
1.Memorias_Primer encuentro de bosques Ecuador 08
PRIMER ENCUENTRO NACIONAL DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Memorias del Evento PROGRAMA NACIONAL DE FORESTERÍA ESTACIÓN EXPERIMENTAL SANTA CATALINA INSTITUTO NACIONAL DE INVESTIGACIONES AGROPECUARIAS Quito, junio de 2015 Editores: Grijalva Olmedo Jorge, Ramos Veintimilla Raúl, Vera Vélez Roy; Barrera Aguilar Paulo; Sigcha Morales Franklin. Comité Técnico Comité de Publicaciones de la Estación Experimental Santa Catalina: Ing. David Galarza Ing. Iván Reinoso Ing. Ricardo Limongi Instituto Nacional de Investigaciones Agropecuarias Programa Nacional de Forestería Panamericana Sur. Km 1. Sector Cutuglahua, Quito-Ecuador Teléfono: (593) 269 0692 Edición electrónica localizable en las páginas: www.bosquesyagroforesteriainiap.com www.iniap.gob.ec Forma de citar este documento: Grijalva, J.; R. Ramos; R. Vera; P. Barrera y F. Sigcha (eds). 2013. Primer Encuentro Nacional de Bosques, Recursos Genéticos Forestales y Agroforestería. Memorias del Evento. Instituto Nacional de Investigaciones Agropecuarias. Quito, Ecuador. 318 p. Publicación Miscelanea No. 425 Registro ISBN: 978-9942-13-642-8 ÍNDICE PRESENTACIÓN........................................................................................................................................................... 6 INTRODUCCIÓN ........................................................................................................................................................... 7 CONFERENCIAS MAGISTRALES ............................................................................................................................... 8 RECURSOS GENÉTICOS FORESTALES, ALTERNATIVAS DE USO Y CONSERVACIÓN EN EL MUNDO CONTEMPORÁNEO ......................................................................................................................................................... 9 EL INVENTARIO FORESTAL NACIONAL DE ESPAÑA: HERRAMIENTA PARA LA INVESTIGACIÓN Y LA GESTIÓN FORESTAL ..................................................................................................................................................................... 16 PLANTACIONES FORESTALES MIXTAS: UNA OPORTUNIDAD PARA MEJORAR LA PRODUCTIVIDAD Y LA RESILIENCIA .................................................................................................................................................................. 21 PRODUCTOS FORESTALES NO MADEREROS (PFNM)….UN APORTE AL MANEJO FORESTAL SOSTENIBLE .. 25 DOMESTICACIÓN DE ÁRBOLES AGROFORESTALES: UN REPASO ANALÍTICO.................................................... 30 PATRONES Y DETERMINANTES HISTÓRICOS Y GEOGRÁFICOS DE LA DEFORESTACIÓN EN EL ECUADOR, 1950-2010 Y TENDENCIAS AL FUTURO. ..................................................................................................................... 35 BOSQUES Y CAMBIOS CLIMÁTICOS ...................................................................................................................... 41 CUANTIFICACIÓN DE STOCK DE CARBONO EN UNA PLANTACIÓN DE PINO (Pinus radiata) EN ESTRATO DE PÁRAMO DE OZOGOCHE BAJO, PARQUE NACIONAL SANGAY. ............................................................................. 42 EFECTO DE LA FRAGMENTACIÓN DEL BOSQUE TROPICAL SECO SOBRE LA VIABILIDAD DE PROGENIES DEL ÁRBOL DIOICO Brosimum alicastrum (Moraceae) ........................................................................................................ 54 EVALUACIÓN ECOLÓGICA RÁPIDA DE LA FAUNA EXISTENTE EN EL BOSQUE TROPICAL SIEMPRE VERDE DEL CANTÓN EL CARMEN, MANABÍ ........................................................................................................................... 66 ESTRUCTURA Y COMPOSICIÓN DEL BOSQUE SECO SEMIDESIDUO DE LA RESERVA DE BOSQUE SECO LALO LOOR DEL CANTÓN JAMA. MANABÍ ................................................................................................................. 73 ESTRUCTURA Y COMPOSICIÓN FLORÍSTICA DEL BOSQUE TROPICAL SIEMPRE VERDE DEL CANTÓN EL CARMEN, MANABÍ ......................................................................................................................................................... 79 CONTRIBUCIÓN A LA DIVERSIDAD Y SIMILITUD ARBOREA DE LOS BOSQUES MONTANOS ALTOS DEL ECUADOR CENTRAL ..................................................................................................................................................... 84 DIVERSIDAD Y COMPOSICIÓN FLORÍSTICA EN LA ESTACIÓN CIENTÍFICA RÍO PALENQUE .............................. 93 CARACTERIZACIÓN DE LAS ÁREAS NATURALES (BOSQUES Y PÁRAMOS) COMO HERRAMIENTA DE ORDENAMIENTO TERRITORIAL EN LA PROVINCIA DE CHIMBORAZO. .................................................................. 99 CONSERVACIÓN Y USO SOSTENIBLE DE RECURSOS GENÉTICOS FORESTALES ....................................... 106 PATRONES ESPACIALES DE RECLUTAMIENTO Y VARIABILIDAD GENÉTICA DE LA PALMA Oenocarpus bataua Mart., EN EL PARQUE NACIONAL YASUNÍ (PNY) ..................................................................................................... 107 DIVERSIDAD GENÉTICA DEL MORETE (Mauritia flexuosa L. f.) EN PANTANOS DE LA REGIÓN SEPTENTRIONAL DEL PARQUE NACIONAL YASUNÍ ............................................................................................................................. 114 PRIORIDADES DE INVESTIGACIÓN GENÉTICA Y ECOLÓGICA PARA LA CONSERVACIÓN DE ESPECIES Y BOSQUES Y EL DESARROLLO FORESTAL EN ECUADOR ..................................................................................... 123 ESTRUCTURA Y DIVERSIDAD GENÉTICA DE LA PALMERA Oenocarpus bataua Mart. (ARECACEAE): UNA HERRAMIENTA PARA LA CONSERVACIÓN DE UN RECURSO OLEAGINOSO FORESTAL .................................. 131 EVALUACIÓN DEL ESTADO DE CONSERVACIÓN Y CARACTERIZACIÓN POBLACIONAL DE DOS ESPECIES FORESTALES PRIORIZADAS EN LA MICRO-CUENCA DEL RIO CHIMBORAZO. ................................................... 139 COLECTA Y CARACTERIZACIÓN MORFOLÓGICA IN SITU DE CAPULÍ (Prunus serotina Ehrh) DEL BANCO NACIONAL DE GERMOPLASMA DEL INIAP-ECUADOR ........................................................................................... 145 GESTIÓN FORESTAL SOSTENIBLE ....................................................................................................................... 152 COMPORTAMIENTO AGRONÓMICO DEL LAUREL (Cordia alliodora), BAJO DOS SISTEMAS DE PASTOREO EN LA ZONA DE FEBRES-CORDERO, PROVINCIA DE LOS RÍOS ................................................................................ 154 EFECTOS DE LA EXCLUSIÓN DE GRANDES HERBÍVOROS SOBRE LA VEGETACIÓN Y EL SUELO, EN MINAS DE CARBÓN RESTAURADAS EN EL NORTE DE PALENCIA ................................................................................... 162 ESTRUCTURA POBLACIONAL, PRODUCTIVIDAD Y FENOLOGÍA FLORAL DE LA TAGUA (Phytelephas aequatorialis) EN LA COSTA ECUATORIANA ............................................................................................................. 169 PROPUESTA METODOLÓGICA DE CRITERIOS E INDICADORES DEL MANEJO FORESTAL SOSTENIBLE EN LAS CONDICIONES DE CUBA .................................................................................................................................... 176 EFECTO DE VARIABLES CLIMÁTICAS Y LITOLÓGICAS SOBRE EL CRECIMIENTO EN ALTURA DOMINANTE EN MASAS DE Pinus nigra Arn .......................................................................................................................................... 