia relativa o CPUE promedio (± DS) en toda el área de estudio fue 0,90 N/100 anzuelos*h (± 1,03). Las estaciones de pesca que presentaron mayor CPUE en su orden fueron la EP14 (3,70 N/100 anzuelos*h) y EP11 (3,19 N/100 anzuelos*h). Las estaciones con menor CPUE fueron la EP08 (0,16 N/100 anzuelos*h) y EP16 (0,21 N/100 anzuelos*h) [Figura 4-10 ]. Istiophoridae Coryphaenidae Alopiidae 17,3% 43,0% 5.804kg Figura 4-7 Composición porcentual de la captura en peso a nivel de familias durante el crucero de investigación de peces grandes pelágicos en el Pacífico colombiano. El test de Kruskal-Wallis no evidenció diferencias significativas para la CPUE en las zonas de muestreo (X2=1,27, p-valor >0,05; Figura 4-11 ), a pesar de obtener los valores más altos de CPUE para la zona costera, representados principalmente por el tiburón sedoso (C. falciformis), pez vela (I. platypterus), dorado (C. hippurus) y el tiburón látigo (A. pelagicus), siendo esta última la especie con mayor índice de abudancia (CPUE) en la zona intermedia. El análisis de clasificación mediante el coeficiente de similitud de Morisita-Horn evidenció la formación de dos asociaciones de peces partir del umbral de similitud del 40 % aproximadamente: (I) las especies P. glauca (tiburón azul) y K. audax (marlín blanco) y (II) las especies C. falciformis (tiburón sedoso), A. pelagicus (tiburón látigo), C. hippurus (dorado) y I. platypterus (pez vela) [Figura 4-12 ]. Se presentó poca asociatividad (<10 %) de las especies grupo I con las del grupo II [Figura 4-12 ]. PESO (Kg) 500 ZONA INTERMEDIA ZONA OCEÁNICA ZONA DE MUESTREO Scombridae 23,8% 1.000 ZONA COSTERA Sphyrnidae Molidae 1.500 0 Carcharhinidae 15,1% Figura 4-9 Comparación de la captura total en peso (kg) y b) Número de individuos, por zona de muestreo de peces grandes pelágicos en el Pacífico colombiano. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA a ZONA COSTERA 41 Istiophorus platypterus Pez vela 68 Coryphaena hippurus Dorado 65 Carcharhinus falciformis Tiburón sedoso 2 Galeocerdo cuvier Tiburón tigre 6 Alopias pelagicus Tiburón látigo 10 Alopias pelagicus Tiburón látigo 10 Istiophorus platypterus Pez vela 8 Kajikia audax Marlin blanco 3 Makaira nigricans Marlin 16 Coryphaena hippurus Dorado 13 Carcharhinus falciformis Tiburón sedoso 8 Prionace glauca Tiburón azul 2 Makaira nigricans Marlin 5 Kajikia audax Marlin blanco 13 Coryphaena hippurus Dorado 4 Istiophorus platypterus Pez vela 1 Alopias pelagicus Tiburón látigo 3 Carcharhinus falciformis Tiburón sedoso 1 Sphyrna lewini Cachuda 2 Acanthocybium solandri Sierra wahoo 1 Mola mola Pez luna 344 380 1224 3.185 kg N=182 559 678 b ZONA INTERMEDIA 156 477 192 1.603 kg N=60 224 308 247 c ZONA OCEÁNICA 13 13 55 134 40 15 250 1.016 kg N=40 135 214 147 Figura 4-8 Composición de la captura en peso (kg) y número de individuos (N) para las zonas de muestreo: a) costera, b) intermedia y c) oceánica, en el Pacífico colombiano. 39 40 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA CAPTURA (Kg) 1800 CAPTURA (Kg) 1400 4,0 3,70 3,5 3,19 3,0 1200 2,5 1000 2,0 800 1,50 0,88 400 0,25 200 1,5 1,20 600 0,65 0,21 0,36 0,85 0,60 0,30 1,0 0,45 0,25 0,31 0,38 EP09 EP10 0,16 0 0,5 0,0 EP11 EP12 EP13 EP14 EP15 EP16 EP17 EP01 ZONA COSTERA EP02 EP03 EP04 EP05 EP06 EP07 ZONA INTERMEDIA EP08 ZONA OCEÁNICA ESTACIONES POR ZONAS DE MUESTREO Figura 4-10 Comportamiento de la captura (kg) y la captura por unidad de esfuerzo (CPUE: N/100 anzuelos*h) en cada estación (EP) y zona de muestreo en el Pacífico colombiano. CPUE (N/100 anzuelos*h) 3 2 1 0 ZONA COSTERA ZONA INTERMEDIA Figura 4-11 Comparación de la captura por unidad de esfuerzo -CPUE (N/100 anzuelos*h) por zona de muestreo de grandes pelágicos en el Pacífico colombiano. ZONA OCEÁNICA ZONA DE MUESTREO SIMILITUD 0,96 0,84 0,72 0,60 0,48 0,36 0,24 0,12 0,00 Marlin blanco (K. audax) Tiburón azul (P. glauca) I Dorado (C. hippurus) Pez vela (I. platypterus) Figura 4-12 Análisis de clasificación de las especies calculado con el coeficiente de similitud de Morisita-Horn utlizando la abundancia a partir de la CPUE (N/100 anzuelos*h) para los grandes pelágicos en el Pacífico colombiano. Tiburón látigo (A. pelagicus) Tiburón sedoso (C. falciformis) II CPUE (N/100 anzuelos*h) 1600 CPUE (N/100 anzuelos*h) EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 43 DISTRIBUCIÓN ESPACIAL DE LA ABUNDANCIA, ESTRUCTURA DE TALLAS Y ESTIMACIÓN DE BIOMASA 5 Alfredo Rodríguez, Jorge Viaña, José Correa, Alexander Girón y Mario Rueda INTRODUCCIÓN Para un manejo adecuado de los recursos pesqueros es fundamental contar con información relacionada con los patrones de distribución espacial de la abundancia y la estimación de la biomasa (Thurow, 1997; Perry et al., 1999; D´Elia et al., 2009), con el fin de determinar el estado de las poblaciones y fortalecer el diseño de estrategias de manejo operacionales (e.g. vedas espacio-temporales, cuotas y esfuerzo de pesca). Una forma de obtener información rápida y confiable consiste en la evaluación directa mediante tecnologías en hidroacústica complementadas con muestreos de pesca exploratoria, brindando así en corto tiempo un cubrimiento amplio de un área dada y contando además con un tamaño muestral representativo que genere mayor certidumbre en la cuantificación del potencial de recursos pesqueros (Bertrand y Josse, 2000; Simmonds y MacLennan, 2005). En este sentido, este capítulo presenta información sobre la distribución espacial de la abundancia de peces y su estructura de tallas, además de una aproximación a la estimación de biomasa efectiva obtenida por el método hidroacústico para el atún aleta amarilla (Thunnus albacares) en el Caribe y el dorado (Coryphaena hippurus) en el Pacífico colombiano. MÉTODOS ESPECÍFICOS En los cruceros se utilizaron embarcaciones funcionales de pesca como plataformas de investigación, siguiendo un diseño sistemático con grilla de muestreo de amplio cubrimiento espacial. En cada campaña se realizó un barrido hidroacústico mediante una ecosonda SIMRAD ES70 (split beam) con transductor de frecuencia 38 kHz [Figura 5.1 ]. Con base en el a b conocimiento de la distribución vertical de peces grandes pelágicos, el rango de detección acústica fue de entre 3 y 200 m, aunque también se recolectó información a mayor profundidad. La ecosonda fue previamente calibrada de acuerdo con el procedimiento estándar para los sistemas de sondeo vertical (Foote et al., 1987; Simrad, 2003). La calibración consistió en medir Figura 5-1 a) Montaje del transductor en la parte lateral del barco para la detección instantánea de peces. b) Diagrama del sistema de información hidroacústica para visualización de las ecodetecciones (ecograma). Fuente: Tomado y modificado de Manual SIMRAD EK60 (2003). 44 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA las señales de intensidad de blanco o “target strength” (TS) y de ecointegración NASC (Nautical Area-Scattering Coeficient, por sus siglas en inglés) provenientes de un blanco estándar (esfera de cobre de 60 mm de diámetro) de fuerza de blanco conocida y suspendida directamente bajo el eje del haz acústico [Figura 5.2 ]. Este método ajustó la ganancia y la señal de ecointegración, necesarios para igualar las intensidades observadas y esperadas del eco producido. La detección hidroacústica fue completada con lances de pesca exploratoria, con el fin de identificar la proporción de especies, estructura de tallas y peso individual. Los datos colectados por el método hidroacústico fueron procesados mediante un análisis visual de los ecotrazos (ecogramas) integrados en intervalos de 50 m y espaciados cada 1 mn (unidades básicas de muestreo - ubm). Este proceso se realizó utilizando el software Echoview 4.0 especializado en el posproceso de información hidroacústica (Simmonds y MacLennan, 2005; https://www. echoview.com/). En cada ecograma se determinaron los ecos de especies objetivo (e.g. pelágicos) y se eliminaron aquellos ecos asociados a otras fuentes, tales como capas de plancton, ruido de la embarcación, fondos levantados, ruido ambiental, burbujas de aire, entre otros (Simmonds et al., 1992; Simmonds y MacLennan, 2005). De esta forma, se obtuvieron valores del coeficiente de retrodispersión por unidad de área (NASC) como medida de densidad acústica. Una vez obtenido el NASC, se procedió con la asignación de la proporción de especies capturadas en los lances de pesca. Para cada especie se determinó el sigma-ton σTon(i)(x,y), el cual define la relación longitud y peso promedio de los peces, reflejando la estructura de tamaños del área donde se realizó el lance de pesca: σTon(i)(x,y) = σi(x,y) Wi(x,y) donde Wi(x,y), es el peso promedio (en toneladas) de la especie i del lance de identificación en la localidad (x,y), y σ(x,y) es la sección promedio transversal de retrodispersión sonora (en m2) de la especie i: σi(x,y) = donde fi es la frecuencia de clases de tamaños j a partir de los lances de pesca y TS es la fuerza de blanco de tamaño de clase Li [Tabla 5.1 ]. Tabla 5-1 Función de fuerza de blanco o Target Streght (TS) del atún aleta amarilla (Thunnus albacares) y dorado (Coryphaena hippurus). Especie Función TS Fuente Thunnus albacares (Atún aleta amarilla) TS=25,26*log(L)-80,62 Bertrand y Josse, 2000 Coryphaena hippurus (Dorado) TS=20*log(L)-67,4 (peces fisoclistos) Simmonds y McLennan, 2005 La densidad acústica o NASC se transformó en una medida densidad biológica (t/mn2) de la especie i en la localidad (x,y), zi(x,y), al dividirlo entre la sección transversal de retrodispersión sonora normalizada por unidad de peso (sTon(i)) en m2/t: zi (x,y) = NASC i (x,y) σTon(i) (x,y) La biomasa de la especie i se calculó a partir del producto entre la densidad media zi (x,y) (t/mn2) y el área efectiva ocupada por la especie Aei (mn2). La densidad media de cada especie fue estimada mediante un análisis geostadístico (Rivoirard et al., 2000): Bi=zi(x,y)*Aei Para determinar la distribución espacial de la densidad o abundancia relativa, se realizó un análisis geostadístico, el cual permite cuantificar la continuidad espacial de una variable regionalizada (autocorrelación espacial) a través de la construcción de una variograma. El variograma es la función que describe la variabilidad espacial a partir de qué tan parecidos son los puntos en el espacio a medida que estos se encuentran más alejados y las varianzas en función de la distancia que separa a las muestras (Rivoirard et al., 2000; Gallardo, 2006): γ(h) = Figura 5-2 Esquema de calibración de la ecosonda mediante una esfera con fuerza de blanco conocida y suspendida bajo el eje del haz acústico del transductor. Fuente: Tomado y modificado de Manual SIMRAD EK60 (2003). 1 2N(h) N(h) i=1 [ z(x i)-z(x i+h) ] 2 donde γ es el variograma en función de la distancia (h, medida en km); N es el número de pares de datos separados por h; Z(xi) es el valor de la variable en la muestra (xi); Z(xi + h), es 45 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA El mejor modelo y parámetros de variograma se escogieron para la construcción de mapas de distribución espacial mediante la técnica de interpolación Kriging (Rivoirard et al., 2000). El análisis espacial se realizó en la plataforma computacional R-Project con la librería geoR (R Development Core Team, 2015). La distribución RESULTADOS Y DISCUSIÓN 0,06 Varianza o Sill parcial (C) 0,03 0,02 15 30 45 60 75 90 Distancia (km) Figura 5-3 Representación gráfica de la función de variograma. espacial de las tallas en las especies más representativas de la captura se determinó mediante cartografía temática a través de un sistema de información geográfica –SIG–. Semivarianza 40 35 30 25 20 15 10 5 0 30 60 90 120 150 180 60 75 90 Distancia (km) b 60 50 40 30 20 10 0 15 30 45 Distancia (km) Figura 5-4 Variograma omnidireccional de la densidad de a) atún aleta amarilla (Thunnus albacares) y b) dorado (Coryphaena hippurus), en el Caribe continental colombiano. Tabla 5-2 Resumen del análisis geostadístico para la estimación de biomasa del atún aleta amarilla (Thunnus albacares) y el dorado (Coryphaena hippurus) en el Caribe continental colombiano. Co: nugget o efecto pepita; C+Co: Sill o varianza; Ao: Rango o distancia de autocorrelación (km); CME: Cuadrado medio del error. Especie Efecto pepita o Nugget (Co) Rango (Ao) 0,00 Semivarianza La distribución espacial de la densidad del atún aleta amarilla (Thunnus albacares) fue explicada por un variograma isotrópico ajustado con un modelo esférico [Figura 5-4a ]. Lo anterior implicó que no hubo efecto direccional de la distribución de la abundancia o en otras palabras la especie se distribuyó de igual forma en todas las direcciones. El rango de autocorrelación fue 69,6 km y representó el tamaño umbral de los parches de atún con alta densidad [Tabla 5-2 ]. El modelo ajustado presentó una estructura no resuelta por el diseño de muestreo equivalente al 5,2 % de la varianza total [Tabla 5-2 ]. El análisis de distribución espacial mostró un parche con densidad alta en la zona sur frente al golfo de Morrosquillo (>171<334 t/mn2) y otro parche de menor tamaño y densidad intermedia (>44<85 t/mn2) frente a Barranquilla en la zona centro [Figura 5-2 ]. La biomasa instantánea estimada fue 9.200 t, con una densidad media igual a 7,44 t/mn2 y un área efectiva de ocurrencia del recurso de 1.236 mn2 [Tabla 5-2 ]. 0,04 0,01 a CARIBE Sill (Co+C) 0,05 Semivarianza el valor de la variable separada por xi y h. Se ajustaron modelos (e.g. gaussiano, esférico, exponencial) al variograma experimental para escoger el mejor modelo mediante la validación cruzada que produjo el menor valor del cuadrado medio del error (Cressie, 1993; Bez, 2002). Los parámetros de variograma estimados fueron: i) efecto nugget (Co), que indica las variaciones que ocurren a una escala espacial no explicada por el modelo; ii) sill parcial (C), es la varianza explicada por el modelo; iii) sill (Co+C), define el valor máximo de semivarianza de la variable dependiente y, iv) el rango (Ao), es la distancia (en km) hasta la cual ocurre la autocorrelación espacial [Figura 5-3 ]. Parámetros de variograma Área efectiva Densidad Biomasa (t) (mn2) media (t/mn2) Varianza Modelo Co C+Co Ao CME Thunnus albacares (atún aleta amarilla) Esférico 2 36,6 69,6 0,01 1.236 7,44 9.200 201 Coryphaena hippurus (dorado) Esférico 5 5,6 64,5 0,05 530 3,21 1.700 66 46 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Figura 5-5 Distribución espacial de la densidad (t/ mn2) del atún aleta amarilla (Thunnus albacares) en el Caribe continental colombiano. La distribución espacial de la densidad del dorado (Coryphaena hippurus) también fue explicada por un variograma omnidireccional isotrópico ajustado con un modelo esférico [Figura 5-4b ]. El rango de autocorrelación fue 64,5 km y representó el tamaño de los parches con alta densidad [Tabla 5-2 ]. El modelo ajustado presentó una estructura no resuelta por el diseño de muestreo equivalente al 8,8 % de la varianza total [Tabla 5-2 ]. El dorado presentó parches con densidad alta (>61<113 t/mn2), principalmente en la zona centro frente a Barranquilla [Figura 5-6 ]. La biomasa estimada fue 1.700 t, con una densidad media igual a 3,21 t/mn2 y un área efectiva de 530 mn2 [Tabla 5-2 ]. Figura 5-6 Distribución espacial de la densidad (t/mn2) del dorado (Coryphaena hippurus) en el Caribe continental colombiano. 47 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA El 90 % de los individuos del atún aleta amarilla fueron adultos, los cuales se encontraron agregados principalmente en la zona norte, exhibiendo tallas entre 164 y 191 cm de longitud total (LT), mientras que la mayoría de individuos con menores tallas se localizaron en la zona centro y sur [Figura 5-7 ]. Por el contrario, el dorado exhibió los individuos más grandes principalmente hacia las zonas centro y sur con tallas superiores entre 136 y 152 cm de LT [Figura 5-8 ]. Figura 5-7 Distribución espacial de las tallas (cm de longitud total) del atún aleta amarilla (Thunnus albacares) en el Caribe continental colombiano. Figura 5-8 Distribución espacial de las tallas (cm de longitud total) del dorado (Coryphaena hippurus) en el Caribe continental colombiano. 48 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA PACÍFICO 150 125 Semivarianza La distribución espacial de la densidad del dorado (C. hippurus) fue explicada por un variograma isotrópico ajustado con un modelo esférico [Figura 5-9 ]. El rango de autocorrelación fue 41,7 km y representó el tamaño del parche con alta densidad [Tabla 5-3 ]. El modelo ajustado presentó una estructura no resuelta por el diseño de muestreo equivalente al 9,0 % de la varianza total [Tabla 5-3 ]. El análisis espacial mostró solo un parche con densidad alta en la zona costera frente a Buenaventura (>66<125 t/mn2), mientras que en el resto del área evaluada las densidades fueron bajas (<16 t/mn2) [Figura 5-10 ]. La biomasa estimada fue 3.900 t, con una densidad media igual a 3,6 t/mn2 y un área efectiva de 1.087 mn2 [Tabla 5-3 ]. 100 75 50 25 0 20 Coryphaena hippurus (dorado) 80 100 120 Figura 5-9 Variograma omnidireccional de la densidad del dorado (Coryphaena hippurus) en el Pacífico colombiano. Tabla 5-3 Resumen del análisis geostadístico de la densidad del dorado (Coryphaena hippurus) en el Pacífico colombiano. Co: nugget o efecto pepita; C+Co: Sill o varianza; Ao: Rango o distancia de autocorrelación (km); CME: Cuadrado medio del error. Parámetros de variograma Modelo Co C+Co Ao CME Área efectiva (mn2) Esférico 10,61 107,7 41,7 0,12 1.087 Figura 5-10 Distribución espacial de la densidad (t/mn2) del dorado (Coryphaena hippurus) en el Pacífico colombiano. 60 Distancia (km) Respecto a la distribución espacial de las tallas de dorado, se encontró a los individuos más grandes al norte de la zona costera e intermedia, con tallas entre 127 y 145 cm de LT; mientras que se observó una tendencia de individuos de menores tallas (>103<117 cm de LT) hacia la zona oceánica [Figura 5-11 ]. Especie 40 Densidad media (t/ mn2) Biomasa (t) Varianza 3,59 3.900 78 Figura 5-11 Distribución espacial de las tallas (cm de longitud total) del dorado (Coryphaena hippurus) en el Pacífico colombiano. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 6 51 ASPECTOS REPRODUCTIVOS DE RECURSOS GRANDES PELÁGICOS Diana Bustos-Montes, Sarith Salas-Castro, Judith Liliana Prieto y Eliana Cárdenas INTRODUCCIÓN El conocimiento de la biología reproductiva de las especies es esencial para el análisis de la dinámica poblacional, la evaluación de poblaciones y el diseño de estrategias para el manejo pesquero (Saborido, 2004). En el caso específico de los recursos pesqueros de Colombia, en particular para la especies que habitan en aguas oceánicas, la información en cuanto a los aspectos reproductivos es limitada (Rueda et al., 2014; Grijalba et al., 2012). De acuerdo con esto, el presente capítulo aborda aspectos reproductivos (estado y distribución espacial) de las principales especies capturadas durante cruceros de investigación de recursos grandes pelágicos en el Caribe y Pacífico colombiano en los años 2013 y 2014, a través de la determinación de los estados de madurez con énfasis en el dorado (Coryphaena hippurus) y el atún aleta amarilla (Thunnus albacares), información relevante para inferir el estado de las poblaciones evaluadas. MÉTODOS ESPECÍFICOS Con base en muestras de gónadas recolectadas y procesadas en los cruceros de investigación realizados en 2014, complementadas con muestras de los cruceros llevados a cabo en 2013 para la misma área de estudio y similar periodo en el Caribe y Pacífico, con base en análisis microscópicos (histología) se confirmó la determinación del estado de madurez establecida a bordo mediante observaciones macroscópicas. Las especies estudiadas fueron, para el Caribe, el atún aleta amarilla (T. albacares) y el dorado (C. hippurus) en el Pacífico, teniendo en cuenta que fueron las más abundantes en la captura. Para el análisis histológico, las gónadas fueron fijadas en formalina al 4 % y preservadas en alcohol al 70 %, se realizó un corte transversal poniendo la muestra en casetes plásticos para proceder con la deshidratación, pasando por una serie sucesiva de alcoholes así: 70 % (2 h), 90% (2 h), 96 % (dos recambios cada uno de 1 h), 100 % (tres recambios, uno de 1 h y 2 de 2 h), xileno (tres recambios, uno de 1 h y 2 de 1:30 h), y finalmente parafina líquida (a 56°C dos recambios de 2 y 3 h respectivamente). Este procedimiento se efectuó en un procesador de tejido automático histokinette marca Shandon Citadel 2000 ® (Gómez-León, 2005) [Figura 6-1 ]. Posteriormente, se hicieron cortes de 5 μm de espesor utilizando un micrótomo Microm HM 325. Los cortes se pusieron en agua tibia al baño maría entre 1 y 5 minutos y después en un portaobjeto. El tejido adherido al portaobjetos se tiñó con la técnica de hematoxilina-eosina (Gómez-León, 2005; Figura 6-2 ; Tabla 6-1 ). Por último, se hizo el montaje colocando el cubreobjetos con pegamento (Permounth). Los cortes fueron observados en un microscopio óptico y se realizó un registro fotográfico de las gónadas en cada uno de los estadios de madurez encontrados. Además se hicieron observaciones y mediciones de microestructuras para establecer el sexo y estado de maduración sexual predominantes para cada organismo [Figura 6-3 ]. Se presenta la descripción histológica gonadal de los estados de madurez encontrados para atún aleta amarilla y el dorado, así como el porcentaje de coincidencia entre las observaciones macroscópicas y microscópicas. Además se muestran fotografías que ilustran los diferentes estados de desarrollo a fin de que puedan ser usados como guía en futuros estudios reproductivos. Por último, con la información generada en el crucero de prospección de 2014, se realizó un análisis de distribución espacial de la madurez mediante un sistema de información geográfica –SIG–, con el fin de identificar potenciales áreas de interés para el ciclo reproductivo de estas especies. 