183 ADS80 AND MODIS BASED AND REMOTE SENSING APPLICATIONS IN FOREST MANAGEMENT .................... 190 VIABILIDAD TÉCNICA, PRODUCTIVA, COMERCIAL Y AMBIENTAL DEL JABÓN ARTESANAL “Yakumama” ELABORADO CON PRODUCTOS FORESTALES NO MADERABLES DE LA COMUNIDAD KICHWA RÍO BLANCO ...................................................................................................................................................................................... 202 VALIDACIÓN DE UN PROTOCOLO PARA LA PROPAGACIÓN MASIVA IN VITRO DE Paulownia elongata, Paulownia fortunei y un híbrido (Paulownia fortunei x Paulownia elongata) BAJO SISTEMAS DE PROPAGACIÓN CONVENCIONAL E INMERSIÓN TEMPORAL ............................................................................................................ 208 ZONIFICACIÓN AGROECOLÓGICA DEL CULTIVO DE CAUCHO Hevea brasiliensis EN EL ECUADOR EN BASE A LA INCIDENCIA DEL MAL SURAMERICANO DE LAS HOJAS ................................................................................... 214 ANÁLISIS DE LOS PATRONES DE CRECIMIENTO RADIAL DE MASAS DE Pinus pinaster Ait., EN RESPUESTA A LA VARIABILIDAD CLLIMÁTICA EN EL CENTRO-SUR DE ESPAÑA ........................................................................ 219 POBLACIONES RURALES Y MEDIOS DE VIDA ASOCIADAS AL BOSQUE ........................................................ 227 SEGURIDAD ALIMENTARIA EN COMUNIDADES INDÍGENAS DE COSTA RICA: EL CASO DE COMUNIDADES CABÉCAR DE ALTO CHIRRIPÓ .................................................................................................................................. 228 BIOINDICADORES: “SEGUROS LOCALES” PARA PEQUEÑOS PRODUCTORES AGROPECUARIOS ................. 236 SISTEMAS INTEGRADOS: SISTEMAS AGROFORESTALES................................................................................ 240 EVALUACIÓN DE ESPECIES FORRAJERAS BAJO SISTEMA SILVOPASTORILES PARA LA SOSTENIBILIDAD DE LA GANADERÍA EN LA AMAZÓNICA ECUATORIANA ............................................................................................... 242 MATERIAL DIDACTICO PARA EL APRENDIZAJE AGROFORESTAL ...................................................................... 252 ESTABLECIMIENTO DE ESPECIES ARBÓREAS DE USO AGROFORESTAL MEDIANTE TÉCNICAS CONSERVACIONISTAS EN CONDICIONES DE SECANO ........................................................................................ 258 ENLAZANDO CONOCIMIENTO LOCAL Y RASGOS FUNCIONALES DEL COMPONENTE ARBÓREO, BANANOS Y PALMAS EN SISTEMAS AGROFORESTALES CON CACAO Y CAFÉ EN COSTA RICA.......................................... 263 RUNA GUAYUSA – DESARROLLO DE UN SISTEMA DE CULTIVO AGROFORESTAL DE Ilex guayusa Loes. ...... 271 EVALUACIÓN DE ALTERNATIVAS SILVOPASTORILES QUE PROMUEVAN LA INTENSIFICACIÓN Y RECUPERACIÓN DE PASTURAS DEGRADADAS Y CONTRIBUYAN A REDUCIR EL IMPACTO AMBIENTAL DE LA ACTIVIDAD GANADERA EN LA AMAZONÍA ECUATORIANA AL SEGUNDO AÑO DE ESTABLECIMIENTO. ......... 280 Ilex guayusa COMO MOTOR PARA EL DESARROLLO SOSTENIBLE EN LOS SISTEMAS AGROFORESTALES EN LA AMAZONÍA OCCIDENTAL ...................................................................................................................................... 287 MEJORAMIENTO DE CHAKRAS, UNA ALTERNATIVA DE SISTEMA INTEGRADO PARA LA GESTIÓN SOSTENIBLE DE BOSQUES EN COMUNIDADES NATIVAS DE LA AMAZONÍA ECUATORIANA........................... 296 RESÚMENES DE PONENCIAS ................................................................................................................................ 305 EVALUACIÓN DEL ESTADO DE CONSERVACIÓN DE DOS ESPECIES FORESTALES PRIORIZADAS EN LA SUBCUENCA DEL RÍO QUIJOS, NAPO, ECUADOR ......................................................................................................... 307 ESTADO DEL BOSQUE PROTECTOR CHONGÓN Y COLONCHE SUBCUENCA LOMA ALTA Y AMPLIACIÓN, CORDILLERA CHONGÓN COLONCHE Y LA SUBCUENCA DEL RÍO CHONGÓN EN EL CANTÓN SANTA ELENA ...................................................................................................................................................................................... 308 “INCORPORACIÓN DE ÁRBOLES MADERABLES DE ALTO VALOR EN PLANTACIONES NUEVAS DE CACAO FINO EN EL NORTE DE LA AMAZONÍA ECUATORIANA”.......................................................................................... 309 CRECIMIENTO Y FENOLOGÍA DE 13 ESPECIES ARBÓREAS DEL BOSQUE SECO TUMBESINO. RESERVA NATURAL LAIPUNA – MACARÁ – LOJA - ECUADOR ................................................................................................ 310 PROSPECCIÓN DE SEMILLAS DE ESPECIES LEÑOSAS NATIVAS DE USO MÚLTIPLE (AUM) PRIORIZADAS POR LAS COMUNIDADES KICHWAS CON POTENCIAL AGROFORESTAL PARA EL MANEJO SOSTENIBLE DE LA TIERRA ......................................................................................................................................................................... 311 ORQUÍDEAS: PRODUCTO FORESTAL NO MADERABLE (PFNM) ........................................................................... 312 USO DE LA DIVERSIDAD GENÉTICA PARA LA CONSERVACIÓN DEL BOSQUE CASO DEL CULTIVO DE LA NARANJILLA EN ECUADOR ........................................................................................................................................ 313 EL GUARANGO (Caesalpinea spinosa Kuntze), UNA ESPECIE FORESTAL DE PROPÓSITO MÚLTIPLE PARA HACER FORESTACIÓN PRODUCTIVA ...................................................................................................................... 314 CONDICIONES DE SITIO Y CRECIMIENTO DE Centrolobium ochroxylum ESPECIE NATIVA CON POTENCIAL AGROSILVOPASTORIL DEL LITORAL ECUATORIANO ............................................................................................ 315 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 PRESENTACIÓN El Instituto Nacional de Investigaciones Agropecuarias (INIAP), en colaboración con SENESCYT, MAGAP, ESPOCH y la GIZ, desarrolló el Primer Encuentro Nacional Bosques, Recursos Genéticos Forestales y Agroforestería. Este Encuentro pretende celebrarse cada dos años y convertirse en un referente para la difusión de los trabajos científicos de los investigadores nacionales y afianzar la fructífera colaboración que se viene desarrollando entre los miembros de los diferentes grupos de trabajo, entre los que se incluyen investigadores y profesionales que comparten problemas y objetivos comunes en el ámbito de la investigación de sistemas forestales. La variedad geográfica de nuestros ponentes junto con los ecosistemas estudiados y las aportaciones de investigadores invitados, hace que las presentaciones durante el encuentro hayan sido diversas, constituyéndose en un lugar de encuentro privilegiado para el intercambio de ideas, tecnología, metodologías y oportunidades en el ámbito de la conservación de sistemas forestales. Con ello se pretende facilitar la transferencia o intercambio de información, tecnología y/o metodologías entre investigadores y gestores forestales de las administraciones públicas y de empresas, y generar un sitio para la coordinación de trabajos en relación con el cultivo en vivero e implantación de especies en sistemas forestales. Me gustaría agradecer de forma especial la participación en la jornada de los Drs. Bryan Finegan de CATIE-Costa Rica, Carlos Balbino de EMBRAPA/Procitropicos-Brasil, Jonathan Cornellius del ICRAF-Perú, Felipe Bravo y Mirén del Río de la Universidad de Valladolid-España, Leo Gallo del INTA-Argentina, Eduardo Morillo de INIAP-Ecuador, Rodrigo Sierra de GEO-15 y Mario Añazco de la Universidad Técnica del Norte y de manera general a todos los expositores que participaron en el evento. Espero que este libro sirva para orientar y esclarecer a estudiantes, maestros e investigadores sobre la gestión de bosques, la evaluación del estado de conservación de los Recursos Genéticos Forestales y el desarrollo futuro de la agroforestería en el Ecuador. Dr. Jorge Grijalva Coordinador del evento 6 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 INTRODUCCIÓN Los días 13, 14 y 15 de noviembre de 2013 se celebró en el Centro de Convenciones Eugenio Espejo de la ciudad de Quito-Ecuador, EL PRIMER ENCUENTRO NACIONAL DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA. Se realizaron 52 presentaciones: 10 magistrales, 31 orales y 11 paneles, que fueron impartidas por destacados expertos, locales y foráneos, en la gestión y manejo forestal sostenible, la evaluación de recursos genéticos forestales en el país, Europa, América Latina y el Caribe, así como en el diseño y evaluación de sistemas agroforestales/Sistemas Integrados como propuestas sobresalientes de uso sostenible de la tierra, de cara al futuro el análisis de relaciones entre bosques y cambios climáticos. La variedad temática de los trabajos presentados fue notable, agrupándose en cinco sesiones temáticas: Gestión Forestal sostenible, Conservación y uso sostenible de recursos genéticos forestales, Poblaciones rurales y medios de vida asociadas al bosque, Bosques y cambios climáticos y Sistemas integrados: sistemas agroforestales. Este Encuentro fue planificado con el propósito de recoger, valorar e intercambiar experiencias y conocimientos relacionados con la gestión de bosques, la evaluación del estado de conservación de los recursos genéticos forestales y el desarrollo y futuro de la Agroforestería en el país. La parte protocolaria del evento estaba a cargo del Ing. Raúl Ramos del Programa Nacional de Forestería. Las Jornadas fueron inauguradas por el Ing. Pablo Noboa, Subsecretario Forestal del MAGAP y clausuradas por el Ing. Fernando Romero, Decano de la Facultad de Recursos Naturales de la ESPOCH. La coordinación del Encuentro lo realizó el equipo técnico del Programa Nacional de Forestería del INIAP. Es una satisfacción presentar esta publicación que reúne los resúmenes de todas las contribuciones científicas presentadas, así como las correspondientes a las ponencias magistrales. Una edición digital del libro de resúmenes es accesible desde la página web del INIAP http://www.iniap.gob.ec La coordinación del Encuentro quiere agradecer la dedicación así como la activa y enriquecedora participación de todos los expositores en sus comunicaciones y en la discusión posterior de los resultados presentados y sobre todo a los técnicos, becarios y demás personal administrativo del Programa Nacional de Forestería y del INIAP en general que colaboraron activamente en la organización. Comité Organizador 7 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 CONFERENCIAS MAGISTRALES Leonardo Gallo, INTA RECURSOS GENÉTICOS FORESTALES, ALTERNATIVAS DE USO Y CONSERVACIÓN EN EL MUNDO CONTEMPORÁNEO Felipe Bravo Oviedo, Universidad de Valladolid EL INVENTARIO FORESTAL NACIONAL DE ESPAÑA: HERRAMIENTA PARA LA INVESTIGACIÓN Y LA GESTIÓN FORESTAL Miren del Río, INIA PLANTACIONES FORESTALES MIXTAS. UNA OPORTUNIDAD PARA MEJORAR LA PRODUCTIVIDAD Y LA RESILIENCIA Mario Añazco, Universidad Técnica del Norte PRODUCTOS FORESTALES NO MADEREROS (PFNM) EN EL ECUADOR…UN APORTE AL MANEJO FORESTAL SOSTENIBLE Jonathan Cornelius, ICRAF DOMESTICACIÓN DE ESPECIES FORESTALES PARA AGROFORESTERÍA Rodrigo Sierra, GEO-15 PATRONES Y DETERMINANTES HISTÓRICAS Y GEOGRÁFICAS DE LA DEFORESTACIÓN EN EL ECUADOR, 1950-2010 Y TENDENCIAS AL FUTURO 8 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 RECURSOS GENÉTICOS FORESTALES, ALTERNATIVAS DE USO Y CONSERVACIÓN EN EL MUNDO CONTEMPORÁNEO Leonardo Gallo Grupo de Genética Ecológica y Mejoramiento Forestal, INTA Bariloche, Argentina [email protected] El contexto de la pérdida del bien común genético forestal (= recursos genéticos forestales) El bien común boscoso cubre en el planeta una superficie de aproximadamente 4000 millones de hectáreas (36 % de bosques primarios) representando el 30 % de la superficie terrestre. Entre las diferentes funciones de los bosques se consigna que la productiva representa solo el 34 % cubriendo las funciones sociales, de conservación, de protección o finalidades múltiples el resto (FAO 2007). Anualmente se pierden en total 13 millones de has, la mitad en los bosques primarios. Entre las causas técnicas y amenazas que más se destacan como responsables de la pérdida de superficie boscosa se encuentran: la substitución del bosque para uso agrícola y ganadero, la explotación forestal (= aprovechamiento descontrolado), los incendios forestales, la extracción selectiva sin manejo planificado, la substitución del bosque nativo por plantaciones de especies exóticas y el cambio climático. Sin embargo en los países latinoamericanos (bien se podría decir en los países periféricos de todos los continentes) a esas causas técnicas se suman causas socio-económicas que son muchas veces los verdaderos motivos de la pérdida de superficie boscosa. Entre ellas se destacan tres: la fuerte dependencia económica internacional (el uso del suelo muchas veces se decide fuera de nuestros países), la corrupción (existen buenas leyes y normas de aplicación pero fallan generalmente los controles) y la pobreza que aumenta la presión sobre el bosque (el principal uso de la madera continúa siendo desde el período neolítico para leña). Con respecto a la primera causa socio-económica mencionada arriba, la misma se ha repetido en la historia de nuestros países cientos de veces. El avance de la frontera agrícola y ganadera viene asociado a nuestro papel de proveedores de materias primas al “mundo desarrollado”. Muchos de los grandes cambios del uso del suelo con bosques tienen relación con la definición de políticas de desarrollo mundial por parte de los centros de poder económico y han sido originadas en el marco de la teoría de distribución internacional del trabajo. Por ejemplo en el sur de nuestro continente, la misión del Banco Mundial que recorrió en la década de los 60 la región, definió entre otras cosas que en Paraguay, Argentina, Uruguay y Brasil debían ser productores de madera de pinos y eucaliptos y proveedores de porotos de soja. El cambio de uso del suelo encontró cierta resistencia en los 70 por los movimientos políticos que reivindicaban una mayor soberanía nacional y un cambio en el modelo socio-económico de la región. La respuesta de los grandes centros de poder económico fue el establecimiento de dictaduras militares, genocidio de parte del pueblo que se oponía al modelo neoliberal, e imposición a sangre y fuego de su sistema de explotación. A partir de entonces aumentó ostensiblemente la tasa de deforestación en la región. El caso argentino es paradigmático. La curva de crecimiento exponencial de la superficie de soja se inicia en 1977 (Fig. 1), a un año de la toma del poder político y económico por parte de la cruenta dictadura militar que impuso el modelo neoliberal, originó 30.000 desaparecidos, quintuplicó la deuda externa, estuvo a punto de iniciar a una guerra con Chile y 9 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 condujo a otra contra Inglaterra. La imposición