52 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA a b c d Figura 6-1 Proceso metodológico para la obtención de cortes histológicos de una fracción de gónada. a) ubicación de las gónadas en bandeja plástica para su posterior corte, b) herramientas de trabajo para el procedimiento (bisturí, pinza y rejilla), c) corte longitudinal de la parte media de la gónada seleccionada para el análisis histológico, y d) gónada recolectada de gran tamaño. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 53 a b d c Figura 6-2 Proceso metodológico para la obtención de cortes histológicos de una fracción de gónada. a) realización de cubos de parafina con la gónada inmersa, b) organización de los cubos para su posterior corte, c) micrótomo utilizado para los cortes, y d) corte de los cubos de parafina mediante el micrótomo para la obtención de las placas histológicas. 54 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Tabla 6-1 Tiempos para la tinción con la técnica de hematoxilina-eosina (Gómez-León, 2005). Reactivo Tiempo Xileno (35 o 36°C) 2 recambios por 5 minutos Etanol 100% 5 minutos Etanol 100% 5 minutos Etanol 96% 5 minutos Etanol 10% 5 minutos Agua destilada 5 minutos Hematoxilina 8 minutos Agua destilada 2 minutos Alcohol ácido 5 minutos Agua corriente 5 minutos Agua destilada 3 minutos Etanol 96% 3 minutos Eosina 2 minutos Etanol 96% 25 inmersiones Etanol 96% 25 inmersiones Etanol 96% 3 minutos Etanol 100% 1 minuto 30 segundos Etanol 100% 3 minutos Xileno 5 minutos Xileno 5 minutos Figura 6-3 Ejemplos de placas histológicas obtenidas, para su posterior análisis descriptivo y comparativo. RESULTADOS Y DISCUSIÓN CARIBE En la Tabla 6-2 se presentan los estados de madurez macroscópicos para las diferentes especies de peces capturados durante los cruceros de investigación en el Caribe desarrollados en mayo de 2013 y mayo-junio de 2014. Específicamente para el atún aleta amarilla, T. albacares, se capturaron 115 individuos, 55 en 2013 y 60 en 2014, con prevalencia en ambos cruceros peces maduros (estados IV y V) y capturas similares de hembras y machos. Como afirman Rueda et al. (2014a), los estudios de biología reproductiva de T. albacares, se han enfocado en el océano Pacífico e Índico (Wild 1986; Schaefer, 1998; Lehodey y Leroy, 1999; Itano, 2000; Kikkawa y Cushing, 2002; Sun et al., 2003; Sun et al. 2005; Hoyle et al. 2009; Prathibha y Mohan, 2009; Prathibha et al. 2012 y Zudaire et al., 2013) y existen muy pocos estudios para el Atlántico (Rossignol, 1968; Albaret, 1977, Cayré et al., 1993 y Lessa y Duarte-Neto, 2004). Rossignol (1968) estimó para el Atlántico oriental una talla mínima de madurez entre 50 - 60 cm de longitud total (LT), mientras que Albaret (1977), para la misma región, afirma que a los 108 cm de longitud horquilla (LH) todas las hembras han madurado. Posteriormente, Cayré et al. (1993), calcularon una talla de madurez (L50) entre 97 y 101 cm LH y Lessa y Duarte-Neto (2004) esti- maron la edad de madurez al 50 % en 3,4 años. Respecto a la madurez en machos no hay reportes específicos para el Atlántico, y varios autores han optado por asumir que es similar a la de las hembras. Recientemente, Brown-Peterson et al. (2014), analizaron capturas deportivas de atún aleta amarilla en el Golfo de México entre 2000 y 2011, concluyendo que la temporada reproductiva ocurre entre mayo y septiembre, que la especie es de desarrollo asincrónico, fecundidad indeterminada, desovador por lotes cada 1,16 días aproximadamente y que alcanza la talla mínima de madurez a los 68,5 cm LH. A la fecha se desconoce la variación espacial, temporal y batimétrica de los aspectos reproductivos de T. albacares en el Caribe, lo que representa un gran vacío de información base para el conocimiento de la dinámica poblacional de la especie y un adecuado manejo pesquero. Por lo anterior tiene relevancia el presente estudio ya que brinda información más precisa de aspectos reproductivos de este atún en territorio de Colombia, lo que puede servir como insumo para iniciativas de manejo y conservación que puedan adoptarse en el país. 55 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Tabla 6-2 Sexo y estados de madurez macroscópicos de peces capturados en los cruceros de investigación de grandes pelágicos realizados durante el 2013 y 2014 en el Caribe colombiano. Crucero Familia Especie Sexo Estado de madurez I II III IV V Total Coryphaenidae Coryphaena hippurus Dorado Hembras – – 14 2 – 16 Machos – 6 1 1 – 8 Sphyraenidae Sphyraena barracuda Barracuda Hembras – 1 – 1 – 2 Machos – – 1 3 – 4 Acanthocybium solandri Sierra wahoo Hembras – – – 2 – 2 Machos 3 – 1 – – 4 Machos – 1 – – – 1 Thunnus albacares Atún aleta amarilla Hembras 1 4 14 2 2 23 Machos 1 5 11 14 1 32 Istiophorus albicans Pez vela Hembras – – 1 2 Machos – – 1 – 1 2 Kajikia albida Marlín blanco Hembras – – 1 – – 1 Machos – – 1 – – 1 Makaira nigricans Marlín Hembras 2 – – 1 – 3 Machos – – – 2 – 2 Machos – 1 – – – 1 Scombridae Crucero 2013 Istiophoridae Thunnus alalunga Albacora Tetrapturus pfluegeri Marlín picudo 3 Coryphaenidae Coryphaena hippurus Dorado Hembras – 1 2 25 1 29 Machos 5 10 4 10 – 29 Sphyraenidae Sphyraena barracuda Barracuda Hembras 1 3 2 4 – 10 Machos – – 1 1 – 2 Acanthocybium solandri Sierra wahoo Hembras – 1 2 6 – 9 Machos 1 – 1 – – 2 Hembras 1 – – – – 1 Hembras 4 3 1 23 – 31 Machos 6 3 1 19 – 29 Hembras 6 4 3 2 – 15 Machos 3 – – – – 3 Indeterminados – – – – – 4 Hembras 9 3 5 4 – 21 Machos 3 2 6 1 12 Machos 1 – – – – 1 Hembras 1 – 4 1 – 6 Machos – – 1 1 – 2 Scombridae Katsuwonus pelamis Barrilete Thunnus albacares Atún aleta amarilla Crucero 2014 Xiphiidae Xiphias gladius Pez espada Istiophorus albicans Pez vela Istiophoridae Kajikia albida Marlín blanco Makaira nigricans Marlín La Figura 6-4 muestra la frecuencia de tallas de los atunes aleta amarilla capturados durante ambos cruceros, en el caso de las hembras, en su mayoría fueron hembras en estados de madurez avanzada. Tomando como referencia la talla de madurez propuesta por Cayré et. al. (1993) y asumiendo que se alcanza alrededor de la misma talla/edad para ambos sexos. Teniendo en cuenta lo anterior, puede afirmarse que cerca del 85,1 % de los ejemplares capturados durante ambos cruceros fueron individuos maduros. Considerando que el arte de pesca empleado y las estaciones muestreadas corresponden a los habitualmente usados y visitados por la flota industrial de atún con palangre en el Caribe colombiano, podría inferirse que la selectividad de pesca de la flota es buena en el sentido que una alta proporción de la captura son individuos adultos. 56 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 18 16 TMM=97-101 cm LH (Cayré et ál., 1993) Hembras 14 Estado I 12 10 Estado II 8 Estado III Frecuencia absoluta 6 4 Estado IV 2 Estado V 16 14 40 50 60 70 80 90 100 110 120 130 140 150 160 170 Figura 6-4 Histograma de frecuencia de tallas de los ejemplares de Thunnus albacares (atún aleta amarilla) capturados durante los cruceros de grandes pelágicos en el Caribe colombiano (2013 y 2014). La TMM indicada fue estimada por Cayré et al. (1993). Longitud horquilla (cm) Machos 12 10 8 6 4 2 En cuanto a la distribución espacial de los estados de madurez gonadal durante el muestreo realizado entre mayo y junio de 2014, se encontró la mayoría de los individuos maduros (estado III a V) de T. albacares agregados frente de Santa Marta y frente al golfo de Morrosquillo, mientras que la ocurrencia de individuos inmaduros fue muy baja [Figura 6-5 y Figura 6-6 ]. Figura 6-5 Distribución espacial de los individuos maduros del atún aleta amarilla (T. albacares) en el Caribe colombiano durante la campaña realizada entre mayo-junio de 2014. Es común que se presenten errores en la determinación de sexo y estado de madurez a nivel macroscópico, razón por la cual las escalas son validadas con técnicas histológicas. Para este caso, se validaron 89 observaciones gonadales de T. albacares, encontrando un porcentaje de coincidencia de 41,5 % (N=37), mientras que la diferencia se ve representada en un 58,4 % (N=52). Asimismo, solo en tres individuos se presentó confusión en la determinación del sexo, mientras los restantes hacen refe- rencia al mismo sexo pero en diferentes estados. Se observó que la discrepancia más reincidente se presentó en las determinadas a nivel macroscópico como HII (hembras estado II, es decir inmaduras), que luego fueron clasificadas con el análisis histológico en estado madurando (HIII; 8,57 %; Tabla 6-3 ) y los machos a los que se les asignó el primer estado de inmadurez (MI) y que por técnicas histológicas se observó que ya habían madurado (MIII y MIV). EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 57 Figura 6-6 Distribución espacial de los individuos inmaduros del atún aleta amarilla (T. albacares) en el Caribe colombiano durante la campaña realizada entre mayo-junio de 2014. Diferencias N %D HIV-MIV 1 1,92 MI-HI 1 1,92 MI-HII 1 1,92 HI-HII 1 1,92 HI-HIII 3 5,77 HII-HI 1 1,92 HII-HIII 8 15,38 HII-HV 1 1,92 HIII-HIV 1 1,92 HIV-HIII 1 1,92 HIV-HV 4 7,69 H?-HII 1 1,92 H?-HIV 1 1,92 MI-MII 4 7,69 MI-MIII 5 9,62 MI-MIV 4 7,69 MII-MIII 3 5,77 MII-MIV 4 7,69 MIII-MIV 4 7,69 MIV-MII 1 1,92 MIV-MIII 1 1,92 MIV-MV 1 1,92 Total 52 100,00 Tabla 6-3 Porcentaje de diferencia relacionando el estado definido a nivel macroscópico y el determinado en el análisis histológico para Thunnus albacares (atún aleta amarilla). N número de individuos que presentan diferencias en la clasificación, %D porcentaje que representa cada diferencia. HI hembra uno, HII hembra dos, HIII hembra tres, HIV hembra cuatro, HV hembra cinco, H? hembra sin identificar estado de madurez, MI macho uno, MII macho dos, MIII macho tres, MIV macho cuatro, MIV macho cinco. Nota: Las posibles confusiones siempre se efectúan ubicando inicialmente el sexo y estado de madurez macroscópico. El registro fotográfico gonadal de campo y la verificación microestructural de 89 cortes histológicos permitieron proponer una escala de madurez gonadal macroscópica y microscópica ilustrada para T. albacares en el Caribe colombiano que se presenta en la Tabla 6-4 . 58 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Tabla 6-4 Escala de madurez gonadal macroscópica y microscópica propuesta para Thunnus albacares (atún aleta amarilla). Se incluye la descripción macro y microestructural de los estados de madurez gonadal. HEMBRA ESTADO I • Inmaduro DESCRIPCIÓN MACROSCÓPICA Descripción microscópica Ovarios cerca de 1/3 de la longitud de la cavidad abdominal, de color rosáceo translúcido. Huevos invisibles a simple vista. Los oocitos se encuentran organizados en forma de lamelas, de manera paralela, en estadio nucléolo cromatina inicial, intermedia y tardía. En el estadio intermedio, las células gonadales presentan un núcleo circular y de gran tamaño, en comparación de los nucléolos, los cuales aumentan su volumen a medida que se avanza en los estadios de desarrollo. También se hacen visibles vacuolas grandes en el citoplasma. Vista macroscópica Vista microscópica No disponible a a Microfotografía (100 X) de ovario de Atún aleta amarilla Thunnus albacares en estado I (Inmaduro). OI oocito inmaduro, N Núcleo, Cp citoplasma HEMBRA ESTADO II • Inmaduro DESCRIPCIÓN MACROSCÓPICA Descripción microscópica Ovario cerca de 1/3 de la longitud de la cavidad abdominal, de color rosáceo translúcido, huevos invisibles a simple vista. Los oocitos más grandes se encuentran en posición central, mientras que los más pequeños se ubican hacia la periferia de la célula. En este estado se pueden diferenciar los estadios perinuclear inicial, intermedio y tardío. En el inicial, se distinguen oocitos con núcleos grandes y nucléolos que han aumentado su volumen, estos últimos se van ubicando en la periferia del núcleo, las células carecen de vacuolas. En el estadio intermedio los nucléolos se organizan en fila en la periferia del núcleo, mientras que en el estadio tardío el núcleo se hace más grande y circular, con nucléolos en su periferia y el citoplasma aumentado en tamaño. Vista macroscópica Vista microscópica a EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 59 b a Microfotografía (10 X) de ovario de Atún aleta amarilla Thunnus albacares en estado II (Inmaduro). OI Oocito inmaduro, Lm Lamela b Microfotografía (40 X) de ovario de Atún aleta amarilla Thunnus albacares en estado II (Inmaduro) N Núcleo, Cp citoplasma HEMBRA ESTADO III • Madurando Descripción macroscópica Descripción microscópica Ovarios cerca de 2/3 de la longitud de la cavidad abdominal. Ovarios de color rosáceo amarillo con aspectos granular, no hay huevos transparentes o translúcidos visibles. Se observan lamelas en posición radial. En general el ovario presenta un tabique de tejido muscular liso que atraviesa la gónada. Las células están provistas de un núcleo grande y circular, los nucléolos se ubican en la periferia, siendo estos pocos visibles, el citoplasma es poco basófilo y contiene vesículas o vacuolas grandes y circulares. Los oocitos nucléolo cromatina, perinuclear y alveolo cortical se distribuyen en los extremos de la célula. Vista macroscópica Vista microscópica a b a Microfotografía (100 X) de oocito de Atún aleta amarilla Thunnus albacares en estado III (madurando). OI oocito inmaduro, AI alveolos corticales, OAC , oocito en alveolos corticales b Microfotografía (40 X) de oocito de Atún aleta amarilla Thunnus albacares en estado III (madurando) N Núcleo, Cp citoplasma, AC alveolos corticales, Nu nucléolo 60 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA HEMBRA ESTADO IV • Maduro Descripción macroscópica Descripción microscópica Los ovarios ocupan 2/3 de toda la longitud de la cavidad abdominal, son de color naranja con vasos sanguíneos superficiales visibles, contienen grandes huevos maduros. El ovario alcanza su máximo desarrollo, no se logran diferenciar las lamelas, los oocitos se encuentran muy cercanos unos con otros y se distribuyen por toda la gónada. Se diferencian todos los estadios (nucléolo cromatina, perinucleolar, alveolo cortical y vitelogénico) y se pueden notar oocitos inmaduros tanto en el centro como en la periferia de la lamela. Vista macroscópica Vista microscópica a a Microfotografía (40 X) de ovario de Atún aleta amarilla Thunnus albacares en estado IV (maduro). Zr Zona radiada GV granulos de vitelo, Nu núcleolo, N núcleo. HEMBRA ESTADO V • Desovando Descripción macroscópica Descripción microscópica Los ovarios ocupan entre 1/2 y 2/3 de El ovario se muestra como una estructura laxa, con grandes espacios entre oocitos, toda la longitud de la cavidad abdominal, se observan muchos de ellos hidratados (algunos atrésicos) y solo unos pocos en presentan grandes oocitos transparentes. estados inmaduros y maduros. Algunos puedes ya haber desovado y se observarían paredes flojas, oocitos opacos, maduros, en desintegración, oscurecidos o translúcidos. Vista macroscópica No disponible Vista microscópica a a Microfotografía (10 X) de ovario de Atún aleta amarilla Thunnus albacares en estado V (Desovando). OH oocito hidratado, Gl gota lipidica 61 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA MACHO ESTADO I • Inmaduro Descripción macroscópica Descripción microscópica Testículos cerca de 1/3 de la longitud de Se pueden observar cistos con espermatogonias, espermatocitos primarios y secunla cavidad abdominal, de color blancuzco. darios en menor cantidad. MACHO ESTADO II • Inmaduro Descripción macroscópica Descripción microscópica Testículos cerca de la 1/2 de la longitud Se observan espermatogonias, espermatocitos primarios y secundarios, espermade la cavidad abdominal. De color blan- tides y muy pocos espermatozoides. Los espermatocitos secundarios se ubican en la cuzco a crema. periferia de los cistos. Vista macroscópica Vista microscópica a a Microfotografía (40 X) de testículo de Atún aleta amarilla Thunnus albacares en estado II (Inmaduro). Ecg Espermatogonia, Ecs espermatocito secundario, Ez espermatozoide, Ct cisto MACHO ESTADO III • Madurando Descripción macroscópica Descripción microscópica Testículos cerca de 2/3 de la longitud de la cavidad abdominal, de color crema. En el testículo se encuentran cistos con todos los estadios, la espermatide se localiza en la periferia del cisto y se diferencian espermatozoides con la cabeza en forma de coma y la cola eosinófila y larga de color rosa a roja, las lagunas están llenas de espermatozoides. Vista macroscópica Vista microscópica a a Microfotografía (100 X) de testículo de Atún aleta amarilla Thunnus albacares en estado III (Madurando). Ez espermatozoides, Ecp Espermatocitos primario, Ecs espermatocito secundario 62 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA MACHO ESTADO IV • Maduro Descripción macroscópica Descripción microscópica Los testículos ocupan 2/3 de toda la Se diferencia una capa delgada de tejido muscular liso y se presenta una porción longitud de la cavidad abdominal, blan- gruesa donde se encuentra un canal central que agrupa y expulsa los espermatocuzcos a crema y de apariencia blanda. zoides. A los lados se encuentran lagunas y cistos llenos de espermatozoides que se fusionan al canal. Vista macroscópica Vista microscópica a a Microfotografía (40 X) de testículo de Atún aleta amarilla Thunnus albacares en estado IV (Maduro) Ez espermatozoides Ct Cisto, Ecs espermatocito secundario, Ecp Espermatocitos primario MACHO ESTADO V • Desovado Descripción macroscópica Descripción microscópica Testículos cerca de la 1/2 de la longitud de la cavidad abdominal, los cuales se observan sanguinolentos y flácidos. La gónada presenta una capa delgada de tejido muscular liso en comparación con los estados anteriores, no hay presencia de espermatozoides en el canal central o éstos son muy pocos, al igual que los cistos y las lagunas que se encuentran alrededor del canal. Vista macroscópica Vista microscópica No disponible a b a Microfotografía (10 X) de testículo de Atún aleta amarilla Thunnus albacares en estado V (desovado). Zv Zonas vacías producto de la expulsión. b Microfotografía (40 X) de testículo de Atún aleta amarilla Thunnus albacares en estado V (desovando) Cn Canal delgado, Ez espermatozoides, Ecp Espermatocito primario 63 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA PACÍFICO Durante las campañas de grandes pelágicos en el Pacífico colombiano (diciembre de 2013 y noviembre-diciembre de 2014) el dorado C. hippurus representó la especie de mayor frecuencia de captura. La Tabla 6-5 muestra los estados de madurez macroscópicos para las diferentes especies de peces y específicamente para C. hippurus, donde se aprecia que prevalecieron los peces maduros (estados IV y V) y se capturaron muchas más hembras que machos. Los aspectos reproductivos de C. hippurus en el Atlántico han sido estudiados por Oxenford (1999), Beardsley (1967), Bentivoglio (1988), Perez y Sadovy (1991), Perez et al. (1992) y específicamente, para el Pacífico, Solano- Sare et al. (2008) determinaron que en el Pacífico peruano, C. hippurus alcanza la madurez a los 89 cm de LT. Posteriormente, Ortega-García et al. (2011) determinaron en el Pacífico mexicano que el dorado alcanza la madurez entre 80 y 85 cm LH. De acuerdo con la observación microscópica de las gónadas, se encontraron individuos en su mayoría hembras en estado IV (N=28) y solo cinco en estado V, mientras que el estado III fue el más representativo en machos (N=22). Teniendo en cuenta lo anterior y tomando como referencia la TMM reportada por Solano-Sare et al., (2008), se determinó que el 85,8 % de los individuos capturados se encontraron por encima de la TMM [Figura 6-7 ].Considerando que el arte de pesca empleado y las estaciones muestreadas corresponden a los habitualmente usados y visitados por la flota industrial de grandes pelágicos del Pacífico colombiano, podría inferirse que la selectividad de pesca de la flota es adecuada en el sentido que captura individuos adultos, aunque vale la pena profundizar en la desproporción sexual observada. Tabla 6-5 Sexo y estados de madurez macroscópica de peces capturados en la campaña de grandes pelágicos en el Pacífico colombiano. Crucero Familia Carcharhinidae Crucero 2013 Especie Carcharhinus falciformis Tiburón sedoso Sexo Estado de madurez Total I II III IV V H – – – – – 58 M – – – – – 32 H – – – – – 1 M – – – – – 1 Alopiidae Alopias pelagicus Tiburón látigo Dasyatidae Dasyatis violacea Raya pelágica M – – – – – 1 Coryphaena equiselis Dorado H – – – 1 – 1 H – – 16 96 6 118 M – 7 32 11 1 51 H – 1 – – – 1 H – – – 1 – 1 M – – – 1 2 3 H – – 3 1 – 4 M – – 2 – – 2 H – – – 55 1 56 M – 3 9 25 – 37 H – – 1 – – 1 M – – 1 – – 1 H 2 – 3 – – 5 M – 2 1 35 – 38 H 1 4 6 – – 11 M 1 1 – – – 2 H – 2 3 – – 5 M – – – – – – Coryphaenidae Scombridae Istiophoridae Coryphaenidae Scombridae Coryphaena hippurus Dorado Thunnus albacares Atún aleta amarilla Istiophorus platypterus Pez vela Kajikia audax Marlín rayado Coryphaena hippurus Dorado Acanthocybium solandri Sierra wahoo Istiophorus platypterus Pez vela Crucero 2014 Istiophoridae Kajikia audax Marlín rayado Makaira nigricans Marlín azul 64 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA La distribución espacial de la condición reproductiva del dorado mostró una agregación de individuos maduros hacia el norte de la zona costera entre Cabo Corrientes y Bahía Solano, mientras que la ocurrencia de individuos inmaduros fue casi nula. Lo anterior confirma que el norte del Pacífico es un área importante para el ciclo reproductivo de las especies (Rodríguez et al., 2012; Rueda et al., 2014), donde las condiciones oceanográficas con aguas frías y productivas favorecen el desove [Figura 6-8 , Figura 6-9 ]. Figura 6-7 Histograma de frecuencia de tallas de los ejemplares de dorado Coryphaena hippurus capturados durante la campaña de grandes pelágicos en el Pacífico colombiano. La TMM indicada fue estimada por SolanoSare et al. (2008) para el Pacífico peruano. 