de ese monocultivo a partir de la deforestación, generó una fuerte dependencia económica de la que aún no puede salir el país. . Soja Trigo Girasol Maíz Figura 1. Aumento exponencial de la superficie del cultivo de soja, principal causa socio-económica de la pérdida de superficie boscosa en Argentina, impuesta por los centros de poder económico internacional a partir de la dictadura militar instalada en 1976. (Nótese que la curva asciende exponencialmente a partir de1977) La importancia de la relegada diversidad genética El concepto de biodiversidad fue acuñado por consenso político durante las reuniones del Convenio sobre la Diversidad Biológica organizado por la Naciones Unidas en Rio de Janeiro en Junio de 1992. Allí se definió que en términos generales la Biodiversidad está conformada por tres niveles de variación: diversidad de los ecosistemas, diversidad entre las especies y diversidad dentro de cada especie. Otros autores extendieron la idea con más detalle acotando que Biodiversidad comprende desde la diversidad dentro de la molécula de ADN, es decir la diversidad de las bases nucleotídicas, hasta la diversidad entre los ecosistemas (ej. Simitinand 1995) Sin embargo, a pesar del reconocimiento por definición del nivel que incluye a la diversidad genética, muy pocas veces ella es tenida en cuenta. Se la reconoce como muy importante y se le asigna un papel dentro de la complejidad de la diversidad biológica pero probablemente porque su evaluación, interpretación y monitoreo es tema de especialistas, no se incluye en casi ninguno de los estudios ecorregionales que pretenden ordenar y manejar en forma sostenible los ecosistemas boscosos en toda su integridad. No obstante ello la importancia de la diversidad genética en la sostenibilidad de la vida del planeta en general y en la sostenibilidad productiva, ecológica y social de los ecosistemas boscosos en particular ya se encuentra teóricamente consolidada y prácticamente probada con infinidad de ejemplos. De ella depende la capacidad de adaptación actual y futura de las poblaciones boscosas (Namkoong et al. 2000) y por lo tanto constituye la base de su potencialidad evolutiva, es decir de su productividad y sus beneficios ambientales y sociales. Qué conservar? 10 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Los organismos vivos poseen dos grandes estrategias para adaptarse a los cambios del ambiente biótico y abiótico: la plasticidad fenotípica y la adaptación evolutiva. En el primer caso no se modifica el genotipo del individuo. Un mismo genotipo puede modificar su fenotipo adaptándolo a los cambios ambientales. Por ejemplo, la parte superior de la copa de un árbol expuesta a la radiación directa presenta hojas más pequeñas y coriáceas que las hojas inferiores y del interior de la copa. Algunos cambios ambientales moderados pueden ser amortiguados por esta plasticidad fenotípica que se constituye así en un mecanismo adaptativo muy eficiente. Pero si los cambios ambientales son muy intensos y/o permanentes los umbrales de plasticidad fenotípica de los diferentes genotipos son superados y su aptitud reproductiva se ve afectada. En ese caso la población reacciona modificando las frecuencias génicas esencialmente en dos etapas: primero los cambios ambientales profundos afectan la sobrevivencia de la regeneración en el ambiente modificado. La etapa de germinación y establecimiento constituyen las más sensibles en la vida de las poblaciones arbóreas. Pero si el cambio es extremo, mueren individuos adultos y por lo tanto se modifica la frecuencia de los genes también en la población adulta. Quedan de esa manera solo los genotipos adultos aptos que se cruzarán entre sí y generarán una descendencia adaptada a las nuevas condiciones ambientales. De esta manera, la población, evoluciona. En biología, evolución significa fundamentalmente un cambio en la frecuencia de los genes de una población. La unidad evolutiva es por lo tanto la población. Es por ello que conservar ecosistemas y especies no garantiza la conservación de la diversidad genética. Nos debemos focalizar en estudiar, ordenar y monitorear la variación genética que existe entre las poblaciones. Una vez caracterizada la variación genética dentro de una especie, los programas de conservación de los recursos genéticos forestales deben orientarse a definir cómo conservar las poblaciones con mayor diversidad y/o con alguna particularidad genética. Mantener la mayor diversidad genética es la estrategia más segura para permitir a las poblaciones su adaptación a los cambios ambientales, particularmente en el contexto de Cambio Climático en el que estamos inmersos. Esa variación intra-específica constituye gran parte de lo que comúnmente denominamos “recursos genéticos”, cuya definición e identificación más precisa deberíamos buscarla en el uso ancestral que las comunidades de los pueblos originarios les han otorgado. Los métodos tradicionales de evaluación de los recursos genéticos forestales La genética forestal utiliza los denominados ensayos de ambiente común para discernir qué proporción de la varianza fenotípica de una característica de interés es heredable. Al criar la descendencia de diferentes poblaciones en un mismo ambiente, las diferencias de crecimiento, u otra característica, entre las poblaciones se pueden inferir como de origen genético. Este principio de homogeneizar el ambiente para poder detectar la varianza atribuible a la contribución genética dentro de la varianza fenotípica total, es el principio rector de todos los ensayos de procedencias, pruebas de progenie y ensayos clonales. El genetista y el mejorador tienen, por lo tanto, siempre en cuenta el ambiente en todas sus evaluaciones. Desde lo metodológico el ambiente ya está incorporado en la percepción de la realidad biológica del genetista como una de las fuentes de variación que, interactuando con la información genética, da lugar a la expresión fenotípica. Lamentablemente no ocurre lo contrario con la mayoría de los profesionales relacionados a las ciencias biológicas. Gestores de bosques, ecólogos, biólogos, ingenieros forestales y agrónomos, muy poca veces consideran que parte de la variación que observan en los sistemas que estudian o manejan, es debida a cuestiones genéticas. Los primeros ensayos de procedencias se remontan a estudios de variación en Pinus sylvestris del Siglo XVII (Langlet 1970) y algunos autores indican que ya en el Siglo XIII en Japón, había comparaciones de procedencias de Cryptomeria japónica plantadas en un mismo sitio (Savolainen 2004). Con los ensayos de procedencias se demostró 11 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 que el 90 % de las especies arbóreas muestran una notable adaptación local, diferenciándose genéticamente entre ellas (Alberto et al. 2013) y gran parte (80 %) se distribuyen a lo largo de clines ambientales con variación genética clinal mimética (Fig. 2). Esta variación natural existente tiene dos consecuencias relevantes: por un lado, como cada población podría ser genéticamente diferente de las otras, conservar solo unas pocas poblaciones dentro de áreas protegidas podría implicar que estemos perdiendo importante variación genética pre-adaptada a condiciones ambientales requeridas en la actualidad o en el futuro, ubicada en poblaciones fuera del sistema nacional de áreas protegidas. Esto ocurre en algunos casos como el Ciprés de la Cordillera (Austrocedrus chilensis) en los Andes del Sur (Chile y Argentina). Del lado argentino, las poblaciones