45 40 TMM= 89 cm LH Hembras 35 30 25 Estado II 20 Estado III Frecuencia absoluta 15 10 Estado IV 5 Estado V 50 30 25 60 70 80 90 100 110 120 130 140 150 160 170 180 Longitud horquilla (cm) Machos 20 15 10 5 Figura 6-8 Distribución espacial de los individuos maduros de dorado (Coryphaena hippurus) en el Pacífico colombiano durante la campaña realizada en 2014. 65 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Figura 6-9 Distribución espacial de los individuos inmaduros de dorado (Coryphaena hippurus) en el Pacífico colombiano durante la campaña realizada en 2014. Se comparó entre la determinación del sexo y estado de madurez a nivel macroscópico y el análisis microscópico, estableciéndose que el porcentaje de coincidencia fue de 67,5 % (N=50), mientras que la diferencia fue representada en 32,5 % (N=24). En dos individuos se presentó confusión en la determinación del sexo, mientras los restantes hacen referencia al mismo sexo pero en diferentes estados. La confusión más frecuente fue MIV determinados a nivel macroscópico, los cuales el análisis histológico definió como MIII con 31,8 % [Tabla 6-6 ]. Diferencias N %D HII-HIII 1 4,55% HIII-HIV 3 13,64% Al igual que en T. albacares, el registro fotográfico gonadal de campo y la verificación microestructural de 74 cortes histológicos del Pacífico colombiano permitieron proponer una escala de madurez gonadal macroscópica y microscópica ilustrada para C. hippurus presente en territorio colombiano que se presenta en la Tabla 6-7 . HIII-HV 1 4,55% HIV-HIII 2 9,09% HIV-HV 3 13,64% MII-MIII 2 9,09% MIII-MIV 1 4,55% MIV-MIII 7 31,82% M?-MIII 2 9,09% Total 22 100,00% Nota: las posibles confusiones siempre se efectúan ubicando inicialmente el sexo y estado de madurez macroscópico. Tabla 6-6 Porcentaje de diferencia relacionando el estado definido a nivel macroscópico y el estado determinado en el análisis histológico. N número de individuos que presentan diferencias en la clasificación, %D porcentaje que representa cada diferencia. HII hembra dos, HIII hembra tres, HIV hembra cuatro, HV hembra cinco, M? macho sin identificar estado de madurez, MII macho dos, MIII macho tres, MIV macho cuatro, MIV macho cinco. 66 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Tabla 6-7 Escala de madurez gonadal macroscópica y microscópica propuesta para el dorado Coryphaena hippurus, se incluye la descripción macro y microestructural de los estados de madurez gonadal. HEMBRA ESTADO III • Madurando Descripción macroscópica Descripción microscópica Los ovarios ocupan cerca de 2/3 de la cavidad abdominal, de color rosáceo o amarillo, de aspecto granular, con poca irrigación sanguínea y huevos poco visibles. En este estado se observan gránulos de vitelos localizados en la zona cortical del citoplasma, indicando el inicio de la vitelogénesis, siendo de esta forma los oocitos más grandes con diámetros cercanos a 0,1 mm. Además se establece la presencia de gotas lipídicas y los nucléolos migran a la periferia del núcleo. La forma general de los oocitos en estado de vitelogénesis es irregular Sin embargo en esta etapa se presentan oocitos en estado de alveolo cortical e incluso primario pero son menos frecuentes. Vista macroscópica Vista microscópica a b c a Microfotografía (4X) de ovario de dorado Coryphaena hippurus en estado III (Madurando), Gv gránulos de vitelo, Oi oocito inmaduro, N núcleo, Nu nucléolos. b Microfotografía (10X) de ovario de dorado Coryphaena hippurus en estado III (madurando). Cp citoplasma y Gl gotas lipídicas, N núcleo, Nu nucléolos. c Microfotografía (40X) de ovario de dorado Coryphaena hippurus en estado III (madurando). Ov oocito en vitelogénesis Ot oocito en crecimiento terciario, Os oocito en crecimiento secundario, Gv gránulos de vitelo. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 67 HEMBRA ESTADO IV • Maduro Descripción macroscópica Descripción microscópica Los ovarios ocupan cerca de 2/3 de la cavidad abdominal, son de color rosáceo, amarillo o naranja de aspecto granular con huevos maduros y transparentes, se observan con vasos sanguíneos superficiales visibles. En este estado la principal diferencia con el anterior (HIII) es el tamaño de los oocitos, teniendo en cuenta que se da inicio al proceso de maduración. De acuerdo con esto, el diámetro es de aproximadamente 0,2 mm. Por otra parte se observan gotas lipídicas, las cuales finalmente se unen generando una solo gota de gran tamaño. El núcleo comienza a migrar e incluso en ciertos casos no es evidente, observando únicamente el oocito con abundantes gránulos de vitelo. Al igual que el núcleo la vesícula germinal se traslada y por último los oocitos hidratados hacen presencia en menor medida. Vista macroscópica Vista microscópica a b c a Microfotografía (4X) de ovario de dorado Coryphaena hippurus en estado IV (maduro), N núcleo, Nu nucléolos, Gv gránulos de vitelo, Gl gotas lipídicas, Cp citoplasma, Ac Alveolo cortical. b Microfotografía (10X) de ovario de dorado Coryphaena hippurus en estado IV (maduro). Os oocito en crecimiento secundario, Ov oocito en vitelogénesis, Gv gránulos de vitelo, Oi oocito inmaduro. c Microfotografía (40X) de ovario de dorado Coryphaena hippurus en estado IV (maduro), TEPS tejido epitelial plano simple, N núcleo, Nu nucléolos, Ac Alveolo cortical. 68 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA HEMBRA ESTADO V • Desovando Descripción macroscópica Descripción microscópica Ovarios flácidos, contraídos cerca de la mitad de la cavidad abdominal, con restos de huevos opacos y vasos sanguíneos rotos y ocupan cerca de ½ de la longitud de la cavidad abdominal. Se caracteriza por presentar oocitos hidratados en gran proporción, siendo estos grandes y de formas irregulares. No obstante, en dicho caso para los organismos capturados y determinados como HV, se observó en una gónada la presencia de numerosos oocitos en estado de crecimiento primario acompañados por folículos posovulatorios, estructuras propias de especímenes maduros, explicado con la biología reproductiva de la especie, siendo C. hippurus asincrónica. Esta característica permite encontrar oocitos en estados tempranos con oocitos en estado avanzado de maduración en especímenes que han pasado la talla de primera madurez. A nivel general, las células foliculares se rompen y solo se evidencian los oocitos sin estructuras rodeándolos. Vista macroscópica Vista microscópica No disponible a b c A Microfotografía (4X) de ovario de dorado Coryphaena hippurus en estado V (desove), Oi oocito inmaduro, Os oocito en crecimiento secundario, Fp folículo posovulatorio. B Microfotografía (4X) de ovario de dorado Coryphaena hippurus en estado V (desove). Oh oocito hidratado. C Microfotografía (10X) de ovario de dorado Coryphaena hippurus en estado V (desove) Fp folículo posovulatorio, Nu nucléolos, Cp citoplasma, Oi oocito inmaduro. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 69 MACHO ESTADO II • Inmaduro Descripción macroscópica Descripción microscópica Los testículos ocupan cerca de ½ de la longitud de la cavidad abdominal, su coloración es blancuzco a crema. Este estado se caracteriza por la presencia de cistos de gran tamaño cerca del canal central, mientras que hacia los conductos testiculares van reduciendo su tamaño hasta el punto de hacerse menos evidente. En los cistos se encuentran los espermatocitos primarios y hacia el borde de los cistos los espermatocitos secundarios. Vista macroscópica Vista microscópica a b a Microfotografía (4 X) de testículo de dorado Coryphaena hippurus en estado II (inmaduro). Ct cisto, Cnt canal central. b Microfotografía (40X) de testículo de dorado Coryphaena hippurus en estado II (inmaduro). EcS espermatocito secundario, EcP espermatocito primario. MACHO ESTADO III • Madurando Descripción macroscópica Descripción microscópica Los testículos ocupan aproximadamente cerca de 2/3 de la longitud de la cavidad abdominal, presenta una tonalidad crema. Este estado de maduración intermedia, se caracterizó por la presencia de cistos de gran tamaño que se encuentran en el canal central. En los conductos testiculares, los cistos son alargados y poco frecuentes. De igual forma en dichos conductos el epitelio germinal es discontinuo, mientras en la parte central es continuo. En los cistos se encuentran embebidos los espermatozoides y existen pocos espermatocitos en crecimiento secundario. Vista macroscópica Vista microscópica a b c a Microfotografía (4 X) de testículo de dorado Coryphaena hippurus en estado III (madurando). Ez espermatozoides, Ecs espermatocito secundario. b Microfotografía (10 X) de testículo de dorado Coryphaena hippurus en estado III (madurando). CnT Canal central, Ct cisto c Microfotografía (40X) de testículo de dorado Coryphaena hippurus en estado III (madurando). CnT Canal central, Ez espermatozoides. 70 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA MACHO ESTADO IV • Maduro Descripción macroscópica Descripción microscópica Los testículos ocupan cerca de 1/3 de Los cistos que contienen grandes cantidades de espermatozoides, se hacen más la longitud cavidad abdominal, de color frecuentes en los conductos y comienzan a migrar hacia la pared de la gónada, blancuzco. presentando una forma alargada. En dicha etapa la espermatogénesis se encuentra al máximo, razón por la cual los espermatozoides liberados serán numerosos. Por otro parte rodeando algunas lagunas se encuentra espermatocitos en crecimiento secundario, sin embargo aproximadamente el 90% de la gónada posee agrupaciones de espermatozoides. Vista macroscópica Vista microscópica a b c a Microfotografía (4X) de testículo de dorado Coryphaena hippurus en estado IV (maduro). Ez espermatozoides, Tr tejido reproductor (epitelio germinal), Ecs espermatocito secundario. b Microfotografía (10X) de testículo de dorado Coryphaena hippurus en estado IV (maduro) Ez espermatozoides. c Microfotografía (40X) de testículo de dorado Coryphaena hippurus en estado IV (maduro) Tr tejido reproductor (epitelio germinal), Ct cisto. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 7 73 COMPOSICIÓN, ABUNDANCIA Y DISTRIBUCIÓN ESPACIAL DEL ICTIOPLANCTON Ana Galeano-Chavarría, Gustavo Ramírez y Eugenia Escarria INTRODUCCIÓN El estudio del ictioplancton (huevos y larvas de peces) es un aspecto fundamental para el manejo de recursos pesqueros, debido a que el reclutamiento de peces depende de la supervivencia y disponibilidad de los primeros estadios de desarrollo (Petersen y Warner, 2002), los cuales son muy susceptibles a las variaciones ambientales (Legget y Deblois, 1994; Miller y Kendall, 2009). De esta manera, el estudio de huevos y larvas de peces sirve como un indicador de la producción futura de la población con intereses para la pesca, ya que estos se comportan diferente a los adultos frente a las variaciones en su entorno (Jiménez, 2008). En el ambiente marino diferentes procesos oceanográficos pueden influenciar el desarrollo, distribución y abundancia del ictioplancton y su posterior asociación con otras especies marinas. Entre estos, se encuentran los procesos de circulación de corrientes, frentes térmicos y topografía que incrementan la dispersión tanto vertical como horizontal en diferentes escalas espaciales y temporales (Moser et al., 2002; Silva, 2005; Sánchez et al., 2009; Avendaño et al., 2010). Con base en lo anterior, describir la distribución espacial y la abundancia del ictioplancton y su relación con los factores ambientales, es de gran importancia para entender las fluctuaciones de las poblaciones de peces. Este capítulo muestra los resultados en términos de composición, distribución y abundancia de huevos y larvas de peces en el Caribe y Pacífico colombiano, con el propósito de aportar información que apoye la gestión de los recursos marinos en el país. MÉTODOS ESPECÍFICOS Con base en las muestras de ictioplancton colectadas en los cruceros (para mayor detalle del muestreo ver sección 3), se realizó la estimación del volumen de agua filtrada a través de la red en cada estación de muestreo utilizando la siguiente función: VF(m 3 )=π * m * d 2 donde, VF es el volumen de agua filtrada en m3; π es la constante de 3,1416; m la distancia recorrida por la red (metros) y d el diámetro de la red (metros). Una vez analizadas las muestras, los valores independientes de abundancia de huevos y larvas de peces se estandarizaron a un volumen de 10 m2 de superficie marina teniendo en cuenta la profundidad de arrastre y de acuerdo con las recomendaciones de Smith y Richardson (1979): B C= c * VF donde, C es el número de larvas o huevos por cada 10 m2; c el número de larvas o huevos de la muestra i; B la profundidad máxima de arrastre y VF el volumen de agua filtrada. Con los datos estandarizados por unidad de área se realizó un análisis descriptivo mediante histogramas de frecuencia y tabulaciones para representar la abundancia relativa de huevos y larvas de peces. Para conocer la estructura comunitaria y el ensamblaje de larvas de peces se determinaron índices ecológicos tales como diversidad Shannon-Weiner (H’), el cual representa una medida relativa de la diversidad y es la expresión de dos factores: la riqueza o número de especies y la uniformidad o grado de proporción entre el número de individuos de cada especie en la muestra (Ramírez y Giraldo, 2006): s * 10 H‘= – Pi i=1 log2 Pi 74 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA donde, H’ = medida de diversidad.; s =es el número de especies.; Pi= es la proporción de individuos de la especie i. Todos los índices ecológicos fueron calculados a nivel de familia (Leis, 1993). A partir de una matriz de similtud calculada con el índice de Bray-Curtis, utilizando el algoritmo del grupo promedio y previa transformación de los datos a raíz cuarta para reducir el efecto de las familias y/o especies abundantes (Clarke y Warwick, 2001), se realizó un análisis de varianza mediante permutaciones (PERMANOVA) de una vía (Factor: Zona), para establecer diferencias en la estructura del ensamblaje (Anderson et al., 2008), en las diferentes zonas establecidas a priori. Cuando se encontraron diferencias significtivas, se efectuaron pruebas pareadas entre los grupos. Finalmente, se llevó a cabo un análisis de porcentaje de disimilitud (SIMPER), para identificar aquellas familias que en promedio contribuyeron en mayor medida a la variación entre estas zonas y a la similitud dentro de las mismas. RESULTADOS Y DISCUSIÓN CARIBE Distribución y abundancia de ictioplancton Cerca del 50 % de los huevos obtenidos se encontraron en estado de división o no fecundados, el resto fueron discriminados en tres morfotipos (M): MI (vitelo y/o larva pigmentada), MII (huevo sin pigmento) y MIII (huevo con ornamentación del corión) [Figura 7-1 ]. La distribución espacial de los huevos fue relativamente homogénea en el Caribe, sin embargo, las estaciones que presentaron un mayor valor de densidad estuvieron hacia la zona norte (>90 huevos/10 m2) entre La Guajira y Santa Marta [Tabla 7-1 y Figura 7-2a ], patrón que coincidió con lo expuesto por Rueda et al. (2014). En cuanto a las larvas de peces, estas también presentaron un patrón homegéneo en su distribución espacial con una alta densidad en la zona norte (>178 larvas/10 m2, Tabla 7-1 ; Figura 7-2b ) debido la alta abundancia de la sardina (familia Clupeidae), los cuales son pequeños peláZona Nor te Centro Sur gicos de interés comercial para algunas pesquerías del mundo (Zapata et al., 2011). El patrón de distribución espacial exhibido puede atribuirse a las características oceanográficas de la zona norte, la cual está influenciada por la corriente Caribe, los vientos Alisios y la surgencia estacional entre otros, que contribuyen a una mayor actividad y productividad planctónica (Fajardo, 1979; Andrade, 2000). Tabla 7-1 Valores de densidad (N/10 m2) de huevos y larvas de peces durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. Estación de muestreo Latitud (N) Longitud (W) E01 13°04,836 72°53,776 Huevos Larvas (N/10 m ) (N/10 m2) 103 347 2 E02 12°48,780 73°16,928 12 31 E03 12°52,656 73°26,730 19 40 E04 12°37,607 73°52,158 5 38 E05 12°35,263 73°53,245 2 15 E06 12°19,918 74°23,805 5 14 E07 12°22,475 74°27,700 6 13 E08 12°07,178 74°54,733 7 177 E09 11°42,708 75°16,219 0 120 E10 11°31,211 75°34,124 1 13 E11 11°23,584 75°43,752 12 119 E12 10°53,869 75°53,964 1 76 E13 10°54,697 76°15,133 2 56 E14 10°51,390 76°08,933 10 101 E15 09°34,129 76°34,838 5 93 E16 09°55,098 76°42,303 0 93 E17 10°14,232 76°34,125 6 109 E18 10°24,974 76°28,158 0 43 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA a b c d Figura 7-1 Morfotipos de huevos encontrados durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. a) Morfotipo I; b) Morfotipo II; c) Morfotipo III y d) En estado de división. Fotografías: Ana María Galeano - INVEMAR. a b Figura 7-2 Distribución espacial de la densidad (N/ 10 m2) de a) huevos y b) larvas, durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. 75 76 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Composición de especies y estructura comunitaria Clupeidae 20,03 % Myctophidae 14,97 % Se registraron un total de 42 familias y 87 taxones a nivel de género y especie. Las familias más representativas fueron Clupeidae (sardinas, 20 %); Myctophidae (peces linterna, 15 %); Tetraodontidae (pez globo, 9 %) y Gobiidae (gobios, 7 %). Se destaca la presencia de familias de importancia comercial como Serranidae (chernas, 7 %); Carangidae (jureles y pampanos, 7 %); Scombridae (atunes y sierras, 4,60 %); Paralichthydae (lenguados, 2,87 %); Gerreidae (mojarras, 0,99 %); Lutjanidae (pargos, 0,82 %) [Figura 7-3 ]. Las especies más abundantes y frecuentes en las estaciones de muestreo fueron de la familia Clupeidae (20 %; 300 larvas/10 m2); Diaphus sp. (6,5 %); Sphoeroides spp. (6,5 %); Scombridae sp. (2,4 %) y Serranus spp. (2,2 %), entre otras (Tabla 7-2 y Figura 7-4 ). Otras familias 14,20 % Tetraodontidae 8,98 % Serranidae 7,20 % Gobiidae 6,93 % Carangidae 6,69 % Scombridae 4,60 % Paralichthyidae 2,87 % Gonostomatidae 2,58 % Scaridae 2,23 % Stomiidae 1,95 % Balistidae 1,60 % Bothidae 1,45 % Tabla 7-2 Composición, porcentaje de abundancia (%A) y frecuencia de ocurrencia en porcentaje (%FO) de larvas de peces encontradas durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. NF: identificados a nivel de familia. Taxón Bregmacerotidae 1,41 % Labridae 1,00 % Gerreidae 0,99 % Lutjanidae 0,82 % %A %FO %A %FO Letharchus aliculatus 0,08 5,56 NF 0,08 5,56 Diaphus sp. 6,53 61,11 Lampadena sp. 0,24 5,56 0,20 5,56 Lampadena uraphaos atlantica 0,26 5,56 Lampanyctus sp. 0,79 27,78 0,12 5,56 Myctophum sp. 0,15 5,56 NF 7,01 55,56 20,03 27,78 0,39 5,56 Cyclothone sp. 1,91 55,56 Bregmaceros atlanticus 0,60 27,78 Gonostoma sp. 0,36 11,11 Bregmaceros sp. 0,44 11,11 NF 0,31 11,11 NF 0,36 11,11 0,59 22,22 Lamprogrammus sp. 0,12 5,56 Ophidion sp. 0,26 11,11 0,09 5,56 0,40 22,22 0,05 5,56 0,09 5,56 Ophichthidae Mycthophidae Congridae Heteroconger sp. Derichthyidae Derichthys sp. Clupeidae Clupidae spp. Gonostomatidae Phosichthyidae Vinciguerria sp. 11,11 Astronesthes sp. 0,20 5,56 Bathophilus sp. 0,47 11,11 Eustomias sp. 0,69 22,22 Heterophotus ophisthoma 0,15 5,56 Stomias sp. 0,16 11,11 NF 0,30 11,11 0,12 5,56 Stomiidae Paralepididae Bregmacerotidae Ophidiidae Melanocetidae Melanocetus sp. Holocentridae Myripristis sp. Melamphaidae Melamphaes sp. Scorpaenidae Evermannellidae Evermannella sp. GADIFORMES Macrouridae 0,24 Lestidiops affinis Taxón Figura 7-3 Porcentaje de abundancia de las familias de larvas de peces durante el crucero de investigación de grandes pelágicos en el Caribe continental colombiano. 0,48 11,11 Sebastes sp. 77 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Taxón %A Taxón %FO %A %FO Labridae Serranidae Diplectrum sp. 0,45 5,56 Elegatis bipinnulata 0,17 5,56 Paralabrax sp. 0,15 5,56 Pseudogramma gregoryi 0,47 16,67 Rypticus sp. 0,32 5,56 Serranus sp. 2,17 55,56 Serranus tigrinus 0,15 5,56 NF 3,32 38,89 Priacanthidae Halichoeres sp. 0,49 16,67 Tautogolalorus sp. 0,17 5,56 Xyrichhthys sp. 0,12 5,56 NF 0,22 11,11 Scarus sp. 0,13 5,56 Sparisoma atomarium 1,82 22,22 Sparisoma sp. 0,29 5,56 Coryphopterus sp. 0,32 11,11 NF 6,61 72,22 0,31 11,11 0,44 16,67 0,68 22,22 Scaridae Gobiidae Pristigenys alta 0,12 5,56 NF 0,39 11,11 Carangoides bartholomaei 0,16 5,56 Caranx crysus 0,34 11,11 Caranx sp. 1,81 33,33 Chloroscombrus chrysurus 1,62 27,78 Decapterus macarellus 0,30 5,56 Diplospinus multistriatus 0,05 5,56 Decapterus punctatus 0,20 5,56 Nesiarchus nasutus 0,29 11,11 Decapterus sp. 0,15 5,56 NF 0,27 11,11 Oligoplites saurus 0,16 5,56 Selar crumenophthalmus 1,12 5,56 Acanthocybium solandri 0,08 5,56 Selar sp. 0,24 5,56 Auxis sp. 1,02 22,22 Selene setapinnis 0,60 16,67 Euthynnus alleteratus 0,29 5,56 Katsuwonus pelamis 0,17 5,56 0,13 5,56 Scomberomurus caballa 0,23 11,11 Carangidae Bramidae Brama sp. Lutjanidae Lutjanus sp. 0,58 16,67 NF 0,24 11,11 Gerreidae Eucinostomus sp. 0,45 11,11 NF 0,15 5,56 Menticirrhus sp. 0,09 5,56 NF 0,29 5,56 Mullidae 0,09 5,56 Sciaenidae Pomacanthidae Centropyge argi 0,19 5,56 Microdesmidae Microdesmus sp. Acanthuridae Acanthurus sp. Sphyraenidae Sphyraena sp. Gempylidae Scombridae Scombridae sp. 2,41 44,44 Thunnus albacares 0,20 5,56 Thunnus obesus 0,20 11,11 Cubiceps pauciradiatus 0,58 5,56 Psenes sp. 0,13 5,56 Citharichthys sp. 0,69 22,22 Syacium sp. 1,97 33,33 NF 0,20 11,11 Bothus ocellatus 1,13 38,89 Monolene sp. 0,23 11,11 NF 0,08 5,56 Symphurus sp. 0,48 5,56 Symphurus tesellatus 0,16 5,56 Balistes capriscus 1,54 27,78 Canthidermis maculata 0,09 5,56 Nomeidae Paralichthyidae Bothidae Cynoglossidae Pomacentridae Stegastes sp. 0,12 5,56 NF 0,19 5,56 0,29 11,11 Mugil sp. 0,12 5,56 Sphoeroides sp. 1 4,89 44,44 NF 0,23 11,11 Sphoeroides sp. 2 4,09 44,44 Howellidae Howella sp. Mugilidae Balistidae Tetraodontidae 78 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA a b c d e f EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA g h i j k l 79 Figura 7-4 Larvas de peces encontrados durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. a) Sardina (Clupeidae); b) Pez Linterna (Diaphus sp.); c) Merlucita (Bregmaceros atlanticus); d) Melamphaes sp; e) Cherna (Serranus spp.); f) Jurel (Caranx spp.); g) Gobiidae; h) Pez cirujano (Achanthurus sp.); i) Escolar (Gemphylidae); j) Lenguado (Bothus ocellatus); k) Globo (Sphoeroides spp.); l) Balistes sp. Fotografías: Ana María Galeano, INVEMAR. 