occidentales de la especie (húmedas y mésicas) se encuentran protegidas en gran parte dentro de los más de 1 millón de has de Parques Nacionales de esa región. Los fragmentos xéricos orientales de esta especie, por el contrario, se encuentran en propiedades privadas y son ellos los que, en conjunto, mantienen la mayor diversidad genética de la especie (Pastorino y Gallo 2002, Pastorino et al. 2004, Arana et al. 2010, Aparicio 2013). Por otro lado, desde el punto de vista de la producción, estas evidencias de diferenciación genética poblacional nos hacen remarcar la importancia del origen de la semilla cuando debemos elegir el material de propagación para actividades de forestación, restauración o enriquecimiento. Las ciencias forestales demostraron, 100 años antes que las otras disciplinas biológicas, que el origen de la semilla es vital para la adaptación de las plantaciones. En nuestra región Latinoamericana sabemos muy poco sobre la variación intra-específica de nuestras especies forestales nativas. El colonialismo cultural ha impactado también en ese aspecto ya que en el hemisferio sur cultivamos esencialmente especies forestales introducidas mientras que en el hemisferio norte cultivan principalmente sus especies nativas. Es por ello que resulta imprescindible iniciar programas de domesticación y mejora de nuestras especies forestales nativas en forma integral entre diferentes disciplinas como genética, fisiología, silvicultura, edafología, biología reproductiva, etc., de modo de conocer qué plantar, dónde plantar y cómo procesar el material de propagación y para lograr un mejor diseño y procedimiento de manejo de las plantaciones. Debemos cultivar nuestras especies ya que ello es una forma de generar o aumentar su valorización por parte de nuestras sociedades y con ello su conservación. Los métodos moleculares de evaluación de los recursos genéticos forestales Hace solo 50 años la investigación genética forestal y sus aplicaciones tecnológicas como el mejoramiento, incorporaron la utilización de marcadores genéticos moleculares. Estas herramientas han posibilitado en muchos casos llegar a resultados contundentes en tiempos muy breves. Los principales usos de esta tecnología se concentran en la caracterización de la diversidad genética, tanto dentro como entre poblaciones, identificación de individuos, análisis de parentesco, estudios de procesos de hibridación, flujo génico, selección asistida en programas de mejora, etc. El espectro que abarca la utilización de los marcadores moleculares es por lo tanto amplísimo y de diversa índole. A través de ellos se pueden resolver cuestiones tan prácticas como determinar el grado de contaminación polínica en un huerto semillero o la identificación de un clon para su registro como material de propagación comercial hasta el monitoreo de variantes genéticas que permitan estudiar el sistema de apareamiento y dispersión en un bosque nativo disturbado. 12 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figura 2. Variación genética clinal en daño por heladas dentro de una transecta altitudinal de Pinus devoniana (P. michoacana). Los orígenes de mayor altitud presentaron los menores daños. (de Saenz Romero y Tapia-Olivares 2008) Con el correr de los años estas herramientas se han hecho de uso corriente en muchos laboratorios ya que sus costos se redujeron sensiblemente. Lo importante es definir muy bien cuál es la pregunta científica o el problema tecnológico a resolver y elegir el marcador adecuado para tal fin. Se debe tener en cuenta que la fabricación de los equipos y la principal provisión de los reactivos provienen de los países industrializados del hemisferio norte que intentan hacer sus negocios basados muchas veces en nuestra ignorancia. Es por ello que hace falta conocer muy bien qué tecnología molecular se debe usar, cómo usarla y dónde usarla para minimizar los gastos de recursos económicos y humanos escasos en nuestra región. En primer lugar debemos definir si el problema de nuestro interés requiere del uso de marcadores de ADN nuclear (en general más variable) o del ADN ubicado en organelas (mitocondrias, cloroplastos). Este último, por ejemplo, por su menor tasa mutacional y su multiplicación clonal, mantiene las mutaciones génicas que se produjeron en tiempos remotos por lo que se ajusta muy bien para estudios filogenéticos y filogeográficos. Grandes zonas de similitudes genéticas entre poblaciones pueden ser identificadas en un plazo muy breve y con una gran seguridad y contundencia. Tal es el caso de la definición de dos grandes zonas genéticas en la distribución natural de Nothofagus nervosa (raulí) y Nothofagus obliqua (Roble) en Argentina. Estas especies de los bosques templados del sur de nuestro continente, presentan claramente una diferenciación latitudinal cuyo límite divisorio es una cadena de volcanes que corren en dirección Oeste-Este (Fig. 3). El estudio que permitió observar esta diferenciación se realizó en apenas dos años. Para llegar a la misma conclusión un estudio convencional con ensayos de campo habría necesitado al menos 20-30 años de evaluación (Marchelli y Gallo 2006, Azpilicueta, Marchelli y Gallo 2009) Los marcadores nucleares, poseen tasas mutacionales más grandes, por lo que su variabilidad es mucho mayor. En ellos debemos distinguir aquellos que responden a lo que se denomina “variación neutra” de los que identifican “variación adaptativa”. En el primer caso los de uso más adecuado en los últimos años son los microsatélites (SSRs), o repeticiones de secuencias simples de ADN. Con ellos es posible realizar la identificación genética de 13 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 individuos, el monitoreo de procesos de dispersión de polen y semillas, el análisis de reducciones poblacionales drásticas como consecuencia de disturbios naturales o antropogénicos (“cuellos de botella”) y su probable impacto genético (deriva génica), evaluación y monitoreo de procesos de hibridación interespecífica, etc. En los últimos años, aprovechando la facilidad y reducción de costos en la secuenciación del ADN, se ha comenzado a utilizar SNPs (polimorfismo de nucleótidos únicos) para parte de ese tipo de estudios. La combinación de estudios de estrés fisiológico (hídrico, térmico, lumínico, salino, etc.) con el monitoreo de marcadores moleculares ha permitido avanzar en el conocimiento de la relación entre caracteres fenotipos y la constitución genética de los individuos. Estudios de asociación genética, la identificación de loci marcadores de características cuantitativas (QTLs), genes candidatos y otros, permiten utilizar estas herramientas moleculares en programas de selección asistida que se incluyen dentro de los programas convencionales de mejora. Del mismo modo los modelos estadísticos empleados para la evaluación de caracteres fenotípicos de variación continua y el ranqueo de los individuos a propagar, pueden ser ajustados combinando en ellos la información obtenida a través del empleo de marcadores moleculares. Por ejemplo, en el ajuste de la matriz de parentesco en los modelos lineales mixtos empleados en la evaluación de pruebas de progenie instaladas a campo. L. Ñorquinco CHILE Lags. de Epulafquen 36º 49´ 39º 20' 71º 04’ L. Quillén North L. Tromen Vn. Lanín L. Huechulafquen L. Currhue Referencias 40º 00' N othof agus nerv os a N othof agus obliqua L. Lolog Bosque m ix to N othof agus sp. S. Martín de los Andes L. Lacar poblaciones RAULÍ L. Hermoso L. Espejo L. Traful ARGENTINA 40º40' 30 km 71º 21´ Figura 3. Variación genética en la distribución natural de Nothofagus nervosa y N. obliqua en la cordillera andina de Patagonia Norte, Argentina. Para ambas especies existe una clara diferenciación entre los haplotipos al norte y al sur de una cadena de volcanes que estarían marcando la línea divisoria en la evolución de esas variantes genéticas (Marchelli y Gallo 2006; Azpilicueta, Marchelli y Gallo 2009) 14 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 La mejor estrategia es por lo tanto el uso combinado de la información proveniente de las mediciones de vivero, ensayos de campo, bosque nativo realizadas en las plantas y su ambiente con la información proveniente de los laboratorios de genética molecular. Las herramientas moleculares son solo eso, herramientas, algo que uno usa integrando a otras fuentes de información para poder realizar una interpretación más adecuada de todos los resultados, jamás deben llegar a ser un fin en sí mismas. La utilidad de las herramientas moleculares es por lo tanto muy grande en la conservación y en el uso de los recursos genéticos forestales y seguirá avanzando con gran celeridad. Debemos saber utilizarlas para ahorrar costos y tiempo y para discernir en su utilización lo importante de lo superfluo, teniendo en cuenta siempre que detrás del uso de toda tecnología existe un interés comercial que beneficia generalmente a las empresas de los países dominantes que son quienes la generan, difunden y comercializan. 15 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 EL INVENTARIO FORESTAL NACIONAL DE ESPAÑA: HERRAMIENTA PARA LA INVESTIGACIÓN Y LA GESTIÓN FORESTAL Felipe Bravo Oviedo Instituto Universitario de Investigación en Gestión Forestal Sostenible Universidad de Valladolid-INIA, Avda. de Madrid 44, 34004, Palencia [email protected] Resumen Los Inventarios Forestales Nacionales componen bases de datos de gran valor tanto para describir el estado forestal del país estudiado como para el desarrollo de instrumentos de gestión y para la investigación selvícola y ecológica. En la presente comunicación se presenta el diseño y la evolución del Inventario Forestal de España y los trabajos realizados para comprobar la adecuación del diseño de muestreo y especial del tipo de parcela, diseñar nuevos métodos de muestreo para la madera muerta y para la estimación de variables. Después se presentan diversos trabajos realizados con datos del Inventario Forestal Nacional como el cálculo de balances de biomasa y CO 2, el desarrollo de herramientas selvícolas básicas y los modelos de crecimiento y producción. Finalmente se presentan ejemplos de análisis selvícolas a partir de los datos y las herramientas generadas con el Inventario Forestal Nacional de España. Introducción Los Inventarios Forestales Nacionales (IFN) son un fuente de datos muy valiosa para mejorar tanto la gestión de los bosques y sistemas agroforestales como para desarrollar un cuerpo científico adecuado para la realidad forestal que se estudie. Una planificación y un desarrollo cuidadosos de los trabajos asociados a los IFN permitirán no solo extraer la información relevante para el sector sino también generar conocimiento nuevo y desarrollar tecnologías que servirán para una gestión forestal sostenible. Objetivo El objetivo principal de esta presentación es mostrar el diseño del Inventario Forestal Nacional y los desarrollos científicos y tecnológicos desarrollados en España durante el último decenio. Material y Métodos A partir de datos del Inventario Forestal Nacional (IFN) español se ha desarrollado una ingente labor de investigación y transferencia tecnológica de gran utilidad para el sector forestal. Para ello se han utilizado las parcelas del IFN que se pueden descargar de forma gratuita de la página web del IFN (http://www.magrama.gob.es/es/biodiversidad/temas/inventarios-nacionales/inventario-forestal-nacional/) y se han integrado resultados de diversos proyectos de investigación. 16 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Resultados Los principales resultados obtenidos hasta la fecha y que serán presentados oralmente en el Primer Encuentro Nacional de Bosques, Recursos Genéticos Forestales y Agroforestería-Ecuador son los siguientes: Manejo de datos del IFN 1. Evaluación del diseño de muestreo del Inventario Forestal Nacional (IFN) de España (Bravo et al, 2002) 2. Herramienta informática BASIFOR (www.basifor.es) para el manejo de datos del IFN (Bravo et al, 2002) 3. Ecuaciones de biomasa (Montero et al, 2005 y Ruiz-Peinado et al, 2011 y 2012) 4. Contenido en carbono en diferentes tejidos de pinos (Pinus sp) y robles (Quercus sp) en España (Herrero et al, 2011 y Castaño y Bravo, 2012) 5. Balance del carbono fijado en diferentes macizos forestales (Bravo et al, 2007 y Herrero y Bravo, 2012) 6. Relación entre estructura forestal y diversidad de plantas vasculares (Osorio et al, 2009) Estimación de nuevas variables y diseño de nuevos métodos de cubicación 7. Modelo de masa incorporada en pinares mediterráneos (Bravo et al, 2008) 8. Modelo de madera muerta en pinares mediterráneos (Herrero et al, 2010) y la comparación del contenido en madera muerta de diversos ecosistemas en el mundo (Herrero et al, 2013) 9. Ecuaciones de perfil (Rodríguez et al, 2000, 2003, 2008 y 2013, Rodríguez, 2013) y la aplicación CUBIFOR (http://www.cesefor.com/cubifor/) para la cubicación de datos de inventarios. Herramientas selvícolas básicas 10. Caracterización de estructura forestal (Reque y Bravo, 2008) 11. Diagramas de manejo de la densidad forestal (Valbuena et al, 2008) Modelos de crecimiento y producción 12. Modelos de producción de rodal completo (Bravo y Montero, 2003) 13. Modelo de clase diamétrica (Escalante et al, 2011) 14. IBERO-Modelo de árbol individual (Bravo et al, 2010) parametrizado para Pinus sylvestris (Lizarralde et al, 2010) y Pinus pinaster (Lizarralde et al, 2010) integrando diversos submodelos de mortalidad (BravoOviedo et al, 2006), curvas de calidad (Rojo y Montero, 1996, Bravo y Montero, 2001 y Bravo-Oviedo et al, 2004), ecuaciones de crecimiento (Lizarralde, 2008) y de perfil (Lizarralde, 2008) 15. Estudios sobre el impacto del clima en el crecimiento forestal y su potencial influencia en los modelos de crecimiento (Bogino y Bravo, 2008 y 2009 Bogino et al, 2009, Bravo-Oviedo et al, 2008, Olivar et al, 2012 y Lara et al, 2013) 16. Plataforma de simulación forestal SIMANFOR (www.simanfor.es) para la generación de escenarios selvícolas y para la evaluación de modelos de crecimiento forestal (Bravo et al, 2012) Análisis selvícolas 17. Estudios de ciclo de vida de productos forestales (Lizarralde et al, 2008) 18. Evaluación económica de alternativas selvícolas (Bravo et al, 2008) 19. Diseño de estrategias de manejo adaptativo (Bravo et al, 2004, 2007 y 2008) 17 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Se pretende, en resumen, presentar un amplio catálogo de posibilidades de desarrollo de investigación y de herramientas de gestión forestal que anime a utilizar de forma adecuada las bases de datos de este tipo que en muchas ocasiones no son utilizadas en todo su potencial. Bibliografía Bogino, S.M., Bravo, F. 2008. Growth response of Pinus pinaster Ait. to climatic variables in central Spanish forests. Annals of Forest Science 65:506 1-13 DOI 10.1051/forest:2008025 Bogino, S.M., Bravo, F. 2009. Climate and intra-annual density fluctuations in Pinus pinaster in Spanish woodlands. Canadian Journal of Forest Research 39(8):1557-1565 