80 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA De las 1.336 especies ícticas registradas, el 82,3 % pueden observarse en el Caribe colombiano (Polanco et al., 2010), lo cual supone una alta diversidad de especies, sin embargo, Medellín et al. (2013) sugieren que se conoce muy poco acerca de la diversidad de los estadios larvales de los peces llegando solo al 8,7 % de su conocimiento. Las larvas de los peces difieren significativamente de los adultos en tamaño, forma y hábitat, haciendo compleja su identificación, ocasionando registros erróneos de las especies y tener estimaciones bajas de la diversidad. En la presente investigación, el valor promedio de diversidad de larvas de peces (entendida como diversidad de Shannon o H’) para el área se estimó en 3,0 ± 0,5 bits/larvas, siendo un valor moderadamente alto si se compara con otros estudios realizados en la zona (Vergara et al., 1999; Pabón, 2013). En las zonas centro y sur se muestrearon un mayor número de familias; mientras que en la estación E01 ubicada hacia el norte se encontró una alta dominancia de la familia Clupeidae. Esta dominancia puede estar relacionada con eventos reproductivos, donde los adultos pueden llegar a formar cardúmenes en las capas de agua superficiales o subsuperficiales dentro de golfos, bahías o en mar abierto (Cervigón, 1991). En el Caribe colombiano se ha observado que en la zona nororiental se encuentran altas densidades de adultos de engraulidos (anchoas) y clupeidos (sardinas), lo cual puede ser explicado por el efecto que ejercen las surgencia en los eventos reproductivos de poblaciones adultas de estas especies en la zona (Páramo et al., 2003; Pabón, 2013). El análisis estadístico mostró que el ensamblaje de larvas de peces fue diferente entre las zonas norte, centro y sur, en particular entre las zonas norte y centro (p<0,05), en donde la diferencia fue dada por la presencia de las familias Tetraodontidae y Clupeidae. En la zona norte-sur la diferencia estuvo marcada por representantes de las familias Gobiidae y Paralichthyidae. El patrón en la estructura comunitaria de larvas de peces pudo estar influenciado por las características oceanográficas propias de cada zona geográfica y a la presencia de familias predominantes entre estaciones de muestreo. La familia Myctophidae (peces linterna) fue la más representativa en las estaciones de muestreo de la zona norte y sur. Los myctófidos son peces que se encuentran en todos los mares y océanos del mundo. En estado adulto viven en rangos desde los 200 hasta los 2000 m de profundidad en aguas abiertas; son importantes para las redes tróficas ya que son el alimento de muchos peces en los océanos y además, su capacidad de migración a estratos más superficiales representa un gran movimiento de la energía a través de la columna de agua (Clarke, 1973; Gartner et al., 1987). Por otra parte, especies de la familia Gonostomatidae (peces luz) fueron representativas en las estaciones ubicadas en la zona centro. Al igual que los peces linterna, los gonostomátidos se encuentran ampliamente distribuidos y los adultos viven en la zona meso y batipelágica del océano. Esto demuestra que las poblaciones de larvas de peces en el Caribe colombiano tuvieron un predomino de familias típicamente oceánicas. Larvas de peces pelágicos de importancia comercial Se encontraron larvas de peces pelágicos de importancia comercial de la familia Scombridae (túnidos), tales como la bacoreta (Euthynnus alleteratus), barrilete (Katsuwonus pelamis), sierra wahoo (Acanthocybium solandri), atún aleta amarilla (Thunnus albacares) y ojo gordo (Thunnus obesus). Estas especies presentan un amplio rango de distribución en aguas tropicales y subtropicales del mundo y generalmente su hábitat está condicionado por características oceanográficas tales como la temperatura, la salinidad, frentes, corrientes entre otros, incidiendo en el desove y su posterior desarrollo (Fahay, 2007). Tambien se destacó la presencia de larvas de barracuada (Sphyraena sp.), especie que se puede encontrar en hábitats pelágicos o demersales, desde aguas costeras y arrecifes coralinos hasta aguas profundas desovando sobre la plataforma continental (Russell, 2002; Fahay, 2007). En general, se observó la presencia de larvas de grandes pelágicos distribuidas a lo largo del Caribe, con una mayor concentración en las estaciones de muestreo ubicadas en la zona norte y centro [Figura 7-5 ]. Este resultado sugiere que la zona nororiental puede ser un lugar potencial para el desove de estas especies, al igual que otras de importancia comercial que hacen parte de otras pesquerías, tales como jureles (carangidos), chernas (serranidos) y pargos (lutjanidos), predominantes en el área. Probablemente, las condiciones oceanográficas favorecen la reproducción y el desove, no obstante, se deben seguir desarrollando estudios a una escala temporal más amplia que permitan identificar épocas y establecer áreas de eventos reproductivos de los peces en el Caribe colombiano. Figura 7-5 Distribución espacial de la abundancia de larvas (larvas/10 m2) de peces pelágicos de importancia comercial durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. Fotografías: Eugenia Escarria y Ana María Galeano. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Atún aleta amarilla (Thunnus albacares) Patudo/ ojo gordo (Thunnus obesus) 81 82 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Bacoreta (Euthynnus alleteratus) Barrilete (Katsuwonus pelamis) EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Sierra wahoo (Acanthocybium solandri) Barracuda (Sphyraena spp.) 83 84 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA PACÍFICO Distribución y abundancia de ictioplancton La densidad de huevos estuvo entre 75 y 903 huevos/10 m2, siendo los más característicos los morfotipos MI (larva o vitelo pigmentado) y MIII (corion ornamentado) [Figura 7-6 ]. Las mayores densidades de huevos de peces se encontraron en las zona intermedia y oceánica del área de estudio (>500 huevos/10 m2; Tabla 7-3 ; Figura 7-7a ), sin embargo, también se encontraron densidades importantes en la zona costera, registrando huevos en el 100 % de las estaciones muestreadas. La distribución y abundancia de huevos de peces encontrada puede indicar que se presentaron desoves permanentes de peces a lo largo del Pacífico colombiano. Debido al efecto de deriva y las corrientes, los huevos pueden ser hallados en zonas oceánicas y la presencia de una alta densidad de estos en inmediaciones de la isla de Malpelo puede estar asociado a la influencia de los procesos físicos de corrientes, que inciden en el transporte Zona Intermedia Oceánica Costera a de los huevos hacia afuera de la costa (Escarria et al., 2006). En cuanto a larvas de peces, se encontraron densidades entre 70 y 1.145 larvas/10 m2 [Tabla 7-3 ] y al igual que en los huevos, las mayores abundancias de larvas se concentraron en la zona oceánica, en especial en las estaciones E06 y E07 (> 1.000 larvas/10 m2; Tabla 7-3 ; Figura 7-7b ). Tabla 7-3 Valores de abundancia (N/10 m2) de huevos y larvas de peces durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. Huevos Larvas (N/10 m ) (N/10 m2) 80°15,743 118 293 04°11,957 80°29,739 294 142 E03 03°43,724 80°19,588 75 113 E04 03°32,577 80°24,722 618 188 E05 03°02,257 80°22,700 903 96 E06 03°02,183 82°39,260 233 1.145 E07 03°37,676 82°33,148 601 733 E08 04°01,700 82°27,921 80 129 E09 04°17,502 82°24,212 512 256 E10 04°42,121 82°30,005 551 315 E11 05°58,559 78°10,203 410 246 E12 05°47,316 78°16,990 474 215 E13 05°35,312 78°25,718 244 70 E14 05°28,434 78°18,500 241 254 E15 04°49,256 78°28,801 399 426 E16 04°07,150 78°28,610 195 139 E17 03°41,744 78°16,100 515 156 Estación de muestreo Latitud (N) Longitud (W) E01 04°40,738 E02 2 b Figura 7-6 Morfotipos de huevos encontrados durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. a) y b) Morfotipo III. Fotografías: Ana María Galeano - INVEMAR. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 85 b Figura 7-7 Distribución espacial de la densidad (N/ 10 m2) de a) huevos y b) larvas durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. Composición de especies y estructura comunitaria Se recolectaron un total de 549 larvas representadas en 54 taxones (género y especie) y 38 familias. La familia más importante en abundancia fue Myctophidae (43,6 %), seguida por las familias Nomeidae, Paralepididae y Phosichthyidae con porcentajes de abundancia mayores al 5 %. Estas familias son típicas de ambientes meso y batipelágicos (Moser, 1996) y su dominancia durante este estudio pudo estar influenciada por el mayor número de estaciones de muestreo que fueron realizadas en el área más alejada de la costa (zona intermedia y oceánica). Tambien se destacaron larvas de importancia comercial como Paralichthydae (lenguados, 1,74 %), Cynoglossidae (lengüetas, 1,30 %), Coryphaenidae (dorado, 0,94 %), Mugilidae (lisas, 0,89 %); Scombridae (atunes y sierras, 0,61 %) y Engraulidae (carduma, 0,27 %) [Figura 7-8 ]. La especie más abundante y frecuente fue Diogenichthys laternatus (pez linterna), especie de hábitat batipelágico encontrada en todas las estaciones de muestreo representando el 18 % de la abundancia total [Tabla 7-4 ]. En otros estudios realizados en el Pacífico colombiano, D. laternatus ha sido una de las especies de mayor abundancia (Beltran-León, 2007; Martínez-Aguilar et al., 2010), sugiriendo su importancia ecológica en la composición de las asociaciones de larvas de peces en el Pacífico. Otras especies de peces 86 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA linterna, como Myctophum nitidulum y Vinciguerria lucetia, también fueron importantes en cuanto a su frecuencia, ya que se encontraron en más del 40 % de las estaciones de muestreo [Tabla 7-4 ]. En cuanto a la diversidad (H’), el valor promedio para el área se estimó en 2,4 ± 0,4 bits/larvas. En la zona oceánica se presentó el valor más alto de diversidad (3,3 bits/larvas), mientras que la diversidad más baja se encontró en la zona costera (1,7 bits/ larvas). En la zona intermedia y oceánica se encontró un mayor número de familias, mientras que la zona costera dominaron pocas familias. Se encontraron diferencias estadísticas en la estructura de la comunidad de larvas de peces entre zonas. Estas diferencias fueron evidentes entre la zona intermedia-costera (p<0,05), marcada por las especies D. laternatus y Bathylagus wesethi, mientras que Vinciguerria lucetia y Psenes sio fueron las que mejor explicaron las diferencias encontradas entre la zona oceánica y costera (p<0,05). Lo anterior sugiere que la comunidad de larvas de peces estuvo compuesta por especies de hábitos mesopelágicos y con características oceánicas. Los peces mesopelágicos cumplen un papel importante en las redes tróficas marinas y son un componente primordial de los sistemas oceánicos del mundo, debido a su abundancia y amplia distribución. La mayoría de sus especies experimentan migración vertical, ubicándose durante el día a profundidades entre 200 y 1.000 m, mientras que en la noche pueden encontrarse sobre los 200 m e inclusive en la superficie. Estas migraciones influyen en el comportamiento alimenticio y los patrones de distribución de estos peces. Myctophidae 43,59 Otros 9,32 % Nomeidae 6,04 % Paralepididae 5,34 % Phosichthyidae 5,25 % Bathylagidae 4,99 % Bregmacerotidae 3,84 % Hemiramphidae 2,48 % Callionymidae 1,84 % Paralichthyidae 1,74 % Scorpaenidae 1,66 % Gonostomatidae 1,45 % Cynoglossidae 1,30 % Melamphaidae 1,24 % Carangidae 1,12% Ophidiidae 1,12 % Coryphaenidae 0,94 % Mugilidae 0,89 % Stomiidae 1,15 % Scombridae 0,61 % Gempylidae 0,56 % Tetraodontidae 0,54 % Ophichthidae 0,51 % Labridae 0,50 % Carapidae 0,38 % Gobiidae 0,37 % Chiasmodontidae 0,36 Exocoetidae 0,33 % Howellidae 0,27 Engraulidae 0,27 Figura 7-8 Porcentaje de abundancia de las familias de larvas de peces durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. Tabla 7-4 Composición, porcentaje de abundancia (%A) y frecuencia de ocurrencia en porcentaje (%FO) de larvas de peces encontradas durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. NF: identificados a nivel de familia. Taxón %A %FO 0,14 5,88 0,51 11,76 0,27 5,88 0,13 5,88 Bathylagus nigrigenys 0,72 5,88 Bathylagus wesethi 1,98 Leuroglossus sp. 2,29 Muraenidae Gymnothorax sp. %A %FO Cyclothone sp. 0,66 23,53 Diplophos sp. 0,29 11,76 NF 0,50 11,76 5,25 41,18 Bathophilus filifer 0,14 5,88 35,29 Idiacanthus antrostomus 0,38 11,76 11,76 NF 0,63 11,76 Gonostomatidae Ophichthidae Ophicthus sp. Engraulidae Argentinidae Argentina aliceae Taxón Bathylagidae Phosichthyidae Vinciguerria lucetia Stomiidae 87 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Taxón %A %FO Paralepididae %A %FO 0,97 23,53 0,19 5,88 Naucrates ductor 0,75 17,65 Caranx sp. 0,37 5,88 0,94 17,65 0,27 5,88 0,89 29,41 NF Lestidiops sp. 4,75 29,41 Stemonosudis macrura 0,59 17,65 Neoscopelidae Scopelengys tristis Taxón Apogonidae Apogon atricaudus Carangidae 0,19 5,88 Myctophidae Benthosema panamense 0,19 5,88 Diaphus pacificus 4,33 35,29 Diogenichthys laternatus 17,93 82,35 Gonichthys tenuiculus 0,31 11,76 Lampanyctus parvicauda 0,67 17,65 Lampanyctus sp. 0,72 11,76 Myctophum aurolaternatum 4,50 52,94 Myctophum nitidulum 9,15 47,06 Halichoeres sp. 0,19 5,88 Myctophum sp. 0,15 5,88 Thalassoma sp. 0,31 11,76 NF 5,64 35,29 Chiasmodontidae 0,36 11,76 1,84 23,53 0,37 11,76 0,56 11,76 Bregmacerotidae Bregmaceros sp. Ophidiidae Brotula sp. Coryphaenidae Coryphaena hippurus Howellidae Howella sp. Mugilidae Mugil cephalus Labridae Chiasmodon niger 3,84 58,82 0,93 23,53 0,19 5,88 Carapidae Callionymidae Synchiropus atrilabiatus Gobiidae Gempylidae Echiodon exsilium 0,19 5,88 NF 0,19 5,88 Bythitidae 0,18 5,88 Euthynus lineatus 0,19 5,88 Lophiiformes 0,81 23,53 Katsuwonus pelamis 0,28 11,76 NF 0,14 5,88 0,19 5,88 Cubiceps pauciradiatus 1,64 29,41 4,40 29,41 Antennaridae Antennarius sp. 0,14 5,88 Melanocetidae Melanocetus johnsoni Gempylus serpens Scombridae Trichiuridae Trichiurus lepturus 0,20 5,88 Hemiramphidae Nomeidae Hyporhamphus sp. 2,29 41,18 Psenes sio Oxyporhamphus micropterus 0,19 5,88 Paralichthyidae 0,33 5,88 Syacium sp. 1,53 29,41 NF 0,21 5,88 Symphurus oligomerus 0,25 5,88 Symphurus sp. 1,04 29,41 0,54 5,88 7,07 76,47 Exocoetidae Melamphaidae Melamphaes sp. 0,88 29,41 Scopelogadus bispinosus 0,36 5,88 Cetomimidae Eutaeniophorus festivus 0,10 5,88 Scorpaenidae Pontinus sp. Cynoglossidae Tetraodontidae Sphoeroides sp. 0,69 23,53 Otros 88 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA a b c d e f EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA g h i j k l 89 Figura 7-9 Diversidad de larvas de peces encontrada durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. a) Anchoa (Cetengraulis mysticetus); b) Diaphus pacificus; c) Diogenichthys. Laternatus; d) Merlucita (Bregmaceros sp.); e) Merluza (Brotula sp.); f) Lisa (Mugil cephalus); g) Naucrates ductor ; h) Thalassoma sp.; i) Euthynus lineatum; j) Cubiceps pauciradiatus; k) Syacium sp.; l) Symphurus sp. Fotografías: Ana María Galeano - INVEMAR. 90 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Larvas de peces pelágicos de importancia comercial Se identificaron larvas de dorado Coryphaena hippurus con una frecuencia de aparición de 17,65 %, representando el 0,94 % de la abundancia total [Tabla 7-4 ]. Aunque no fue posible detectar huevos de dorado, estos eclosionan alrededor de 38 horas después del desove, basado en una relación entre diámetro de los huevos, la temperatura del agua y el tiempo de desarrollo y sus larvas viven por debajo de los 50 m de profundidad (Ditty et al., 1994). El dorado es una de las especies pelágicas más importante en el Pacífico colombiano, su época de desove posiblemente ocurre entre octubre y diciembre (Lasso y Zapata, 1999), lo cual fue confirmado parcialmente por la presencia de larvas durante el periodo del crucero (noviembre-diciembre), específicamente en las estaciones de muestreo E04 (18 larvas/10 m2), E05 (15 larvas/10 m2) y E15 (13 larvas/10 m2) [Figura 7-10 ]. Esta última estación de muestreo (E14) ubicada en la zona costera, coincide con el área de mayor abundancia de larvas de dorado reportada por Rueda et al. (2014), por lo que la aparición de esta especie en la misma zona durante el mismo periodo en diferentes años puede sugerir la ocurrencia de eventos reproductivos. Adicionalmente, se conoce que el dorado tiene preferencia por los sitios de desove, regresando a los mismos en cada periodo de reproducción (Oxenford, 1999). También se capturaron larvas de escómbridos Katsuwonus pelamis y Euthynnus lineatus (barrilete) que son especies pelágicas de interés comercial (Melo et al., 2011), frecuentes en zonas neríticas y oceánicas, con épocas de desove durante todo el año en aguas tropicales (Moser, 1996), por lo que la frecuencia de sus estados larvales no es estacional. A pesar de la presencia de algunas especies comerciales durante este estudio, su número fue reducido, lo cual puede ser atribuido a diferentes factores como la presencia de larvas en otras áreas, debido al transporte por efecto de las corrientes (Leis, 2010) o que el desove de algunas especies ocurre en otros periodos del año, entre otros. Figura 7-10 Distribución espacial de la abundancia de larvas (larvas/10 m2) del dorado (Coryphaena hippurus) durante el crucero de investigación de grandes pelágicos en el Pacífico colombiano. Fotografía: Gustavo Ramírez, INVEMAR. Dorado (Coryphaena hippurus) EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 8 93 VARIABLES OCEANOGRÁFICAS Y SU RELACIÓN CON LA ABUNDANCIA DEL RECURSO Estefanía Isaza-Toro, Alfredo Rodríguez y Mario Rueda INTRODUCCIÓN La oceanografía juega un papel muy importante en el comportamiento y desarrollo de los organismos marinos (Cury et al., 2003), por lo tanto, determinar el efecto que las variables oceanográficas tienen sobre la disponibilidad y abundancia de los recursos es fundamental para el manejo pesquero basado en el ecosistema (Botsford et al., 1997; Basson, 1999; Perry et al., 1999; Worm et al., 2009). En Colombia, se dan procesos oceanográficos importantes tales como cambios en la temperatura y oxígeno disuelto, periodos de lluvias o surgencia estacional que determinan en gran parte la productividad de las masas de agua y la disponibilidad de las poblaciones marinas (Andrade et al., 2003; Andrade y Barton, 2005; Stewart, 2008; Rodríguez-Rubio y Stuardo, 2002; Rodríguez-Rubio et al., 2003). En este sentido, este capítulo apunta a describir las variables oceanográgicas basado en información tomada in situ durante los cruceros de investigación y proveniente de sensoramiento remoto para determinar relaciones funcionales con la abundancia relativa de grandes pelágicos y del ictioplancton (huevos y larvas), las cuales no constituyen relaciones causa-efecto, pero sí la predicción de una relación que con certidumbre puede indicar la magnitud en que la variabilidad ambiental afecta la disponibilidad de recursos pesqueros. MÉTODOS ESPECÍFICOS Para caracterizar las condiciones oceanográficas fueron colectados datos a partir de dos fuentes de información: i) perfiles de temperatura, salinidad, oxígeno disuelto y pH, colectados in situ con sonda CTDO durante los cruceros de investigación (para mayor detalle del muestreo ver sección 3) y ii) datos superficiales de temperatura (TSM), salinidad, altura del nivel del mar (ANM), clorofila-a (CLA) y corrientes marinas (componente zonal U: sentido Este-Oeste y meridional V: sentido Norte-Sur), adquiridos mediante sensoramiento remoto satelital y salidas de modelos. La información satelital y modelada se descargó del portal de internet [MyOcean, Monitoring and Forecasting (www.myocean.eu)]. Este portal es de libre acceso a 127 colecciones de cubrimiento mundial, ofreciendo así un amplio espectro de información de calidad y en tiempo real de predicciones y salidas de modelos. La primera colección empleada fue Global Ocean 1/12° Physics Analysis and Forecast updated Daily, el cual es un producto generado por un sistema de predicción (modelo Nucleus for European Models of the Ocean 3.1, acoplado con el modelo Louvain sea Ice Model 2), conocido como Operational Mercator global Ocean analysis and Forecast system. Este sistema de predicción consiste en hacer análisis operacionales y pronósticos de las condiciones oceanográficas físicas desde la superficie hasta el fondo del océano, a partir de modelos y asimilación de datos y consta de las siguientes características (Drévillon et al., 2008): • Resolución espacial: 1/12°, lo cual equivale aproximadamente a 9,26 km, con perspectiva tridimensional. Abarca toda la columna de agua. • Resolución temporal: pronósticos de siete días, con actualización diaria y análisis de las condiciones oceánicas actualizado semanalmente. • Serie de tiempo: A partir de enero 01 de 2013. • Incluye campos medios diarios de temperatura, salinidad, corrientes (componente zonal y meridional), altura de nivel del mar y parámetros de hielos. • Información satelital base: ANM (Jason 2, Cryosat, Saral-Altika), TSM (Reynolds AVHRR-AMSR 1/4°) y perfiles de salinidad. 