Bogino, S.M., Fernández-Nieto, M.J., Bravo, F. 2009 Drought index and radial growth of Pinus sylvestris L. in its southern and western distribution threshold Silva Fennica 43(4):609-623 Bravo, F., Bravo-Oviedo, A., Diaz-Balteiro, L. 2008. Carbon sequestration in Spanish Mediterranean forest under two management alternatives: A modeling approach European Journal of Forest Research 127 (3): 225-234. DOI 10.1007/s10342-007-0198-y Bravo, F., Díaz-Balteiro, L. 2004. Evaluation of new silvicultural alternatives for Scots pine stands in northern Spain Annals of Forest Science 61:163-169 Bravo, F., LeMay, V., Gadow, K. von, Jandl, R. (Eds) 2008. Managing Forest Ecosystems: The Challenge of Climate Change. Springer. ISBN: 978-1-4020-8342-6 342 pages Bravo, F., Montero, G. 2001. Site index estimation in Scots pine (Pinus sylvestris L.) stands in the High Ebro Basin (northern Spain) using soil attributes Forestry 74(4):395-406 Bravo, F., Montero, G. 2003. High-grading effects on Scots pine volume and basal area in pure stands in northern Spain Annals of Forest Science 60:11-18 Bravo, F., Pando, V., Ordóñez, C., Lizarralde, I. 2008. Modelling ingrowth in Mediterranean pine forests: A case study from Scots pine (Pinus sylvestris L.) and Mediterranean maritime pine (Pinus pinaster Ait.) stands in Spain Investigación Agraria. Sistemas y Recursos Forestales 17(3):250-260 Bravo, F., Rodríguez, F., Ordóñez, C. 2012 A web-based application to simulate alternatives for sustainable forest management: SIMANFOR Forest Systems 21(2):4-8 Bravo-Oviedo, A., Rorig, S., Bravo, F., Montero, G. Rio, M. 2011 Environmental variability and site index evaluation in Mediterranean maritime pine site index Forest Systems 20(1):50-64 (available at http://recyt.fecyt.es/index.php/IA/article/view/9106/7989) Bravo-Oviedo, A., Sterba, H., Río, M., Bravo, F. 2006. Competition-induced mortality for Mediterranean Pinus pinaster Ait. and P. sylvestris L. Forest Ecology and Management 222:88-98 Bravo-Oviedo, A., Tomé, M., Bravo, F., Montero, G., Río, M. 2008. Dominant height equations including site attributes in generalized algebraic difference approach Canadian Journal of Forest Research 38:2348-2358 DOI: 10.1139/X08-077 18 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Castaño, J., Bravo, F. 2012 Variation in carbon concentration and basic density along stems of sessile oak (Quercus petraea (Matt.) Liebl.) and Pyrenean oak (Quercus pyrenaica Willd.) in the Cantabrian Range (NW Spain) Annals of Forest Science 69(6):663-672 available online at http://dx.doi.org/10.1007/s13595-012-0183-6 Escalante, E., Pando, V., Ordóñez, C., Bravo, F. 2011 Multinomial logit estimation of a diameter growth matrix model of two Mediterranean pine species in Spain. Annals of Forest Science 68(4):715-726 http://dx.doi.org/10.1007/s13595-011-0088-9 Herrero, C., Krankina, O.N., Monleón, V., Bravo, F. 2013 Amount and distribution of CWD in pine ecosystems of Northwest Spain, NW Russia and NW US iForest (disponible en http://www.sisef.it/iforest/pdf/?id=ifor0644-007) Herrero, C., Pando, V., Bravo, F. 2010 Modelling coarse woody debris in Pinus spp. plantations. A case study in northern Spain. Annals of Forest Science 67:708 (8 pages) Herrero, C., Turrión, B., Pando, V., Bravo, F. 2011 Carbon in heartwood, sapwood and bark along stem profile in three Mediterranean Pinus species Annals of Forest Science 68(6):1067-1076 http://dx.doi.org/10.1007/s13595-0110122-y Herrero, C., Bravo, F. 2012 Can we get an operational indicator of forest carbon sequestration? A case study from two forest regions in Spain Ecological Indicators (in press) available online at http://dx.doi.org/10.1016/j.ecolind.2011.04.021 Lara, W., Bravo, F., Maguire, D.A. 2013 Modeling patterns between drought and tree biomass from dendrochronological data: a multi-level approach Agricultural and Forest Meteorology 178-179:140-151 (available online at http://dx.doi.org/10.1016/j.agrformet.2013.04.017) Lizarralde, I. 2008. Dinámica de rodales y competencia en las masas de pino silvestre (Pinus sylvestris) y pino negral (Pinus pinaster Ait.) de los sistemas central e ibérica meridional. Tesis doctoral. Universidad de Valladolid (inédito) 256 pp Lizarralde, I., Broto, M., Rodríguez, F., Bravo, F 2008 Taper Equations and Wood Products: Assessing the Carbon Flow of the Forest Through Its Products. Managing Forest Ecosystems: The Challenge of Climate Change Managing Forest Ecosystems Volume 17, 2008, pp 165-177 Montero G., Ruiz-Peinado R., Muñoz M., 2005. Producción de biomasa y fijación de CO2 por los bosques españoles. Monografías INIA, Serie Forestal 13. Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria, Ministerio de Educación y Ciencia, Madrid, España, 270 pp. Olivar, J., Bogino, S., Spiecker, H., Bravo, F. 2012 Climate impact on growth dynamic and intra-annual density fluctuations in Aleppo pine (Pinus halepensis) trees of different crown classes Dendrochronologia 30:35-47 (available online at http://dx.doi.org/10.1016/j.dendro.2011.06.001) Osorio, LF, Bravo, F., Zaldivar, P. Pando, V. Forest structure and biodiversity in Maritime pine (Pinus pinaster Ait.) stands in Central Spain. Investigación Agraria: Sistemas y Recursos Forestales 18(3):314-321 Reque, J., Bravo, F. 2008. Characterizating forest structure types using data sets of National Forest Inventories Investigación Agraria: Sistemas y Recursos Forestales 17(2):105-113 (freely available on line at www.inia.es/srf) Rodríguez, F. 2013 Ecuaciones de volumen comercial para las principales especies maderables de Castilla y León. Documento Interno, descargable en http://www.cesefor.com/cubifor/ 19 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Rodríguez, F., Broto, M. Cubica v1.2: Programa informático para cubicar y clasificar productos a partir de los datos procedentes de un inventario. Montes Revista de Ámbito Forestal. 72:21-29 Rodríguez, F., Lizarralde, I, Bravo, F. 2013 Additivity on Nonlinear Stem Taper Functions: A Case for Corsican Pine in Northern Spain Forest Science 59(4):464-471 Rodríguez, F., Rodríguez, J. 2000 MatDendro v1.0: Programa para el cálculo de existencias con clasificación de productos. Montes: Revista de ámbito forestal. 62:13-16 Rodríguez, F; Broto, M.; Lizarralde, I. 2008 cubiFOR: Herramienta para cubicar, clasificar productos, calcular biomasa y CO2 en masas forestales de Castilla y León. Montes Revista de Ámbito Forestal (95):33-39 Rojo, A., Montero, G. 1996. El pino silvestre en la Sierra de Guadarrama. Ruiz-Peinado R., Río M., Montero G., 2011. New models for estimating the carbon sink capacity of Spanish softwood species. Forest Systems 20: 176-188. Ruiz-Peinado R., Río M., Montero G., 2012. Biomass models to estimate carbon stocks for hardwood tree species. Forest Systems 21: 42-52. Valbuena, P., Peso, C., Bravo, F. 2008. Stand Management diagrams for two Mediterranean pines in the Eastern of Spain Investigación Agraria: Sistemas y Recursos Forestales 17(2):97-104 (freely available on line at www.inia.es/srf) 20 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 PLANTACIONES FORESTALES MIXTAS: UNA OPORTUNIDAD PARA MEJORAR LA PRODUCTIVIDAD Y LA RESILIENCIA Miren del Río Gaztelurrutia Departamento Selvicultura y Gestión de Sistemas Forestales CIFOR-INIA, Ctra. A Coruña, km 7,5, 28040 Madrid, España Instituto Universitario de Investigación en Gestión Forestal Sostenible UVa-INIA [email protected] Las plantaciones forestales juegan un papel importante en el sector forestal, tanto a escala local como global. Por