94 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA La segunda colección empleada fue Global Ocean- The Optimal Interpolation product of chlorophyll-a observation (expressed in mg/m 3, 4km) at Global level. Este producto ofrece una estimación diaria de campos de pigmentos fotosintéticos eliminando la presencia de nubes, siendo una combinación de diferentes mediciones de clorofila en el agua e interpoladas con el método “Kriging”. Las principales características de esta colección son: • Resolución espacial: 4 km, solamente a nivel superficial. • Resolución temporal: información diaria. • Serie de tiempo: A partir de abril 06 de 2013. • La generación de información parte únicamente de datos obtenidos por el sensor MODIS del satélite AQUA. La descarga de información se realizó siguiendo los manuales de usuario de cada colección: i) Product User Manual for the GLOBAL Ocean Sea Physical Analysis and Forecasting Products” para la información física (Lellouche y Regnier, 2014) y ii) Product User Manual for all Ocean Colour Products” para la información de clorofila (Gianluca et al., 2014). A partir de los datos colectados in situ se construyeron perfiles oceanográficos mediante la plataforma computacional Ocean Data View –ODV–, mientras que en la cartografía fue empelado un Sistema de Información Geográfica –SIG–. Así mismo, fue empleada la plataforma Integrated Data View –IDV– para la cartografía de variables oceanográficas adquiridas por sensoramiento remoto y modelos. Para determinar las relaciones funcionales entre las variables oceanográficas (temperatura, salinidad, clorofila-a, oxígeno disuelto, profundidad, altura del nivel de mar) y la abundancia relativa de peces (CPUE: número de individuos/h*100 anzuelos) y huevos y larvas (CPUA: número/m2), se usaron modelos aditivos generalizados (GAM). Los GAM permitieron modelar la variable respuesta (e.g. abundancia relativa) y los predictores aditivos (e.g. temperatura, clorofila-a, altura del nivel del mar, etc) mediante de funciones suavizadoras no-paramétricas (Hastie y Tibshirani, 1990; Wood, 2006): yi ~ n + i =1 fi ( X i ) + i donde y i es la variable respuesta, a el intercepto, Xi son los predictores aditivos, fi son las funciones suavizadoras y εi es el error ~N(0,σ2). Para el análisis se usó la plataforma computacional R y la librería “mgcv” (R Development Core Team, 2015). Se determinó la devianza explicada (análogo de la varianza), el nivel de significancia (valor p) y la certidumbre del modelo mediante el criterio de validación cruzada generalizado (GCV): GCV = nD ( n - m )2 donde n es el número de datos, D es la devianza explicada y m son los grados de libertad efectivos del modelo. El enfoque estadístico usado a través de los modelos GAM evitó hacer frente a los restrictivos supuestos de los modelos lineales convencionales, tomando en cuenta la naturaleza no lineal de muchos procesos biológicos y ecológicos, además de identificar las ventanas ambientales que inciden sobre rangos críticos de tolerancia de las especies. RESULTADOS Y DISCUSIÓN CARIBE Oceanografía obtenida in situ A nivel superficial se observó un gradiente de temperatura en dirección Este-Oeste, registrando valores cercanos a 27,1 °C, principalmente en la zona norte en frente de Riohacha, mientras que en la zona centro y sur (entre el frente de Barranquilla y el golfo de Morrosquillo), se obtuvieron valores más altos (≥29,1 ≤30,2 °C). El rango de temperatura comprendió valores entre 27,1 °C y 30,1 °C, con un promedio y desviación estándar de 28,8 °C ± 1,087 [Figura 8-1 ]. Figura 8-1 Distribución superficial de la temperatura (°C) durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA El oxígeno disuelto presentó un gradiente alcanzando los valores máximos (88,0 % de saturación) hacia la zona norte y valores mínimos (~76,8 %) en la zona centro-sur. Los valores del oxígeno disuelto estuvieron comprendidos entre 69,5 % y 88,4 %, con un promedio de 76,6 % ± 5,443 [Figura 8-2 ]. Figura 8-2 Distribución superficial del oxígeno disuelto (%) durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. La salinidad presentó un rango de valores entre 33,0 y 36,5 con un promedio de 35,4 ± 0,835, encontrándose los valores más bajos (<34,0) frente a Cartagena (11,0 °N y 76,0 °W) y valores altos en frente de La Guajira en la zona norte [Figura 8-3 ]. Figura 8-3 Distribución superficial de la salinidad durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. 95 96 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Con respecto al pH, este presentó un rango de valores entre 8,0 y 8,3 y un promedio de 8,2 ± 0,038. Los valores más básicos se registraron en el extremo nororiental, mientras que los registros menos básicos se encontraron a la altura de 8°N y 76°W frente al golfo de Urabá [Figura 8-4 ]. Figura 8-4 Distribución superficial del pH durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. Con respecto a los pigmentos fotosintéticos representados por la clorofila-a, se registraron concentraciones bajas cercanas a 0,1 µg/l, sin marcadas variaciones a nivel superficial a lo largo de la región muestreada [Figura 8-5 ]. Figura 8-5 Distribución superficial de la clorofila-a (µg/l) durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA El comportamiento a profundidad de las variables oceanográficas mostraron que la temperatura del agua tuvo una marcada estratificación en los primeros 30 metros de profundidad cercana a los 28,0 °C, a partir de los 50 metros se observó la termoclina y posteriormente hubo un descenso progresivo hasta alcanzar valores mínimos. Se encontró un promedio de temperatura de 25,4 °C±2,741 y un rango de valores desde 19,3 hasta 30,2 °C [Figura 8-6a ]. La salinidad tuvo un promedio de 36,1 ± 0,356 y sus valores oscilaron entre 33,1 y 36,9. Los valores más bajos estuvieron cerca de la superficie (entre 33,0 y 34,0), especialmente a la altura de 11°N y 76°W. En el extremo nororiental se registraron los mayores valores de salinidad, cercanos a 37,0 [Figura 8-6b ]. El oxígeno disuelto promedio fue 69,8 % ± 10,131, con valores mínimos y máximos entre 48,7 % y 89,0 %, respectivamente. Similar a lo registrado para la temperatura, los valores de mayor saturación se ubicaron en las primeras capas de la columna de agua, hasta aproximadamente los 50 metros. Entre los 50 y 90 metros se observó la oxiclina [Figura 8-6c ]. El pH presentó valores comprendidos entre 8,0 y 8,3 y un promedio de 8,2 ± 0,038, siendo las capas superiores de la columna más básicas [Figura 8-6d ]. La clorofila-a presentó los valores más altos entre los 50 y 90 metros de la columna de agua con 0,4 µg/l. El promedio de la clorofila fue 0,2 µg/l ± 0,112, con valores comprendidos entre 0,0 y 0,6 µg/l [Figura 8-6e ]. Figura 8-6 Comportamiento a profundidad de a) temperatura (°C), b) salinidad, c) oxígeno disuelto (%), d) pH y e) clorofila-a (µg/l) durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. a b c d e 97 98 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA El comportamiento de la salinidad en la columna de agua con se encuentra asociada con los valores mínimos de temperatura [Figura 8-7c ]. El comportamiento de la clorofila-a fue diferente a respecto a la temperatura mostró una aparente asociación entre salinidades de 36,0 y temperaturas entre 27,5 y 29,0 °C [Figura lo descrito anteriormente para las demás variables. El valor más 8-7a ]. El porcentaje de saturación de oxígeno disuelto osciló alto de clorofila registrado entre 50,0 y 90,0 metros de profunentre 95,0 % y 100,0 %, relacionado con valores de tempera- didad fue 0,4 µg/l y estuvo asociado con temperaturas cercanas tura cercanos a los 28,0 °C y posteriormente descendió entre los a 27,0 °C. Los valores mínimos registrados para la clorofila se 50 y 90 metros, alcanzando un 55,0 % y relacionado con valores encontraron en la superficie y en la máxima profundidad relacionados con temperaturas máximas (27,0 °C) y mínimas (25,0 de temperatura entre 20,0 y 24,0 °C [Figura 8-7b ]. A pesar que se registró valores de pH cercanos a 8,0 con una desviación °C), respectivamente [Figura 8-7d ]. estándar inferior a 0,05, se apreció que la región menos alcalina a b c d Figura 8-7 Comportamiento a profundidad de la temperatura (°C) de a) salinidad, b) oxígeno disuelto (%) y c) clorofila-a (µg/l) durante el crucero de investigación de grandes pelágicos en el Caribe colombiano. Oceanografía de sensoramiento remoto y salida de modelos En la [Figura 8-8 ] se presenta el comportamiento promedio de la altura del nivel del mar (ANM) y las corrientes generadas a partir de las velocidades zonales y meridionales. La variación promedio de ANM osciló entre -0,097 y 0,4 metros (-9,7 y 40 cm) ubicándose zonas de bajo nivel en el extremo suroccidental del Caribe de -0,09 metros (-9,0 cm). Asociadas con estas zonas donde el nivel del mar fue bajo se presentaron corrientes con tendencia occidental y la presencia de circulación ciclónica que pudieron haber generado divergencia en el agua. Adicionalmente, en el extremo norte de la cuenca se observó un aumento en la ANM de 0,4 metros (40 cm), región que se caracterizó por tener una circulación anticiclónica y convergente. Estas dos regiones claramente diferenciadas se separan por una región de transición o “frente” la cual abarcó casi todo el centro del Caribe colombiano [Figura 8-8 ]. El comportamiento de la temperatura superficial promedio presentó los valores más bajos (~25,0 °C) hacia la zona norte en frente de La Guajira, destacándose la presencia de un filamento de aguas frías con desplazamiento en dirección noroccidental [Figura 8-9 ]. El rango de esta variable comprendió valores desde 25,0 °C hasta 31,0 °C y en el extremo suroccidental y relacionados con la celda ciclónica descrita anteriormente [Figura 8-8 ], se registraron los valores más altos cercanos a 30,0 °C. La zona centro fue de transición con valores intermedios de temperatura entre 27,0 °C y 28,0 °C [Figura 8-9 ]. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 99 Figura 8-8 Comportamiento promedio de la altura del nivel del mar (m) y corrientes superficiales (m/s) entre el 22 de mayo y el 12 de junio de 2014 en el Caribe colombiano. CGSM: Ciénaga Grande de Santa Marta. Figura 8-9 Comportamiento promedio de la temperatura superficial del mar (TSM; °C) y corrientes (m/s) entre el 22 de mayo y el 12 de junio de 2014 en el Caribe colombiano. CGSM: Ciénaga Grande de Santa Marta. Para la salinidad se registraron valores entre 30,0 y 37,0 y dos zonas diferenciadas. En la zona norte en frente de La Guajira se presentaron valores altos de salinidad ~37,0 con presencia de filamentos desplazándose en sentido noroccidental. Sobre el margen costero entre la Ciénaga Grande de Santa Marta (CGSM) y el golfo de Morrosquillo se presentaron valores bajo ~33,0 [Figura 8-10 ]. Se observaron concentraciones altas de clorofila-a sobre el margen costero además de filamentos con dirección noroccidental los cuales alcanzaron concentraciones hasta 0,35 mg/ m3 aproximadamente. Los valores más bajos se registraron mar adentro y cercanos a 0,05 mg/m3 [Figura 8-11 ]. Las altas concentraciones de pigmentos que se observan a lo largo del borde costero, pueden estar asociadas con la descarga de los ríos, por ejemplo, el río Magdalena el cual tiene su delta entre Santa Marta y Barranquilla. Debido a que la descarga del río viene cargada de materia orgánica, los organismos metabolizan y absorben los nutrientes rápidamente, lo que podría causar un mayor incremento en la concentración planctónica. De esta manera, las concentraciones de microorganismos planctónicos son más altas frente a Barranquilla y disminuyen rápidamente mar adentro. Si bien el flujo de las corrientes evita la entrada del río Magdalena en el sistema de afloramientos de la península de La Guajira, la salida de agua dulce y de mayor temperatura proveniente del lago de Maracaibo (al lado oriental de la península), estaría afectando las mediciones de productividad, salinidad y temperatura (Andrade y Barton, 2005). En el caso de la productividad, las concentraciones de clorofila generadas a partir de los procesos de surgencia podrían ser menores en comparación de aquellas generadas por la descarga de los ríos. 100 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Figura 8-10 Comportamiento promedio de la salinidad y corrientes superficiales (m/s) entre el 22 de mayo y el 12 de junio de 2014 en el Caribe colombiano. CGSM: Ciénaga Grande de Santa Marta. Para diferenciar si las concentraciones de clorofila se deben al crecimiento de fitoplancton favorecido por la descarga de ríos o generado a partir de los proceso de afloramiento, sería nece- sario realizar análisis adicionales que involucren información más robusta tomada in situ y observaciones satelitales adicionales (Chuanmin et al., 2004). Figura 8-11 Comportamiento promedio de la clorofila-a (mg/m3) y corrientes superficiales (m/s) entre el 22 de mayo y el 12 de junio de 2014 en el Caribe colombiano. CGSM: Ciénaga Grande de Santa Marta. El comportamiento de las variables físicas en el Caribe, correspondió a lo esperado para la región durante el primer semestre del año. Esta región se encuentra modulada por el desplazamiento latitudinal de la zona de convergencia intertropical (ZCIT), la cual se caracteriza por ser una franja de bajas presiones atmosféricas asociada con vientos y precipitaciones. Lo anterior hace que la estación seca o de vientos, se presente entre los meses de diciembre y abril, mientras que las lluvias se registran entre agosto y octubre, los cuales adicionalmente son meses con baja intensidad del viento (Andrade y Barton, 2005). El efecto de vientos sostenidos sobre la superficie del agua, provoca un transporte de masas con un ángulo de 90° según la dirección del viento. Cuando el viento sopla paralelo a la costa, genera afloramientos costeros, los cuales favorecen la productividad biológica y la disponibilidad de recurso pesquero, debido al agua emerge más fría, de mayor salinidad y concentración de nutrientes. Es esta concentración de nutrientes lo que favorece procesos de fertilización del fitoplancton en la capa superficial (capa de mezcla), activando así la cadena alimenticia (Arévalo-Martínez y Franco-Herrera, 2008). Adicionalmente, las corrientes generadas por los vientos crean giros ciclónicos y anticiclónicos, los cuales, dependiendo de sus características, contribuyen en mayor o menor medida al desarrollo de zonas de productividad y pesca. En el hemisferio horte, los giros o 101 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA celdas ciclónicas se distinguen de las anticiclónicas por girar en sentido antihorario, por la divergencia de agua subsuperficial de menor temperatura a causa del bombeo de Ekman y la advección vertical. Por esta razón, hay mayor salinidad, enriquecimiento y baja concentración de oxígeno disuelto en la región (Stewart, 2008; Ruiz-Ochoa, 2011). Adicionalmente, en la región Caribe, la presencia de los vientos Alisios del norte, la rotación de la Tierra y el efecto de la fuerza de Coriolis dan origen a la Corriente del Caribe, la cual fluye en sentido oriental. A pesar de haberse presentado características propias de la época de afloramientos en el Caribe durante este estudio (regiones de bajas temperaturas, altas salinidades y concentraciones de clorofila frente a La Guajira y Santa Marta), la debilidad de los mismos puede deberse a que mayo y junio son meses de transición entre la estación de fuertes vientos y sequía hacia aquella caracterizada por vientos débiles y lluvias (Andrade y Barton, 2005; Arévalo-Martínez y Franco-Herrera, 2008). De acuerdo con la literatura se conoce que la existencia de un gradiente horizontal de temperatura en la cuenca Caribe colombiana permite identificar cuatro regiones con marcados contrastes climatológicos. Destaca que la zona norte que corresponde a la región costera de La Guajira con aguas más frías, que en contraste con la región al suroeste cerca del golfo de Morrosquillo de aguas más cálidas, en la cual los cambios estacionales son menos pronunciados (Ruiz-Ochoa, 2011). Relación entre variables oceanográficas y la abundancia del recurso Los resultados de la modelación GAM de la abundancia relativa o captura por unidad de esfuerzo -CPUE- (Número de individuos/h*100 anzuelos) integrando la abundancia de todas las especies de grandes pelágicos, mostraron que la temperatura superficial del mar (TSM) y la clorofila-a fueron los predictores oceanográficos de asociación significativa (p<0,05) (Tabla 81). Se identificó la mayor abundancia relativa a temperaturas menores de 27,5 °C y a concentraciones de clorofila-a Grupo/ Especie Temperatura superficial del mar (°C) Clorofila-a (mg/m3) entre 0,15 y 0,22 mg/m3; aunque se observó un segundo pico de alta abundancia relativa a concentraciones de clorofila-a entre 0,45 y 0,55 m/m3 [Figura 8-12 ]. Con respecto al análisis por especie, solo el pez vela I. albicans presentó asociación significativa con temperaturas del agua menor a 27,3 °C, salinidad mayor a 36,45 y espacialmente hacia la zona norte del Caribe a los 73 °W [Tabla 8-1 ; Figura 8-12 ]. En el atún aleta amarilla T. albacares y el dorado C. hippurus, las asociaciones con las variables oceanográficas no fueron significativas [Tabla 8-1 ; Figura 8-12 ]. La abundancia relativa o captura por unidad de área -CPUA(Número de individuos/m2) de huevos y larvas mostraron asociación significativa con la altura del nivel del mar (ANM) (p<0,05), encontrando mayor abundancia en un rango de valores entre 0,08 y 0,2m [Tabla 8-2 ; Figura 10-2 ]. Aunque no se presentaron asociaciones significativas con la temperatura y la salinidad, se conoce que estas variables son determinantes en la abundancia y la ocurrencia de las larvas en el Caribe colombiano (Arteaga et al., 2004). En el Caribe, la surgencia estacional frente a la costa de La Guajira (Andrade y Barton, 2005), se relaciona con la presencia en superficie de masas de agua de menor temperatura y altas concentraciones de clorofila-a. En la zona sur, se presentan filamentos con concentraciones de clorofila cercanas a 0,4 mg/m3, los cuales se desplazan con tendencia noroccidental desde el borde costero hacia el centro del Caribe. De acuerdo con lo anterior, las condiciones oceanográficas particulares que se presentan tanto en la zona norte y sur del Caribe, son favorables para la disponibilidad y alta abundancia de recursos pesqueros. Tabla 8-1 Modelación GAM usada para la relación entre la captura por unidad de esfuerzo -CPUE- (Número de individuos/h*100 anzuelos) y los predictores [temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m) y longitud (°)], en el Caribe colombiano. Dev.Exp: Devianza explicada (%). GCV: criterio de validación cruzada generalizado. Salinidad Nivel del mar (m) Longitud (°) p-value Dev.exp. (%) GCV p-value Dev.exp. (%) GCV p-value Dev.exp. (%) GCV p-value Dev.exp. (%) GCV p-value Dev.exp. (%) GCV Pelágicos (+) 0,04* 47,80 0,013 0,04* 62,40 0,014 0,15 51,30 0,016 0,132 23,30 0,017 0,10 28,70 0,016 I. albicans (pez vela) 0,00* 96,70 0,001 - - - 0,00* 94,00 0,001 - - - 0,00* 83,80 0,001 T. albacares (atún aleta amarilla) 0,54 21,20 0,007 0,19 50,50 0,007 0,31 20,50 0,007 0,89 0,12 0,007 0,95 0,02 0,007 C. hippurus (dorado) 0,22 43,40 0,008 0,49 3,93 0,009 0,27 9,91 0,009 0,58 2,68 0,009 0,28 44,70 0,008 (+): Abundancia relativa o CPUE integrada de todos los peces grandes pelágicos capturados durante el crucero. * Predictores significativos (p<0,05). 102 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Tabla 8-2 Modelación GAM usada para la relación entre la captura por unidad de área -CPUA- (Número de individuos /m2) del ictioplancton y los predictores [temperatura superficial del mar (°C), clorofila-a (mg/ m3), salinidad, altura del nivel del mar (m) y longitud (°)], en el Caribe colombiano. Dev. Exp: Devianza explicada (%). GCV: criterio de validación cruzada generalizado. Grupo Temperatura superficial del mar (°C) Clorofila-a (mg/m3) Salinidad Nivel del mar (m) Longitud (°) p-value Dev.exp. (%) GCV p-value Dev.exp. (%) GCV p-value Dev.exp. (%) GCV p-value Dev.exp. (%) GCV p-value Dev.exp. (%) GCV Huevos 0,24 20,70 554,6 0,55 2,33 647,6 0,55 3,46 643,4 0,00* 98,90 28,9 0,27 20,20 561,3 Larvas 0,65 29,10 8.731,4 0,48 3,21 7.522,3 0,56 2,18 7.603,0 0,03* 78,00 4.645,3 0,90 0,09 7.764,6 * Predictores significativos (p<0,05). Figura 8-12 Modelación GAM de relación entre la captura por unidad de esfuerzo -CPUE(Número de individuos/h*100 anzuelos) y los predictores [temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m) y longitud (°)], en el Caribe colombiano. Se muestran en color gris los intervalos de confianza. (+): Abundancia relativa o CPUE integrada de todos los peces grandes pelágicos. 103 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Figura 8-13 Modelación GAM de relación entre la captura por unidad de área -CPUA- (número de individuos/m2) del ictioplancton y los predictores [temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m) y longitud (°)], en el Caribe colombiano. Se muestran en color gris los intervalos de confianza. PACÍFICO Oceanografía obtenida in situ Los datos oceanográficos tomados in situ corresponden a cuatro estaciones de muestreo de un total de 17 proyectadas, debido a dificultades técnicas en la operación de la sonda oceanográfica durante el crucero de investigación. En general, se observó en las capas superficiales condiciones de baja salinidad, alto porcentaje de oxígeno disuelto y valores altos de pH [Figura 8-14 ]. La salinidad en superficie ~30 se encontró a una temperatura aproximada de 26 °C en los primeros 30 metros de profundidad, ubicando la haloclina entre los 40 y 90 metros. Los valores de salinidad abarcaron un rango de 13 a 32 y un promedio de 30,4 ±1,3 [Figura 8-14a ]. La temperatura osciló en un intervalo de 14 ° hasta 27,3 °C y un promedio de 20 °C ±5,3. El porcentaje de oxígeno disuelto alcanzó sus valores más altos (70 % de saturación) en los primeros 30 metros [Figura 8-14b ]. La oxiclina se encontró a la misma profundidad que la haloclina, mientras que los valores del pH abarcaron un rango desde 8,4 hasta 9 y un promedio de 8,6 ± 0,2, localizando cambios abruptos en los valores entre 60 y 90 metros de profundidad [Figura 8-14c ]. a b c Figura 8-14 Comportamiento a profundidad de a) salinidad, b) oxígeno disuelto (%) y c) pH, en el Pacífico colombiano. 104 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Oceanografía de sensoramiento remoto y salida de modelos Las corrientes superficiales fluyeron en dirección este, curvándose hacia el norte en cercanía del margen costero. La ANM siempre fue positiva, oscilando entre 0,2 y 0,4 metros (20 y 40 cm) [Figura 8-15 ]. Los valores más altos se registraron cerca del margen costero y los más bajos fueron en mar abierto, principalmente con una celda de ciclónica ubicada en el norte. El resto de la cuenca tuvo una altura intermedia de 0,36 metros (36 cm) [Figura 8-15 ]. Figura 8-15 Comportamiento promedio de la altura del nivel del mar (m) y corrientes superficiales (m/s) entre el 20 de noviembre y el 17 de diciembre de 2014 en el Pacífico colombiano. La temperatura presentó un gradiente desde el extremo sur de la cuenca hasta el norte, abarcando un rango desde 26 hasta 29 °C. En el margen costero, a la altura de las bahías de Tumaco y Buenaventura se presentaron las temperaturas más altas. La celda ciclónica descrita anteriormente se relacionó con los valores más altos de temperatura entre ~28 y 28,5 °C [Figura 8-16 ]. Figura 8-16 Comportamiento promedio de la temperatura superficial del mar (TSM; °C) y corrientes superficiales (m/s) entre el 20 de noviembre y el 17 de diciembre de 2014 en el Pacífico colombiano. Desde el extremo sur hasta la mitad de la cuenca cerca de la bahía de Buenaventura, se presentaron los valores más altos de salinidad (~32) y a partir de ese punto y hasta el extremo norte, las salinidades bajaron a ~28. La presencia de corrientes costeras en dirección norte y la celda ciclónica descrita, esta- rían asociadas con bajas salinidades. La región costera de la Cuenca Pacífica Colombiana (CPC) y sus ~80.000 km2 de superficie recibe la influencia directa de la hoya del río San Juan en el norte (7°28’ N), el Río Patía y su afluente el río Guaitará en el sur (0°14’N), el nacimiento del río San Juan al este (75°51’W) EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA y desembocadura del río Mira en Cabo Manglares (79°02’W), además de numerosos afluentes menores provenientes de estos ríos (Tejada et al., 2003). La capa de agua dulce proveniente de los ríos y procesos de precipitación tienen una menor densidad, lo cual la ubica encima de las demás capas de agua de mayor salinidad. Las corrientes que fluyen en dirección norte podrían estar transportando estas masas de agua de menor salinidad hacia el extremo norte de la CPC. Para el caso del Pacífico 105 Oriental Ecuatorial al cual pertenece la CPC, la migración de la ZCIT es el regulador del aporte nutricional y a su vez de productividad en la región. Durante la realización el periodo de estudio (noviembre-diciembre) y según el patrón de migración de la ZCIT, su posición al sur en la cuenca se relaciona con vientos débiles e incremento de precipitaciones, lo cual aumenta también la descarga de ríos (Tejada et al., 2003) [Figura 8-17 ]. Figura 8-17 Comportamiento promedio de la salinidad superficial del mar y corrientes superficiales (m/s) entre el 20 de noviembre y el 17 de diciembre de 2014 en el Pacífico colombiano. Se observó un gradiente de clorofila-a entre ~0,1 y 0,4 mg/m3 desde el margen costero hacia la región oceánica. Las celdas ciclónicas y anticiclónicas, mencionadas anteriormente, estuvieron asociadas a concentraciones de clorofila entre 0,2 y 0,3 mg/m3 [Figura 8-18 ]. El incremento de las lluvias trae como consencuencia un aumento en la disponibilidad nutricional y a su vez en la productividad en los márgenes costeros. Altas concentraciones de clorofila en el borde costero de la CPC puede deberse al crecimiento del fitoplancton, favorecido por la asimilación de los nutrientes en la zona eufótica y que a su vez son transportados por el sistema de corrientes (Gómez y Martínez, 2005). Figura 8-18 Comportamiento promedio de la clorofila-a (mg/m3) y corrientes superficiales (m/s) entre el 20 de noviembre y el 17 de diciembre de 2014 en el Pacífico colombiano. 106 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA La CPC también es altamente influenciada por el desplazamiento integrando la abundancia de todas las especies de grandes de la ZCIT, además del régimen de vientos y sus fluctuaciones. pelágicos, se asoció a mayores concentraciones de clorofila-a (~0,195 mg/m3) y ANM entre 0,360 y 0,365 m (p<0,05) [Tabla Lo registrado para las variables físicas, la dirección de las corrientes y la clorofila corresponde con lo observado previa- 8-3 ]. Para el dorado C. hippurus la TSM, la salinidad, la latitud mente por varios autores (Wooster, 1959; Wyrtki, 1965 y Parra, y la longitud fueron los predictores de asociación significativa 1977). Durante el último cuatrimestre del año, la ZCIT empieza (p<0,05) [Tabla 8-3 ]. La alta abundancia se asoció con valores su desplazamiento hacia el sur favoreciendo el ingreso de los de temperatura entre 27,5 y 27,7 °C, una baja salinidad ~30 y, vientos Alisios del noreste provenientes desde la región Atlán- espacialmente hacia la zona costera y norte del Pacífico (5-6°N; tica norte a la cuenca. Esta entrada de energía, representada ~78°W) [Figura 8-19 ]. Respecto al ictioplancton, solo se presentó por el chorro de viento comúnmente llamado Jet de Panamá, asociación significativa de la abundancia de larvas con la ANM genera giros ciclónicos (alta presión y bajo nivel del mar). A su y la longitud [Tabla 8-4 ; Figura 8-20 ]. La mayor abundancia de vez, la acción divergente del Jet de Panamá trae como conse- larvas de peces se encontró asociada a las aguas más oceánicas cuencia surgencias oceánicas principalmente en el centro (81 y 82° W) y bajo nivel del mar ~0,33 m (divergencia del agua) del Pacífico colombiano, representado por regiones de mayor [Figura 8-20 ]. El Pacífico tiene importantes aportes de numerosos salinidad y bajas temperaturas (Rodríguez-Rubio y Schneider, ríos, que contribuyen al incremento en la concentración de cloro2003). La descarga de los ríos en el margen costero tiene un fila debido a la generación de nutrientes (Diaz-Ochoa y Quiñones, importante efecto fertilizante, dado que proporciona materia 2008). Este aporte de los ríos, sumado a la alta productividad orgánica asimilable por diversos organismos heterotróficos que primaria producto de la surgencia en la zona norte con aguas frías, al metabolizarla, la ponen a disposición activando así la bomba ofrece condiciones favorables para los eventos reproductivos de biológica. Adicionalmente, otro factor importante relacionado especies de importancia comercial tales como el dorado. De igual con las altas concentraciones de pigmentos costeros puede ser forma, se conoce que los eddies o remolinos descendentes que se la batimetría. En regiones más someras, la actividad fotosinté- generan en el norte están relacionados con las zonas potenciales tica se incrementa debido a una mayor influencia de la radiación para la pesca de dorado (Selvaraj et al., 2011). solar (Isaza-Toro, 2013; Casanova et al., 2012). Tabla 8-3 Modelación GAM usada para la relación entre la captura por unidad de esfuerzo -CPUE- (Número de individuos/h*100 anzuelos) de grandes pelágicos y los predictores: temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m), latitud (°) y longitud (°), en el Pacífico colombiano. Dev.Exp: Devianza explicada (%). GCV: criterio de validación cruzada generalizado. Relación entre variables oceanográficas y la abundancia del recurso En el Pacífico, la alta abundancia relativa o captura por unidad de esfuerzo -CPUE- (Número de individuos/h*100 anzuelos) Temperatura superficial del mar (°C) Grupo/ Especie p-value Dev. exp. (%) Clorofila-a (mg/m3) GCV p-value Dev. exp. (%) GCV Salinidad p-value Dev. exp. (%) Nivel del mar (m) GCV Pelágicos(+) 0,14 13,80 0,529 0,00* 87,60 0,205 C. hippurus (Dorado) 0,03* 37,50 0,107 0,09 27,20 0,119 0,02* 35,10 0,103 p-value Dev. exp. (%) 0,10 16,70 0,511 0,00* 98,30 GCV 0,05 Latitud (°) p-value Dev. exp. (%) Longitud (°) GCV p-value Dev. exp. (%) GCV 0,09 19,60 0,50 0,14 13,90 0,529 0,07 28,60 0,121 0,03* 43,40 0,09 0,04* 27,60 0,115 (+): Abundancia relativa o CPUE integrada de todos los peces grandes pelágicos capturados durante el crucero, excluyendo a tiburones. * Predictores significativos (p<0,05). Tabla 8-4 Modelación GAM usada para la relación entre la captura por unidad de área -CPUA(Número de individuos /m2) del ictioplancton y los predictores: temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m), latitud (°) y longitud (°), en el Pacífico colombiano. Dev.Exp: Devianza explicada (%). GCV: criterio de validación cruzada generalizado. Grupo Temperatura superficial del mar (°C) p-value Dev.exp. GCV (%) Clorofila-a (mg/m3) p-value Dev.exp. GCV (%) Salinidad p-value Dev.exp. GCV (%) Larvas 0,10 45,70 63521 0,35 19,00 80061 0,35 5,94 Huevos 0,41 10,80 57554 0,48 12,30 56323 * Predictores significativos (p<0,05). 0,79 60660 0,56 Nivel del mar (m) p-value Dev.exp. GCV (%) 83344 0,04* 80,90 60759 0,97 0,01 60944 Latitud (°) p-value Dev.exp. GCV (%) Longitud (°) p-value Dev.exp. GCV (%) 0,36 10,70 80680 0,01* 83,80 39371 0,59 11,50 56850 0,63 1,63 59958 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 107 Figura 8-19 Modelación GAM de relación entre la captura por unidad de esfuerzo -CPUE- (Número de individuos/h*100 anzuelos) de grandes pelágicos y los predictores: temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m), latitud (°) y longitud (°), en el Pacífico colombiano. Se muestran en color gris los intervalos de confianza. Figura 8-20 Modelación GAM de relación entre la captura por unidad de área -CPUA- (Número de individuos/m2) del ictioplancton y los predictores: temperatura superficial del mar (°C), clorofila-a (mg/m3), salinidad, altura del nivel del mar (m), latitud (°) y longitud (°), en el Pacífico colombiano. Se muestran en color gris los intervalos de confianza. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 9 109 ESTRATEGIAS DE MANEJO PARA LA PESQUERÍA DE GRANDES PELÁGICOS BASADA EN EL ENFOQUE ECOSISTÉMICO Alfredo Rodríguez y Mario Rueda Según la Organización de las Naciones Unidas para la Alimentación y la Agricultura –FAO–, el manejo pesquero se define como el conjunto de “los procesos integrados de recopilación de información, análisis, planificación, consulta, toma de decisiones, asignación de recursos, implementación y formulación de normas que regulen las actividades pesqueras con el fin de asegurar la continua productividad de los recursos…” (FAO, 1997). Si bien en un contexto amplio el concepto de manejo pesquero demuestra la importancia de contar con procesos sistemáticos e integrados dirigidos a un buen funcionamiento de las pesquerías, en la actualidad los altos niveles de explotación y el consecuente declive de algunos recursos pesqueros a nivel mundial, indican un escenario contrario con vacíos y dificultades en la administración pesquera y en algunos casos también evidencian la ausencia o inadecuada gestión y aplicación de planes de ordenamiento y manejo pesquero (FAO; 1996; Steele y Hoagland, 2003; Ward et al., 2004). Probablemente, estas dificultades en la aplicación del manejo pesquero se han dado porque esta dirige su atención estrictamente al recurso pesquero como una provisión de alimento e ingresos económicos mediante el manejo de la actividad pesquera. En contraste, en la práctica lo que se requiere es un enfoque ecosistémico de la pesca (EEP), es decir, acomodar las expectativas y necesidades de la sociedad en el manejo pesquero sin comprometer los beneficios de las generación futuras y los bienes y servicios ecosistémicos (FAO, 2003), buscando así un equilibrio entre la protección de los componentes biofísicos y procesos del ecosistema y el aprovechamiento biológico y socioeconómico de los recursos (Purcell, 2010). Esto quiere decir que para lograr un buen funcionamiento de las pesquerías se deben tener en cuenta los siguientes criterios: i) Biológicos: evaluaciones que den cuenta del estado y potencial del recurso pesquero y establezcan limites en el esfuerzo y la presión de pesca para que las poblaciones marinas sean sustentables (FAO, 1995; Hindson et al., 2005); ii) Socioecómicos: asegurar los intereses culturales y económicos de los pescadores y demás usuarios, al mismo tiempo que se minimizan los impactos sobre las poblaciones marinas y los ecosistemas (FAO, 1995; Hindson et al., 2005); iii) Ambientales: mantener la biodiversidad marina, evitando y mitigando los impactos sobre los ecosistemas mediante el uso de estrategias de manejo y regulaciones eficientes (FAO, 1995; Hindson et al., 2005); iv) Gobernanza: garantizar el cumplimiento de los objetivos y metas de manejo, mediante una interacción continua y participativa en todos los niveles del sector pesquero basada en una solución estratégica de conflictos y además, en donde las acciones de gobernanza se encuentren fundamentadas y apoyadas en la ciencia y tecnología (Hindson et al., 2005; Hilborn, 2006; Mahon et al., 2009; Gutiérrez et al., 2011). Partiendo de la anterior conceptualización, este capítulo reúne y consolida los principales aportes de la investigación, presentando así un modelo conceptual de manejo [Figura 9-1 ], indicadores de valor y estrategias de manejo y conservación [Tabla 9-1 ] que abordan aspectos biológico-pesqueros, ecológicos y tecnológicos, con el objeto de brindar al sector pesquero herramientas de uso en la toma de decisiones para el aprovechamiento sustentable de los recursos grandes pelágicos en aguas marinas jurisdiccionales de Colombia. 110 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA INVESTIGACIÓN Institutos Universidades Organizaciones Gremios Otros de ncia sfere Tran mend Reco nejo e ma es d acion d etas M o nej e ma RECURSO PESQUERO Y ECOSISTEMA y traba Uso res Solució Institucionalidad nflicto y ordena GOBERNANZA cipa;vo le n de co Manejo jo par; ponsab s miento USUARIOS Pescadores Comercializadores Empresas pesqueras Figura 9-1 Modelo conceptual del sistema de manejo pesquero para el recurso grandes pelágicos en Colombia. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Tabla 9-1 Indicadores y medidas de manejo y conservación para recursos grandes pelágicos en Colombia. Indicador Región Recurso Atún aleta amarilla (Thunnus albacares) Caribe Bes/Bexp = Biomasa estimada (Bes) sobre la Biomasa explotada por pesca (Bexp) Dorado (Coryphaena hippurus) Valor Indicador de estado y riesgo del recurso Bexp = 5.700 t 1 Bexp/Bes ≥ 0,9 ≤ 1 Agotado o colapso Bes = 1.700 t Bexp = sin información disponible Bexp/Bes ≥ 0,7 ≤ 0,9 Sobreexplotado Dorado (Coryphaena hippurus) Bexp = 313 t 2 Cuota de pesca anual que no exceda las 4.000 t. - Cuota anual de pesca en 2.000 t. Bexp/Bes ≥ 0,5 ≤ 0,7 Moderadamente explotado Bexp/Bes ≥ 0,2 ≤ 0,5 Plena explotación Bes = 3.900 t Medida de manejo / conservación Bexp/Bes 3 0,5 ≤ 0,7 Moderadamente explotado Bes = 9.200 t Si Pacífico Estado actual Bexp/Bes 3 0,1 ≤ 0,2 Subexplotado Bexp/Bes ≥ 0,1 ≤ 0,2 Subexplotado TMM = 127,8 cm.LH 3 Atún aleta amarilla (Thunnus albacares) %C < TMM = Porcentaje de captura (TMC) por debajo de la Talla Media de Madurez (TMM) Para el atún aleta amarilla se sugiere una talla mínima de captura igual a 128 cm.LH. TMC = 132,7 cm.LH 3 > 50% C Riesgo alto de sobrepesca por crecimiento %C < TMM = 29,5% 3 Caribe TMM = 82,7 cm.LH 3 Dorado (Coryphaena hippurus) TMC = 113,4 cm.LH 3 Si %C < TMM = 11,9% 3 < 30% C Riesgo bajo de sobrepesca por crecimiento TMM = 82,7 cm.LH 3 Pacífico Caribe Dorado (Coryphaena hippurus) Atún aleta amarilla (Thunnus albacares) ≥ 30 ≤ 50% C Riesgo moderado de sobrepesca por crecimiento TMC = 104,6 cm.LH 3 %C < TMM = 6,5% 3 < 30% C Riesgo bajo de sobrepesca por crecimiento Liberación de la captura viva de especies objetivo de tallas menores (juveniles). < 30% C Riesgo bajo de sobrepesca por crecimiento < 30% C Riesgo bajo de sobrepesca por crecimiento FA:CO 3 1:3 ≤ 1:5 Riesgo bajo sobre biodiversidad FA:CO = 2,2:1 3 Para el dorado se sugiere una talla mínima de captura igual a 83 cm.LH. Palangre o longline con operación de anzuelos a profundidad mínima de 50 m, para mitigar captura incidental de especies más cercanas a la superficie. Uso de anzuelos circulares tipo atunero calibre 13/0. FA:CO = Relación en peso de la fauna acompañante (FA) versus captura objetivo (CO) Usar carnada dirigida a peces grandes pelágicos, por ejemplo, calamar y pequeños scombridos. FA:CO > 1:5 o mayor Riesgo alto sobre biodiversidad Pacífico Dorado (Coryphaena hippurus) Si FA:CO = 4,8:1 ³ FA:CO ≥ 1:3 ≤ 1:5 Riesgo medio sobre biodiversidad FA:CO ≥ 1:1 ≤ 1:3 Riesgo bajo sobre biodiversidad No usar calamar como carnada en los anzuelos más cercanos a la línea principal para mitigar captura de tortugas. FA:CO ³ 1:3 ≤ 1:5 Riesgo medio sobre biodiversidad No realizar a la carnada un corte en la parte abdominal, para evitar la exposición de visceras y sangrado en el agua, lo cual actua como cebo principalmente para tiburones. El material de las líneas secundarias reinales debe ser en nylon monofilamento (no usar guaya o material metálico), para facilitar el escape de tiburones. Liberación de la captura viva de especies incidentales de tallas menores, mediante corte en la línea de nylon. Liberación de todas las tortugas enganchas en los anzuelos, subiendo en lo posible el individuo a bordo para corte de la línea de nylon y extracción del anzuelo. 1 Biomasa anual desembarcada por la flota pesquera de longline en el Caribe colombiano. Fuente: Documento técnico de cuotas 2011. 2 Biomasa total anual desembarcada por la flota industrial y artesanal en el Pacífico colombiano. Fuente: Boletín anual SEPEC 2013. 3 Valores estimados en este estudio. 111 112 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 10 CONCLUSIONES Algunas conclusiones al margen de los resultados de esta investigación fueron: i) El uso de barcos de pesca comercial como plataformas de investigación que consolidaron la combinación del conocimiento tradicional con el científico en un marco de investigación participativa; ii) El carácter sinóptico de los cruceros de investigación permitió cubrir en relativo poco tiempo gran cobertura espacial para toma de información ecosistémica usando tecnologías de investigación pesquera avanzada; y iii) El trabajo interinstitucional e intersectorial donde tanto los pescadores industriales como la AUNAP, institutos de investigación, la academia y ONG, pueden unirse para mejorar los resultados obtenidos y viabilizar sus alcances. Los principales hallazgos de esta investigación fueron: • El crucero de grandes pelágicos realizado en el Caribe continental obtuvo una captura de 8.385 kg correspondientes a 311 individuos identificados dentro de 16 especies agrupadas en diez familias, siendo las más abundantes en peso Scombridae (Thunnus albacares; 2.635 kg) y Carcharhinidae (Carcharhinus falciformis; 1.211 kg). Las mayores abundancias relativas promedio (0,56 N/100 anzuelos*h), se encontraron frente a las costas de Córdoba y el golfo de Urabá. • En el Pacífico se capturaron 5.804 kg correspondientes a 282 individuos identificados dentro de 11 especies agrupadas en siete familias, entre las cuales se destacaron las familias Istiophoridae (Istiophorus platypterus, 1.666 kg) y Coryphaenidae (Coryphaena hippurus, 1.005 kg). La mayor abundancia relativa promedio ocurrió en la zona costera (1,3 N/100 anzuelos*h). • Para el Caribe continental, se estimó la biomasa instantánea de atún aleta amarilla (T. albacares) de 9.200 t para en un área efectiva de 1.236 mn2); mientras que para el dorado (C. hippurus) la biomasa fue 1.700 t en un área de 530 mn2. El atún mostró un parche de densidad alta en la zona sur frente al golfo de Morrosquillo (hasta 334 t/mn2), en tanto el dorado se agregó con mayor densidad (hasta 113 t/mn2) frente de Barranquilla en la zona centro. Individuos más grandes de atún (entre 164 y 191 cm LT), se hallaron en la zona norte. • Para el Pacífico, se estimó una biomasa instantánea de dorado (C. hippurus) de 3.900 t en un área efectiva de 1.097 mn2, con un parche de densidad alta frente a Buenaventura. Se identificaron agregaciones de individuos maduros en la zona costera norte entre Cabo Corrientes y bahía Solano; mientras que en la misma zona, pero costa afuera, se encontró alta abundancia de larvas. • En el Caribe continental se determinó que más del 85,1% de los individuos capturados de atún aleta amarilla, estuvieron por encima de la TMM. Así mismo se evidenció una etapa reproductiva adulta en la mayoría de dorados y picudos. La información colectada permitió desarrollar la descripción histológica de estados de madurez de T. albacares, como un valioso aporte al conocimiento de la biología de esta especie e insumo para las medidas de conservación y manejo. • En el Pacífico, se encontró predominancia de estados maduros de C. hippurus (85,8% de los individuos capturados por encima de la TMM), cuya descripción histológica de material gonadal para machos y hembras fue desarrollada. • La mayor densidad de huevos (>90 huevos/10 m2) y larvas (>178 larvas/10 m2) en el Caribe continental ocurrieron EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA en la zona norte, lo que indicaría ser un área de potencial desove. La mayores densidades de larvas de grandes pelágicos estuvieron representadas por Thunnus obesus, T. albacares, Katuswonus pelamis y Acanthocybium solandri; las cuales mostraron una agregación de mayor abundancia en la zona norte y centro. • La abundancia relativa de huevos y larvas en el Caribe mostraron asociación significativa con la altura del nivel del mar (ANM) (p<0,05), encontrando mayor abundancia en un rango de valores entre 0,08 y 0,2 m. • En el Pacífico, las mayores densidades tanto de huevos (>500 huevos/ 10 m2) y larvas (>1.000 larvas/ 10 m2), se encontraron en la zona oceánica. Se determinó la presencia de larvas de dorado C. hippurus (17,65 % de frecuencia de aparición), lo que sugiere la ocurrencia de eventos reproductivos para esta especie en el Pacífico. 113 • En las campañas realizadas en el Caribe continental (mayojunio de 2014), la variabilidad ambiental, medida por diferentes variables oceanográficas, permitió establecer que zonas de alta productividad asociadas a termoclinas estuvieron entre los 50 y 75 m de profundidad (Caribe continental). Se determinó que la temperatura superficial del mar y la clorofila-a fueron predictores significativos de la abundancia relativa de peces grandes pelágicos. • La información oceanográfica del Pacífico durante 2014, se caracterizó por la ocurrencia de frentes térmicos asociados con celdas ciclónicas o anticiclónicas que ocurrieron en límites con Panamá demarcando áreas productivas de interés para la pesca. Específicamente, altas abundancias de grandes pelágicos se asociaron a mayores concentraciones de clorofila-a (~0,195 mg/m3) y ANM entre 0,360 y 0,365 m. 114 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA BIBLIOGRAFÍA biano durante un ciclo anual (2000 2001). Ciencias Marinas, 30(3): 381-390. Thesis, University College of North Wales, Bangor, UK. 37 pp. AUNAP-Unimagdalena. 2013. Sepec Boletín Estadístico enero-diciembre de 2013. Convenio 0058 de 2013 entre la Autoridad Nacional de Acuicultura y Pesca AUNAP y la Universidad del Magdalena. 84p. Bernal, G., G. Poveda, P. Roldán y C. Andrade. 2006. Patrones de variabilidad de las temperaturas superficiales del mar en la costa Caribe colombiana. Rev. Acad. Colomb. Cienc., 30 (115): 195-208. Bertrand, A. & E. Josse. 2000. Tuna target-strength related to fish length and swinbladder volume. ICES Journal of Marine Science., 57: 1143-1146. Andrade, C.A. 1993. Análisis de la velocidad del viento en el mar Caribe. Boletín Científico CIOH., 13: 33-44. Avendaño, R., R. de Silva-Dávila, F.E. OrdóñezGuillén y G. Vázquez-López. 2010. Composición estacional de larvas de peces frente a Baja California Sur (primavera y otoño de 2003). Pp. 413-431. En: DurazoArvizu, R. y G. Gaxiola-Castro (Eds.). Dinámica del ecosistema pelágico frente a Baja California 1997–2007. SEMARNAT-INE-CICESE-UABC. Andrade, C.A. 2000. Circulation and variability of the Colombian Basin in the Caribbean Sea. Tesis doctoral de la Universidad de Wales, U.K., Menai Bridge. 223 p. Basson, M. 1999. The importance of environmental factors in the design of management procedures. ICES Journal of Marine Science., 56: 933 – 942. Andrade C.A., E.D. Barton & CHNK Mooers. 2003. Evidence for an eastward flow along the Central and South American Caribbean Coast. Journal of Geophysical Research., 108: 1-11. Ballesteros, C.A. 2007. La pesquería industrial de tiburones en el archipiélago de San Andrés, Providencia y Santa Catalina: una primera aproximación. Trabajo de Grado (Biólogo Marino). Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Santa Marta. 86 p. Aceves G y R., Saldierna. 2013. Curso de posgrado: Técnicas de muestreo e identificación de huevos y larvas de peces. Programa de postgrado en ciencias Biología. Universidad del Valle. Albaret, J.J., 1977. La reproduction de l’albacore (Thunnus albacares) dans le golfe de Guinée. Cah. ORSTOM (Sér. Océanogr.)., 15(4): 389-419. Anderson, M.J., R.N. Gorley y K.R. Clarke. 2008. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods. PRIMER-E, Plymouth, UK. Andrade, C.A. y E.D. Barton. 2005. The Guajira upwelling system. Cont. Shelf Res., 25(9): 1003-1022. Arevalo-Martinez, D. L y A. Franco-Herrera. 2008. Características oceanográficas de la surgencia frente a la ensenada de Gaira, departamento de Magdalena, época seca menor de 2006. Bol. Invest. Mar. Cost., 37(2): 131-162. Arteaga, E., Hernández, M. I. C., J. Altamar., y L. Manjárres. 2004. Distribución y abundancia de larvas de peces de la familia Lutjanidae en la zona nororiental del Caribe colom- Beardsley, G.L. jr. 1967. Age, growth and reproduction of the dolphin, Coryphaena hippurus, in the Straits of Florida. Copeia., 1967: 441-451. Beltran-Leon, B. y R. Rios. 2000. Estadios tempranos de peces del Pacífico colombiano. Instituto Nacional de Pesca y acuicultura INPA. Buenaventura. 727 pp. Bentivoglio, A.A. 1988. Investigations into growth, maturity, mortality rats and ocurrence of the dolphin (Coryphaena hippurus, Linnaeus) in the Gulf of mexico. M. Sc. Beverly, S., L. Chapman & W. Sokimi. 2003. Horizontal longline fishing methods and techniques: a manual for fishermen. Multipress, Noumea, New Zelanda. 132p. Bez, N. 2002. Global fish abundance estimation from regular sampling: the geostatistical transitive method. Can. J. Fish. Aquat. Sci., 59: 1921-1931. Botsford, L. W., J. C. Castilla & C. H. Peterson. 1997. The management of fisheries and marine ecosystems. Science., 277: 509-515. Brando, A., H. von Prahl y J.R. Cantera. 1992. Malpelo: isla oceánica de Colombia. Banco de Occidente, Santiago de Cali. 195 p. Brown-Peterson, N.J., J.S. Franks, D.M. Gibson & C. Marshall. 2014. Aspects of the reproductive biology of Yellowfin Tuna, Thunnus albacares, in the northern Gulf of Mexico. Proceedings of the Gulf and Caribbean Fisheries Institute., 66: 509-510. Bula-Meyer, G. 1985. Un núcleo nuevo de surgencia en el Caribe colombiano detectado en correlación con las macroalgas. Bol. Ecotrópica, 12: 3-25. Caldas, J.P. 2002. Ictiofauna acompañante de la pesca industrial con palangre horizontal de fondo en los bancos y bajos de la zona EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA norte del archipiélago de San Andrés, Providencia y Santa Catalina, caribe Colombiano. Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Santa Marta. 117 p. Cantera, J. y R. Contreras. 1993. Ecosistemas costeros. Colombia Pacífico. Fondo FEN Colombia, Bogotá. 65-79. Casanova, R., M. Zambrano, S. Latandret, N. Suarez, y C. Alban. 2012. Variabilidad de parámetros fisicoquímicos en una estación oceánica frente a la bahía de Tumaco. Bol. Cient. CIOH., 30: 105-116. Cayré, P., J.B. Amon Kothias, T. Diouf & J.M. Stretta J. 1993. Biology of tuna. En: Fonteneau. A y Marcille. J. (Eds). Resources, fishing and biology of the tropical tunas of the Eastern Central Atlantic. FAO Fisheries Document Paper. No. 292. Rome, FAO. 354p. Parameters for application in the yellowfin tuna stock assessment. WCPFCSC5-BI-WP-1. Cervigón, F. 1991. Los peces marinos de Venezuela. Vol I. Cromotip. 2ª Edición. Caracas, Venezuela. 425 p. Chase, R.E. 1968. Sea floor topography of the central eastern Pacific Ocean. U.S. Fish and Wildlife Service, BCF Circular 291. 33 p. Chuanmin, H., E. T. Montgomery, R. W Schmitt y F. E. Muller-Karger. 2004. The dispersalof the Amazon and Orinoco River water in the tropical and Caribbean Sea: Observations from space and S-PALACE floats. Deep-Sea Research II., 51: 1151-1171. CIAT (Inter-american Tropical Tuna Commission). 2015. Tunas, billfishes and other pelagic species in the eastern Pacific ocean in 2014. Fishery status report No. 13. La Jolla. 204 p. Clarke, T. A. 1973. Some aspects of the ecology of lanternfishes (Myctophidae) in the Pacific Ocean near Hawaii. Fish. Bull. U.S., 71: 401-434. Clarke, K. y R. Warwick. 2001. Change in marine communities: An approach to statistical analysis and interpretation. Second Edition. PRIMER-E. Plymounth. 172 p. Collette, B.B. 2002. Scombridae mackerels and tunas: 1836-1857. En Carpenter, K. (Ed.) The living marine resources of the western central Atlantic. Vol. 3. FAO, Roma, 1375-2127 p. Compagno, L.J.V. 2002. Sharks. In: Carpenter K.E. (ed.). The living marine resources of the Western Central Atlantic. Volume 1: Introduction, molluscs, crustaceans, hagfishes, sharks, batoid fishes, and chimaeras. FAO Species Identification Guide for Fishery Purposes and American Society of Ichth- 115 yologists and Herpetologists Special Publication No. 5.Rome, FAO. pp. 358-371. la pesquería de arrastre del mar Caribe de Colombia. Invest. Mar., 34(1): 23-42. Cressie, N.A.C. 1993. Statistics for spatial data. John. Wiley and Sons, New York. 900 p. Escarria, E., B.S. Beltrán-León y A. Giraldo. 2006. Ictioplancton superficial de la cuenca del océano Pacífico colombiano (septiembre 2003). Invest. Mar., 34(2): 169-173. Cury, P., L. Shannon & Y.J. Shin. 2003. The Functioning of Marine Ecosystems: a Fisheries Perspective. Chapter 7. Pp. 103-123. In: Sinclair, M. and Valdimarsson (eds.). Responsible fisheries in the marine ecosystem. 413 p. D’Elia, M., B. Patti, A. Sulli, G. Tranchida, A. Bonanno, G. Basilone, G. Giacalone, I. Fontana, S. Genovese, C. Guisande & S. Mazzola. 2009. Distribution and spatial structure of pelagic fish schools in relation to the nature of the seabed in the Sicily Straits (Central Mediterranean). Marine Ecology., 30 (1): 151-160. Díaz, J., C. Velasco y A. Ramírez. 1997. Monitoreo de la pesquería del camarón de aguas someras del Pacífico colombiano. INPA, Buenaventura, Colombia. 32 p. Díaz-Ochoa, J. & R.A. Quiñones. 2008. Relationship of precipitation, freshwater input, and sea level height with the abundance of the white shrimp (Litopenaeus occidentalis; Street, 1871) off Buenaventura, eastern tropical Pacific. Fish. Res., 92:148-161. Ditty, J., R. Shaw, C. Grimes, and J. Cope. 1994. Larval development, distribution, and abundance of common dolphin, Coryphaena hippurus, and pompano dolphin, C. equiselis (family: Coryphaenidae), in the northern Gulf of Mexico. Fish. Bull., 92(2): 275-291. Domokos, R., M. Seki, J. Polovina & D. Hawn. 2007. Oceanographic investigation of the American Samoa albacore (Thunnus alalunga) habitat and longline fishing grounds. Fisheries Oceanography., 16(6): 555-572. Drevillon, M., R. Bourdalle-Badie, C. Derval, J. M. Lellouche, E. Remy, B. Tranchant, M.Benkiran, E. Greiner, S. Guinehut, N. Verbrugge, G. Garric, C. E. Testut, M. Laborie,L. Nouel, P. Bahurel, C. Bricaud, L. Crosnier, E. Dombrowsky, E. Durand, N. Ferry, F. Fernandez, O. Le Galloudec, F. Messal & L. Parent. 2008. The GODAE/Mercator-Ocean global ocean forecasting system: results, applications and prospects, Journal of Operational Oceanography., 1:1, 51-57. Duarte, L. O., P. Gómez-Canchong, L. M. Manjarrés, C. B. García, F. D. Escobar, J. Altamar, J.E. Viaña, K. Tejada, J. Sánchez y F. Cuello. 2006. Variabilidad circadiana de la tasa de captura y la estructura de tallas en camarones e ictiofauna acompañante en Fahay, M. 2007. Early Stages of Fishes in the Western North Atlantic Ocean (Davis Strait, Southern Greenland and Flemish Cap to Cape Hatteras) Volume I y II. Northwestern Atlantic Fisheries Organization, Dartmouth, Nova Scotia. 1696 p. Fajardo, E. 1979. Surgencia costera en las proximidades de la península colombiana de La Guajira. Bol. Cient. CIOH. pp. 7-19. FAO. 1994. Examen de la situación mundial de las especies altamente migratorias y las poblaciones transzonales. FAO Documento Técnico de Pesca No. 337. Roma. FAO. 75p. FAO. 1995. Code of conduct for responsible fisheries. FAO, Roma. 41p. FAO. 1997. Technical Guidelines for Responsible Fisheries. No. 4. Roma, FAO. 82p. FAO. 2003. Fisheries management. 2. The ecosystem approach to fisheries. FAO Technical Guidelines for Responsible Fisheries . No.4. Suppl.2. FAO, Roma. 112p. FAO. 2012. El estado mundial de la pesca y acuicultura 2012. Roma. 231 p. Field, J.G., K.R. Clarke and R.M. Warwick. 1982. A practical strategy for analysing multispecies distribution patterns. Mar. Ecol. Prog. Ser., 8: 37–52. Figueiredo, M.B., A.G Santos, P. Travassos, C.M. Torres-Silva, F.H.V. Hazin, R. Coeli & B.R. Magalhães. 2008. Oocyte organization and ovary maduration of the bigeye tuna (Thunnus obesus) in the west tropical atlantic ocean. Collect. Vol. Sci. Pap. ICCAT, 62(2): 579-585. Foote, K.G., H.P. Knudsen, G. Vestnes, D.N. Maclennan and E.J. Simmonds. 1987. Calibration of acoustic instruments for fish density estimation: a practical guide. ICES. Cooperative Research Report N° 144. Franco-Herrera, A. 2005. Oceanografía de la ensenada de Gaira: El Rodadero, más que un centro turístico en el Caribe colombiano. Fundación Universidad de Bogotá Jorge Tadeo Lozano, Bogotá. 56 p. Fromentin, J.M., G. Reygondeau, S. Bonhommeau & G. Beaugrand. 2014. Oceanographic changes and explotation drive the spatio-temporal dynamics of Atlantic bluefin tuna (Thunnus thynnus). Fisheries Oceanography., 23(2): 147-156. Gallardo, A. 2006. Geostadística. Ecosistemas, 15(3): 48-58. 116 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Gartner T. L. Hopkins, R. C. Baird and D. M. Milliken. 1987. The lantern fishes (Pisces: Myctophidae) of the eastern Gu]f of Mexico. Fish. Bull. U.S., 85: 81-98. Herrera-Moreno, A. 2000. La clasificación numérica y su aplicación en la ecología. Instituto Tecnológico de Santo Domingo, República Dominicana, 88 p. Leggett, W.C. & E. Deblois. 1994. Recruitment in marine fishes: is it regulated by starvation and predation in the egg and larval stages?. Neth. J. Sea Res., 32(2): 119 134. Gianluca, V., T. Cristina, S. Rosalia, C. Simone, F. Vega, B. Emanuele, G. Philippe, M. Taberner, S. Pardo, J.M. L, C. R, 2014. Product user manual For theOCEANCOLOUR_GLO_ CHL_L4_NRT_OBSERVATIONS_009_033. MyOcean, [Online] Available at: http:// catalogue.myocean.eu.org/static/ resources/myocean/pum/MYO2-OC-PUM009-ALL-V4.2.pdf [Accessed February 18th 2015]. Hindson, J., D.D. Hoggarth, M. Krishna, C.C. Mees & C. O’Neill. 2005. How to manage a fishery: a simple guide to writing a fishery management plan. Marine Resources Assessment Group, London. 81p. Lehoday. P, and B. Leroy. 1999. Age and growth of yellowfin tuna (Thunnus albacares) form the western and central Pacific ocean as indicated by daily growth increments and tagging data. SCTB12 working paper. YFT-2. 12th standing committee on tuna and billfish. Gómez-León, J. 2005. Estudio epizoótico de agentes patógenos en almeja fina (Ruditapes decussatus) Cultivada en Galicia. Tesis Doctorado Cienc. Biol., Univ. de Santiago de Compostela, España. 220 p. Gómez, C. y I. Martínez. 2005. Nueva productividad biológica en el Pacífico Oriental Ecuatorial: Respuesta al aporte de nutrientes por los ríos del Pacífico colombiano, surgencia costera y polvo atmosférico. Boletín de Geología Vol. 27, No. 2. Grijalba-Bendeck, M., D. Bustos-Montes, C. Posada-Peláez y A. Santafé-Muñoz (Ed.). 2012. La Pesca artesanal marítima del departamento del Magdalena: una visión desde cuatro componentes. Fundación Universidad de Bogotá Jorge Tadeo Lozano, Ministerio de la Agricultura y Desarrollo Rural, Proyecto Transición de la Agricultura, Bogotá, Colombia. 454 p. Gutiérrez, N., R. Hilborn & O. Defeo. 2011. Leadership, social capital and incentives promote successful fisheries. Nature., 470: 386-389. Haggarty, D. R., and J.R King. 2006. CPUE as an index of relative abundance for nearshore reef fishes. Fish. Res., 81(1): 89-93. Haimovici, M., and A.O. Ávila Da Silva. 2007. A comparison of methods for estimating relative abundances in bottom longline fishery surveys. Fish. Res., 85: 226-231. Hastie, T.J., and R.J. Tibshirani. 1990. Generalized Additive Models. Chapman and Hall,London. 344 p. Harley, S. J., R.A Myers & A. Dunn. 2001. Is catchper-unit-effort proportional to abundance?. Canadian Journal of Fisheries and Aquatic Sciences., 58(9): 1760-1772. Herazo, D., A. Torres y E. Olsen. 2006. Análisis de la composición y abundancia de la ictiofauna presente en la pesca del camarón rosado (Penaeus notialis) en el golfo de Morrosquillo, Caribe colombiano. Revista MVZ Córdoba., 11(2): 47-61. Hilborn, R. 2006. Defining success in fisheries and conflicts in objectives. Marine Policy., 31: 153-158. Holden, M.J. y D.F.S. Raitt. 1974. Manual de ciencias pesqueras. Parte 2. Métodos para investigar los recursos pesqueros y su aplicación. FAO. Doc. Tec. Pesca (115), 211 p. Hoyle, S. D., S. Nicol, and D. G. Itano. 2009. Revised biological parameters for application in the yellowfin tuna stock assessment. WCPFCSC5-BI-WP-1. Isaza-Toro, E. 2013. Comportamiento espacial de variables químicas en la Cuenca Pacífica Colombiana durante el Crucero Oceanográfico Cuenca Pacífica Colombiana –CPC LI. Bol. Cient. CIOH., 31: 95-108. Leis, J.M. 1993. Larval fish assemblages near Indo-Pacific coral reefs. Bull. Mar. Sci., 53(2): 362-392. Leis, J. M. 2010. Ontogeny of behaviour in larvae of marine demersal fishes. Ichthyol. Res., 57(4): 325-342. Lessa, R. & P. Duarte-Neto. 2004. Age and growth of yellowfin tuna (Thunnus albacares) in the western equatorial Atlantic, using dorsal fin spines. Fish. Res., 69:157-170. Maguire, J.J., M. Sissenwine, J. Csirke, R. Grainger, & S. Garcia. 2006. The state of world highly migratory, straddling and other high seas fishery resources and associated species. FAO Fisheries Technical Paper. No. 495. Rome. FAO. 84p. Magurran, A. E. 2013. Ecological diversity and its measurement. Springer Science & Business Media. Itano, D.G. 2000. The reproductive biology of yellowfin tuna (Thunnus albacares) in Hawaiian waters and the western tropical Pacific Ocean: project summary. SOEST 00-01, JIMAR. Contribution 00-328. 75 p. Online: http://www.soest.hawaii.edu/ PFRP/biology/itano/itano_yft.pdf. Mahon, R., L. Fanning & P. McConney. 2009. A governance perspective on the large marine ecosystem approach. Marine Policy., 33: 317-321. Jiménez, S. 2008. Asociaciones de larvas de peces por estadio de desarrollo en la costa noroccidental de la península de Baja California. Tesis de Doctorado. Centro Interdisciplinario de Ciencias Marinas. Instituto Politécnico Nacional. 223 p. Manjarrés-Martínez, L.M., J.C. Gutiérrez-Estrada, J. Mazenet-González & M.C. Soriguer. 2010. Seasonal patterns of three fish species in a Caribbean coastal gill-net fishery: Biologically induced or climate-related aggregations?. Fish. Res., 106(3): 358-367. Kikkawa, B., and J.W. Cushing. 2002. Growth of yellowfin tuna (Thunnus albacares) in the equatorial western pacific ocean. Working paper YFT-4. 15th meeting of the standing committee on tuna and billfishes, SCTB 15, Honolulu, Hawai, 22-27 july 2002. 12 p.p. Martínez-Aguilar, T.I., A. Giraldo y E. Rodríguez-Rubio. 2010. Ictioplancton en la zona costera del Pacífico colombiano durante la fase terminal de El Niño 2006-2007. Lat. Am. J. Aquat. Res., 38(1): 151-166. King, M. 2013. Fisheries biology, assessment and management. John Wiley & Sons. Langley, A., K. Briand, D. Séan-Kirby & R. Murtugudde. 2009. Influence of oceanographic variability on recruitment of yellowfin tuna (Thunnus albacares) in the western and central Pacific Ocean. Can. J. Fish. Aquat. Sci. 66: 1462-1477. Lasso, J. & L.A. Zapata. 1999. Fisheries and biology of Coryphaena hippurus (Pisces: Coryphaenidae) in the Pacific coast of Colombia and Panama. Sci. Mar., 63(3-4): 387-399. Maunder, M.N. & A.E. Punt. 2004. Standardizing catch and effort data: a review of recent approaches. Fish. Res., 70: 141-159. Medellín-Mora, J., F. Polanco, S. Navas y R. Gabriel. 2013. Inventario de larvas de peces registradas para el Caribe colombiano. Bol. Invest. Mar. Cost., 42(2): 233-253. Medina, J.A. 2002. Ensamblaje de peces demersales explotados por la flota industrial camaronera en la plataforma continental de La Guajira (Caribe colombiano). Trabajo de Grado (Biólogo Marino). Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Santa Marta. 77 p. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Melo, G., L. Maldonado y L.A. Zapata. 2011. Aspectos generales de la pesquería de atún en Colombia. En: Díaz, J.M., C. Vieira y G. Melo (Eds.). Diagnóstico de las principales pesquerías del Pacífico colombiano. Fundación Marviva – Colombia, Bogotá, pp. 195-213. Miller, B.S. & A.W Kendall. 2009. Early life history of marine fishes. University of California Press, Berkeley. 364 p. Molinari, R.L., D.K. Atwood, C. Duckett, M. Spillane & I. Brooks. 1980. Surface currents in the Caribbean Sea as deduced from satellite tracked drifting buoys. In Proceedings of the Gulf and Caribbean Fisheries Institute, (31): 106-115. Moser, H. G. 1996. The early stages of fishes in the California Current region. California Cooperative Oceanic Fisheries Investigations Atlas., 33: 1-1505. Moser, H.G., R.L. Charter, P.E. Smith, D.A. Ambrose, W. Watson, S.R. Charter & E.M. Sandknop. 2002. Distributional atlas of fish larvae and eggs from Manta (surface) samples collected on CalCOFI surveys from 1977 to 2000. CalCOFI Rep. Atlas No. 35. 97 p. Ortega-García, S., Zuñiga Florea, M. y Rodríguez-Sánchez, R. 2011. Revisión del estado del conocimiento del dorado (Coryphaena hippurus) en el Pacífico Mexicano. El Vigia, 16 (39). Pp 33-35. Oxenford, H. 1999. Biology of the dolphinfish (Coryphaena hippurus) in the western Central Atlantic: a review. In Biology and fishery of dolphinfish and related species. Scientia Marina. Barcelona-España. 63 (3-4): 277-301 pp. Pabón, L. 2013. Composición, densidad y distribución del ictioplancton en aguas oceánicas del mar Caribe colombiano y su relación con algunas variables oceanográficas. Tesis de pregrado Univ. Jorge Tadeo Lozano. Santa Marta. 123 p. Páramo, J., R. Quiñones., A. Ramirez y R. Wiff. 2003. Relationship between abundance of small pelagic fishes and environmental factors in the Colombian Caribbean Sea: an analysis based on hydroacoustic information. Aquat. Liv.Res., 16: 239-245. Páramo, J., Guillot-Illidge, L., Benavides, S., Rodríguez, A., y Sánchez-Ramírez, C. 2009. Aspectos poblacionales y ecológicos de peces demersales de la zona norte del Caribe colombiano en relación con el hábitat: una herramienta para identificar Áreas Marinas Protegidas (AMP) para el manejo pesquero. Caldasia, 31(1): 123-144. Páramo, J., C. Marco y S. Núñez. 2011. Evidencias de desacople físico-biológico en el sistema 117 de surgencia en La Guajira, Caribe colombiano. Rev. Biol. Mar. y oceanog., 46(3): 421-430. A.; M. Vasconcellos y Y. Yimin. FAO Documento Técnico de Pesca y Acuicultura. No. 520. FAO, Rome, Italy . 169pp. Parra, R. S. 1977. Resultados oceanográficos del Pacífico colombiano durante el año de 1976. Boletín Científico Armada Nacional., 1(1): 41p. Ramírez, D., y A. Giraldo. 2006. Estructura comunitaria del fitoplancton de la Cuenca Pacífica Colombiana durante la campaña oceanográfica Pacífico XXXIX-Erfen XXXVII. Boletín Científico CCCP., 13: 65-84. Perez, R.N. & Y. Sadovy. 1991. Preliminary data on landings records and reproductive biology of Coryphaena hippurus L in Puerto Rico. Proc. Gulf Caribb. Fish. Inst., 44: 636-650. Perez R.N., A.M. Roman and G.A. Rivera. 1992. Investigation of the reproductive dynamics and preliminary evaluation of landings data of the dolphinfish Coryphaena hippurus L. Final Report for Dingell-Jhonson project F26-1. Puerto Rico Department of natural Resourses Fishery Research laboratory, Mayaguez, P.R. 95 pp. Perry, R.I., C.J. Walters and J.A. Boutillier. 1999. A framework for scientific advice for the management of new and developing invertebrate fisheries. Rev. Fish. Biol. Fisheries., 9: 125-150. Petersen, C.W. & R.R. Warner. 2002. The ecological context of reproductive behavior. Pp. 103-118. En: Sale, P.F. (Ed.). Coral reef fishes: dynamics and diversity in a complex ecosystem. Academic Press, San Diego. Polanco F., A., A. Acero P. y M. Garrido. 2010. Aportes a la biodiversidad íctica del Caribe colombiano. 316-353. En navas, G.R., C. Segura-Quintero, M. Garrido-Linares, M. Benavides-Serrato y D. Alonso (eds.). Biodiversidad del margen continental del Caribe Colombiano, Serie de Publicaciones Especiales No.20, Invemar, Santa marta.458 p. Poveda, G., W. Rojas, M.L. Quiñones, I.D. Vélez, R.I. Mantilla, D. Ruiz & G.L. Rua. 2001. Coupling between annual and ENSO timescales in the malaria-climate association in Colombia. Environ. Health persp., 109(5): 489. Prahl, H.von. 1990. Malpelo la roca viviente. Editorial Presencia, FEN, Bogotá.105 p. Prathibha. R., & R. Mohan. 2009. Fishery and Biological Aspects of Yellowfin Tuna Thunnus albacares along Andhra Coast, India. Asian Fisheries Science., 22(1): 235-244. Prahitbha. R, Syda. G and K. Rammohan. 2012. Age. Growth and population structure of the yellowfin tuna Thunnus albacares (Bonnaterre, 1788) exploited along the east coast of India. Indian J. Fish., 59(1): 1-6. Purcell, S.W. 2010. Manejo de las pesquerías de pepino de mar con un enfoque ecosistémico. Editado/compilado por Lovatelli, R Development Core Team. 2015. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org/. Rivoirard, J., J. Simmonds, K.G. Foote, P. Fernandes & N. Bez. 2000. Geostatistic for estimating fish abundance. Blackwell Science Ltd. London. 205p. Rodríguez-Rubio, E. and J. Stuardo. 2002. Variability of photosynthetic pigments in the Colombian Pacific Ocean and its relationship with the wind field using ADEOS-I data, Proc. Indian. Acad. Sci. Earth Planet. Sci., 111(3): 227-236. Rodríguez-Rubio, E., W. Schneider & R. Abarca del Rio. 2003. On the seasonal circulation within Panama Bight derived from satellite observations of winds, altimetry and sea surface temperature. Geophys. Res. Lett., 30: 1410-1413. Rodríguez, A. y J. Páramo. 2012. Distribución espacial del pargo rayado Lutjanus synagris (Pisces: Lutjanidae) y su relación con las variables ambientales en el Caribe colombiano. Actual Biol., 34(96): 55-66. Rodríguez, A., M. Rueda, J. Viaña, C. García, F. Rico, L. García, A. Girón. 2012. Evaluación y manejo de la pesquería de camarón de aguas profundas colombiano 2010-2012. Invemar, Colciencias, Incoder. Serie de publicaciones generales del Invemar No. 56. Santa Marta. 114p. Rodríguez, A., M. Rueda, D. Bustos-Montes, A. Galeano, B. Mejía-Mercado, J. Viaña, G. Ramírez, J. Correa, E. Isaza, A. Girón, E. Escarria, S. Salas, L. Prieto, G. Angulo, H. Castillo y J. Sierra. 2015. Programa de investigación pesquera en aguas marinas jurisdiccionales de Colombia. Convenio Aunap-Invemar No. 0140-2014. Informe técnico y financiero final. PRY-VAR-00214-ITF. Santa Marta. 287p+Anexos. Rossignol, M. 1968. Le thon â nageoires jaunes de l’Atlantique Thunnus (Neothunnus) albacores. (Bonnaterre, 1788). Mém. ORSTOM Paris, (25):11 p. Rubio, E.A. 1986. Lista sistemática de peces costeros y de profundidad del Pacífico colombiano. Centro de Publicaciones. 118 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA Facultad de Ciencias, Univ. Valle, Cali. 258 p. biano. Ed. Editorial Universidad Nacional de Colombia. ISBN: 978-958-719-834-8. Rubio, E.A., A. Suarez, F. Estupiñan, W. Henao y B. Vargas. 1992. Los recursos ictiológicos de la Isla de Malpelo (Colombia). I. Una revisión de su conocimiento y nuevos aportes para la íctiofauna de la isla., Mem. VIII Semin. Nal. Cien. Tecnol. Mar, CCO, Bogotá, 642-658. Silva, C. 2005. Asociaciones espacio-temporales de larvas de peces y su relación con las condiciones ambientales de las costas de Jalisco y Colima, México. Tesis de Maestría. Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas. 60 p. Rueda, M., A. Rodríguez, J. Gómez-León, J. López, G. Ospina, M. Santos-Acevedo, D. Bustos, J. Viaña, A. Galeano, A. Girón, M. Bastidas, C. Ricaurte, D. Romero, J. Correa y G. Angulo. 2014. Prospección pesquera en el Caribe continental y océano Pacífico de recursos pelágicos y aportes a la maricultura de pepino de mar en el Caribe colombiano (Etapa I-2013). Convenio No. 0038-2013 Aunap e Invemar. Informe Final. Santa Marta. 211 p. Rueda, M., O. Doncel, E.A. Viloria, D. Mármol, C. García, A. Girón, L. García, F. Rico. A. Rodríguez, C. Borda, C. Barreto. 2012. Atlas de la pesca marino - costera de Colombia: 2010 – 2011. Invemar, ANH e Incoder. Serie de publicaciones del Invemar. Santa Marta. Tomo Pacífico 98 p. Ruiz-Ochoa, M. A. 2011. Variabilidad de la cuenca Colombia (mar Caribe) asociada con El Nino-Oscilación del Sur, vientos Alisios y procesos locales. Tesis Doctor en Ingeniería y Recursos Hídricos., Universidad Nacional, Medellín. 202 p. Russell, B. 2002. Sphyraenidae: Barracuda. Pp. 1807-1809. En: Carpenter, K.E. (Ed.). The living marine resources of the western central Atlantic. Bony fishes part 2. FAO Species identification guide for fishery purposes and American society of ichthyologists and herpetologists special publication. Rome, 3(5): 1375-2127. Saborido-Rey, F. 2004. Ecología de la reproducción y potencial reproductivo en las poblaciones de peces marinos. Universidad de Vigo, España. 69p. Sánchez, L., M.F. Lavín, M. Peguero, C.A. León-Chávez, F. Contreras-Catala, S.G. Marinone, I.V. Gutiérrez-Palacios and V.M. Godínez. 2009. Seasonal changes in larval fish assemblages in a semi-enclosed sea (Gulf of California). Cont. Shelf Res., 29: 1697-1710. Schaefer, M.B. 1998. Reproductive biology of the yellowfin tuna (Thunnus albacares) in the eastern Pacific ocean. Inter-Am Trop Tuna Comm Bull 21: 205-272. Selvaraj, J., A.I. Guzmán y A. Martínez. 2011. Guía para identificación de áreas de pesca para grandes pelágicos en el Pacífico colom- Simmonds, E.J., N.J. Williamson, F. Gerlotto and A. Aglen. 1992. Acoustic survey design and analysis procedure: A comprehensive review of current practice. En: ICES Cooperative Research Report: Rapport des Recherches Collectives. No. 187: 112 p. Simmonds, J. & D. MacLennan. 2005. Fisheries acoustics: theory and practice. Blackwell Science, Oxford. 438 p. Simrad. 2003. Simrad EK60. Scientific echosounder application. Operator manual. 90 p. Smith, P. & S. Richardson. 1979. Standard techniques for pelagic fish egg and larva surveys. FAO fish. Tech. Pap. (175): 100. Solano, O.D. y H. Hernández. 1998. Comunidades ícticas de la isla Malpelo (Pacífico colombiano) y anotaciones sobre estudios marinos en la isla. Informe de proyecto, Invemar, Santa Marta. 7 p. Solano-Sare, A. Tresierra-Aguilar, V. García-Nolasco, T. Dioses, W. Marín, C. Sánchez y C. Wosnitza-Mendo. 2008. Biología y pesquería del perico. Instituto del Mar del Perú. 23 p. Steele, J. & P. Hoagland. 2003. Are fisheries “sustainable”?. Fisheries Research., 64: 1-3. Stewart, R. H. 2008. Introduction to physical oceanography. Department of Oceanography, Texas A & M University., College Station, Texas. 345p. Sun, C., S. Nana-Jay, and Y. Su-Zan. 2003. Estimation of growth parameters and age composition for yellowfin tuna, Thunnus albacares, in the western Pacific using the length-based Multifan method. SCTB16 working paper, Standing committee on tuna and billfish, Qld, Australia 9-16 july 2003. Dirección General Marítima - Centro Control Contaminación del Pacífico. San Andrés de Tumaco, Colombia. (7): 7-19. Tejada, C., L. Castro. A. Navarrete. T. Cardona, L. Otero, F. Afanador, A. Mogollón y W. Pedroza. 2003. Panorama de la contaminación marina del Pacífico colombiano. Centro Control Contaminación del Pacífico colombiano. Ed. Dimar. Serie Publicaciones Especiales Vol. 3, San Andrés de Tumaco, 120 pp. Thurow, F. 1997. Estimation of the total fish biomass in the Baltic Sea during the 20th century. ICES Journal of Marine Science., 54: 444-461. Vergara A., E. Arteaga y J. Rodríguez. 1999. Composición, abundancia y distribución del ictioplancton del Caribe colombiano. Bol. Cient. INPA., 6: 77-83. Ward, J.M., J.E. Kirkley, R. Metzner, & S. Pascoe. 2004. Measuring and assessing capacity in fisheries. 1. Basic concepts and management options. FAO Fisheries Technical Paper. No. 433/1. Rome, FAO. 40p. Wild, A. 1986. Growth of yellowfin tuna, Thunnus albacares, in the eastern Pacific Ocean based on otolith increments. Inter-American Tropical Tuna Commission Bulletin 18:421-482. Wyrtki, K. 1965. Corrientes superficiales del Océano Pacífico Oriental Tropical. Comisión Interamericana del Atún Tropical. Boletin IX (5). Wood, S. 2006. Generalized Additive Models: An introduction with R. Chapman and Hall/ CRC. New York. 385 p. Wooster, W. S. 1959. Oceanographic observations in the Panama Bight, “Askoy” Expedition, 1941. Bulletin of the American Museum of Natural History, 118(3): 113-152. Worm, B., R. Hilborn, J.K. Baum, T.A. Branch, J.S. Collie, C. Costello & D. Zeller. 2009. Rebuilding global fisheries. Science, 325(5940): 578-585. Sun, C., W. Wang, and S. Yeh. 2005. Reproductive biology of yellowfin tuna in the central and western Pacific Ocean. 1st meeting of the Scientific Committee of the Western and Central Pacific Fisheries Commission WCPFC-SC1. Noumea, New Caledonia, August 8-19, 2005. 15 p. Zapata, L.A., J. Herrera, S. Beltran-León, P. Jiménez, L. Prieto, C. Guevara-Fletcher y E. Zambrano. 2011. Estado actual de la pesquería de pequeños pelágicos en el Pacífico colombiano. En: J.M. Díaz, C. Vieira y G. Melo (eds.). Diagnóstico de las principales pesquerías del Pacífico colombiano. Fundación Marviva – Colombia, Bogotá. PP. 157-172. Tchantsev, V. y E. Cabrera-Luna. 1998. Algunos aspectos de investigación de la formación del régimen oceanográfico en el Pacífico colombiano. Boletín Científico CCCP. Zudaire, H. Murua, M. Grande and N. Bodin. 2013. Reproductive potential of Yellowfin Tuna (Thunnus albacares) in the western Indian Ocean. Fish. Bull., 111: 252–264. EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 119 SERIE DE PUBLICACIONES GENERALES DEL INVEMAR Sin número. Referencias bibliográficas publicadas e inéditas de la Ciénaga Grande de Santa Marta, Caribe colombiano. Volumen I. Sin número. Referencias bibliográficas publicadas e inéditas de la Ciénaga Grande de Santa Marta, Caribe colombiano. Volumen II. 1. Programa Nacional de Investigación en Biodiversidad Marinas y Costera (PNIBM) 2. Política nacional ambiental para el desarrollo sostenible de los espacios oceánicos y las zonas costeras e insulares de Colombia 3. Informe del estado de los ambientes marinos y costeros en Colombia: 2000 4. Ojo con Gorgona. Parque Nacional Natural 5. Libro rojo de peces marinos de Colombia 6. Libro rojo de invertebrados marinos de Colombia 7. Las aguas de mi Ciénaga Grande. Descripciones de las condiciones ambientales de la Ciénaga Grande de Santa Marta 8. No asignado 9. Guía práctica para el cultivo de bivalvos marinos del Caribe colombiano: Madreperla, ostra alada, concha de nácar y ostiones 10.Aproximación al estado actual de la bioprospección en Colombia 11.Plan nacional en bioprospección continental y marina 12.Conceptos y guía metodológica para el manejo integrado de zonas costeras en Colombia, Manual 1: Preparación, caracterización y diagnóstico 13.Manual de técnicas analíticas para la determinación de parámetros fisicoquímicos y contaminantes marinos: aguas, sedimentos y organismos 14.Una visión de pesca multiespecífica en el Pacífico colombiano: adaptaciones tecnológicas 15.Amenazas naturales y antrópicas en las zonas costeras colombianas 16.Atlas de paisajes costeros de Colombia 17.Atlas de la calidad de las aguas marinas y costeras de Colombia 18.Manual del Sistema de Información Pesquera del Invemar: una herramienta para el diseño de sistemas de manejo pesquero 19.Bacterias marinas nativas: degradadoras de compuestos orgánicos persistentes en Colombia 20.Política Nacional del Océano y los Espacios Costeros (PNOEC) 21.Manual metodológico sobre el monitoreo de los manglares del Valle del Cauca y fauna asociada, con énfasis en aves y especies de importancia económica (piangua y cangrejo azul) 22.Lineamientos y estrategias de manejo de la Unidad Ambiental Costera (UAC) del Darién 23.Plan de Manejo Integrado de la Zona Costera-UAC Llanura Aluvial del Sur, Pacífico colombiano 24.Cartilla lineamientos y estrategias para el manejo integrado de la UAC del Darién, Caribe colombiano Sin número. Prioridades de conservación in situ para la biodiversidad marina y costera de la plataforma continental del Caribe y Pacífico colombiano 25.Cartilla etapas para un cultivo de bivalvos marinos (pectínidos y ostras) en sistema suspendido en el Caribe colombiano 26.Programa Nacional de Investigación para la Prevención, Mitigación y Control de la Erosión Costera en Colombia (PNIEC) 27.Modelo de uso ecoturístico de la bahía de Neguanje Parque Nacional Natural Tayrona 28.Criadero de postlarvas de pectínidos de interés comercial en el Caribe colombiano 29.Viabilidad de una red de áreas marinas protegidas en el Caribe colombiano 30.Ordenamiento ambiental de los manglares del Archipiélago San Andrés, Providencia y Santa Catalina, Caribe colombiano 31.Ordenamiento ambiental de los manglares en La Guajira 32.Ordenamiento Ambiental de los manglares del municipio de Timbiquí, Cauca (Pacífico colombiano) 33.Ordenamiento Ambiental de los manglares del municipio de Guapi, Cauca 34.Ordenamiento Ambiental de los manglares del municipio de López de Micay, Cauca 35.Avances en el manejo integrado de zonas costeras en el departamento del Cauca 120 EVALUACIÓN DIRECTA DE LAS POBLACIONES DE PECES GRANDES PELÁGICOS DEL PACÍFICO Y CARIBE CONTINENTAL DE COLOMBIA 36.Ordenamiento ambiental de los manglares de la Alta, Media y Baja Guajira 37.Aprendiendo a conocer y cuidar el agua en la zona costera del Cauca 38.Guía de bienes y servicios del Old Point Regional Mangrove Park 39.Aves del estuario del río Sinú 40.Cultivo de pectínidos en el Caribe colombiano 41.Informe técnico. Planificación ecorregional para la conservación in situ de la biodiversidad marina y costera en el Caribe y Pacífico continental colombiano 42.Guía para el reconocimiento de corales escleractinios juveniles en el Caribe 43.Viabilidad socioeconómica del establecimiento de un AMP: la capacidad adaptativa de la comunidad de Nuquí (Chocó) 44.Guía metodológica para el manejo integrado de zonas costeras en Colombia. Manual 2: Desarrollo etapas I y I 45.Pianguando: Estrategias para el manejo de la piangua (CD) 45.Pianguando: Estrategias para el manejo de la piangua (cartilla) 46.Avances en la reproducción y mantenimiento de peces marinos ornamentales 47.Contribución a la biología y mantenimiento de peces marinos ornamentales 48.Estrategia para el fortalecimiento del Sistema de Indicadores Ambientales Marinos y Costeros de Colombia (Proyecto Spincam Colombia) 49.Lineamientos de manejo para la Unidad Ambiental Costera Estuarina río Sinú, Golfo de Morrosquillo, sector Córdoba 50.Guía municipal para la incorporación de determinantes ambientales de zona costera en los planes de ordenamiento territorial municipios de San Antero y San Bernardo del Viento 51.Manual para la pesca artesanal responsable de camarón en Colombia: adaptación de la red Suripera 52.Cuidando la calidad de las aguas marinas y costeras en el departamento de Nariño 53.Lineamientos de manejo para la UAC Estuarina Río Sinú-Golfo de Morrosquillo, sector Córdoba 54.Propuesta de estandarización de los levantamientos geomorfológicos en la zona costera del Caribe colombiano 54.Área de Régimen Común Colombia-Jamaica: un reino, dos soberanos 55.Lineamientos de adaptación al cambio climático para Cartagena de Indias 56.Evaluación y manejo de la pesquería de camarón de aguas profundas en el Pacífico colombiano 2010-2012 57.Gestión costera como respuesta al ascenso del nivel del mar. Guía para administradores de la zona costera del Caribe 58.Articulación del Subsistema de Áreas Marinas Protegidas al Sistema Regional de Áreas Protegidas del Caribe Colombiano 59.Bases de la investigación pesquera participativa para la construcción de acuerdos de pesca responsable con mallas en el Distrito de Manejo Integrado Bahía de Cispatá 60.Articulación del Subsistema de Áreas Marinas Protegidas (SAMP) al plan de acción del Sirap Pacífico 61.Guía metodológica para el manejo integrado de zonas costeras en Colombia. Manual 3: Gobernanza 62.Integración de la adaptación al cambio climático en la planificación territorial y gestión sectorial de Cartagena de Indias 63.Plan 4C Cartagena de Indias competitiva y compatible con el clima 64.Lineamientos de adaptación al cambio climático del área insular del distrito de Cartagena de Indias 65.Adaptación al cambio climático en ciudades costeras de Colombia. Guía para la formulación de planes de adaptación 66.Protocolo Indicador Condición Tendencia Áreas Coralinas (ICTAC) 67.Protocolo Indicador Condición Tendencia Bosques de Manglar (ICTBM) 68.Protocolo Indicador Condición Tendencia Pradera de Pastos Marinos (ICTPM) 69.Protocolo Indicador Calidad Ambiental de Agua (ICAMPFF) 70.Protocolo Indicador Densidad poblacional de pez león (Pterois volitans) 71.Protocolo Indicador Riqueza de aves acuáticas 72.Protocolo Indicador Uso de recursos hidrobiológicos 73.Protocolo Indicador Variación línea de costa: Perfiles de playa 74.Lineamientos del plan de ordenamiento y manejo de la Unidad Ambiental Costera (Pomiuac) río Magdalena, completo Canal del Dique-sistema lagunar Ciénaga Grande de Santa Marta, sector zona costera del departamento de Bolívar 75.Lineamientos para el plan de ordenación y manejo integrado de la Unidad Ambiental Costera (UAC) río Magdalena, complejo Canal del Dique-sistema lagunar Ciénaga Grande de Santa Marta, sector zona costera del departamento de Bolívar. Cartilla 76.Vulnerabilidad de la población costera frente a la contaminación orgánica y microbiológica en la bahía de Buenaventura 77.Plan 4C: Cartagena de Indias competitiva y compatible con el clima. Resumen ejecutivo. 78.Documento de conceptualización del sistema de monitoreo del Subsistema de Áreas Marinas Protegidas en Colombia. 79.Portafolio: “Áreas de arrecifes de coral, pastos marinos, playas de arena y manglares con potencial de restauración en Colombia” 80.Conceptualización del Subsistema de Áreas Marinas Protegidas en Colombia. 81.Tortuga Verde 82.Bosques de Vida: Manglares de Mi Guajira 83.Exploradores del Océano. Cartilla de Actividades. 84.Indicadores en la Unidad de Manejo Integrado Guapi-Iscuandé: caso piloto de implementación de indicadores de manejo integrado de la zona costera a escala local. 85.Guía Metodológica para el desarrollo de proyectos tipo REDD+ en ecosistemas de manglar. 86.Entendiendo las decisiones de producción y consumo de los hogares de pescadores: Una aproximación a través de los modelos de producción de hogares. 00 0 -3 0 00 00 -30 00 -3 - 300 0 -3000 0 00 -3 - 30 -30 00 - 30 0 0 0 -300 -30 00 -3 -3000 0 -3 00 0 00 0 -3 0 0 -3000 -300 0 -3000 -3000 -3000 00 00 0 00 -3 -3 0 - 30 0 00 0 0 -3 -3 -30 00 -3 0 00 0 -3 -3 0 0 00 00 -3 -3 0 0 0 -300 00 0 0 0 -3 00 0 0 -3 0 0 0 -300 0 -3000 -3 0 00 0 -3 0 00 00 0 -3 -3 00 40 00 -3 0 -3000 -3 00 00 0 00 - 30 0 0 -3000 00 - 40 00 -300 -3 3 00 00 0 0 -3 -3000 0 -3000 -300 0 - 3 0 00 00 -3 00 00 -30 00 -3000 - -3 0 - 30 0 0 -30 -30 00 00 00 -3 00 -3000 0 -30 0 0 -30 -30 -30 -30 00 -3 0 0 -3 0 00 0 0 - -40 00 - 30 -30 00 0 - 30 - 3 00 0 00 0 0 -3 00 00 0 00 0 -3 00 -3 0 -30 0 -3000 00 0 - 30 -3000 -30 00 0 00 -3 00 0 -300 0 0 30 00 00 -300 -3 -3 -3000 -3000 -30 -3 0 - 00 00 -30 00 -3000 -30 00 0 40 0 0 - -3 00 00 -30 -3 0 -300 -3000 -3000 0 - 3 00 - 3 00 -30 00 0 -3 0 00 -30 -3 00 00 - 3 00 0 0 -3 0 00 -3000 - 3 00 -30 00 -3000 -3000 - 30 0 00 -3 00 0 00 0 00 -3000 -30 -3 -3000 00 -3 0 00 -300 0 -3000 30 -3 0 0 0
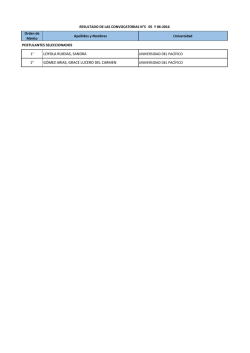



© Copyright 2026