una parte, los bosques primarios se encuentran sometidos a una elevada presión antrópica, habiéndose reducido notablemente su superficie. Y por otra, la demanda de productos forestales se encuentra en crecimiento, por lo que es necesario aumentar la producción de madera. Las plantaciones forestales permiten incrementar esta producción sin aumentar la presión sobre bosques naturales, además de ofrecer otros servicios ecosistémicos. La mayor parte de las plantaciones con vocación productiva están formadas por una sola especie, a pesar de que las plantaciones mixtas pueden ofrecer importantes ventajas frente a las monoespecíficas (Kelty, 2006). En esta presentación se revisan estas ventajas centrándose en dos aspectos claves, la productividad y la resiliencia. La relación positiva entre diversidad de especies y productividad ha sido ampliamente estudiada en la literatura científica, aunque el número de estudios con especies forestales es limitado (Vilà et al 2013). Sin embargo, cuando el número de especies es reducido los resultados difieren en función de la composición específica y de otros factores, como las condiciones ambientales, densidad del rodal y fase de desarrollo. Para comprender las diferentes respuestas en términos de producción de las masas mixtas frente a las monoespecíficas es necesario abordar el estudio de las relaciones inter-específicas, que pueden ser de competencia, complementariedad y facilitación (Kelty 1992). Cuando las especies que cohabitan en un determinado sitio presentan características estructurales y funcionales diferentes, pueden aprovechar los recursos disponibles de un modo más eficiente conllevando una mayor producción en biomasa. Esta complementariedad de nichos o reducción de la competencia entre especies puede tener lugar en la parte aérea y/o en la parte radical, en función de las especies que conforman la mezcla. Entre las características de las especies que pueden conllevar una complementariedad en la parte aérea se encuentran el temperamento o tolerancia a la sombra, la estructura de la copa, la fenología, patrón de crecimiento, etc. Del mismo modo, la estructura del sistema radical y su fenología determinan la presencia o no de complementariedad (Jose et al. 2006). Con el fin de determinar si entre dos especies existe reducción de la competencia por complementariedad no es suficiente analizar estas características de las especies en masas monoespecíficas, ya que algunas de estas características varían dependiendo de si el árbol crece en mezcla con otras especies o no. Por ejemplo, para Fagus sylvatica se ha encontrado que la forma de la copa y la relación entre superficie de copa y diámetro normal varía entre masas monoespecíficas y mixtas (Dieler & Pretzsch, 2013). Otra causa de un aumento de la producción en masas mixtas es la presencia de facilitación entre especies, es decir, una especie beneficia directamente a la otra. Las interacciones de facilitación pueden deberse a una mejora del ciclo de nutrientes, a la capacidad de fijación de nitrógeno de algunas especies o a una mejora de las micorrizas. Existen 21 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 bastantes experiencias en plantaciones mixtas que abordan el estudio de la producción de una especie de interés, con frecuencia eucalipto, cuando se mezcla con alguna especie fijadora de nitrógeno (Forrester et al. 2006). Las interacciones entre especies se ven modificadas por diversos factores, entre los que destacan las condiciones ambientales o características ecológicas del sitio, la densidad del rodal y la fase de desarrollo en la que se encuentre la plantación. Según la hipótesis del gradiente de estrés (Stress Gradient Hypothesis, SGH) las interacciones positivas o de facilitación tienen lugar mayormente en ambientes dominados por un factor de estrés, mientras que en los sitios más benignos dominan las interacciones negativas o de competencia (Bertness y Callaway 1994). Este modelo conceptual, ha sido debatido activamente en la última década y se ha ampliado para tener en cuenta la naturaleza de la especie, el tipo de estrés o variación temporal (Holmgren y Sheffer 2010), aunque los estudios con especies forestales son poco abundantes (Pretzsch et al. 2010). La densidad de la plantación es otro factor determinante en las relaciones inter-específicas. En principio cabe esperar que estas relaciones son más intensas cuando las densidad es elevada, ya que los recursos son más limitados y los efectos positivos de la complementariedad y facilitación pueden ser más evidentes que a bajas densidades. Aunque este patrón es el más frecuentemente encontrado (Amoroso & Turnblom, 2006), se han encontrado tendencias contrarias para algunas especies, atribuidas a la presencia de competencia y facilitación simultáneamente (Condés et al 2013). Con respecto a la edad o fase de desarrollo, es importante considerar que las relaciones inter-específicas pueden variar considerablemente en las distintas fases de desarrollo, fundamentalmente debido a los distintos patrones de crecimiento de las especies (Forrester et al 2004). Finalmente, se revisa brevemente el efecto positivo que pueden tener las plantaciones mixtas frente a las monoespecíficas en la estabilidad y resiliencia frente a agentes bióticos y abióticos. Los trabajos de revisión sobre el efecto de la mezcla de especies en los daños por plagas son poco concluyentes (Griess & Knoke, 2011) fundamentalmente por el reducido número de trabajos que abordan este tema. En principio son dos los posibles mecanismos que pueden reducir el riesgo de daños por plagas: menor concentración de hospedantes y mayor diversidad de hábitats disponibles para los enemigos naturales de la plaga (Kelty, 2006); sin embargo, no siempre se han detectado efectos positivos de las mezclas. Con respecto a la estabilidad de las plantaciones frente a vendavales, el riesgo de daños dependerá en gran medida de la vulnerabilidad de las especies que conforman la mezcla. En general la mezcla de especie reduce el riesgo de este tipo de daños (Griess & Knoke, 2011). Estos autores recopilan algunos estudios que han abordado este tema para plantaciones de Picea abies en Centroeuropa, encontrando efectos positivos en la estabilidad cuando se mezcla con especies de frondosas, generalmente menos susceptibles a daños por vendavales. Las masas formadas por mezcla de especies modifican el ambiente forestal con respecto a las masas puras, siendo otra forma de facilitación. Del mismo modo que la complementariedad y la facilitación pueden mejorar la productividad a largo plazo especialmente en los sitios pobres, también se puede suponer que este efecto sea más patente en años con estrés elevado. Pretzsch et al. (2012) encontraron evidencias de facilitación de roble sobre haya en años de sequía extrema, con menor efecto de la sequía y mejor capacidad de recuperación tras la sequía (resistencia y resiliencia). Río et al (2013) estudiaron la variación temporal entre competencia y facilitación en dos mezclas de especies templadas centroeuropeas y encontraron que en los años con tasas altas de crecimiento dominaba la competencia mientras que en los años malos la facilitación. Es decir, las interacciones entre especies varían temporalmente según las condiciones de crecimiento anuales, como lo hace espacialmente a lo largo de gradientes ecológicos. No obstante, todavía son escasos los trabajos que han estudiado la interacción entre 22 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 especies en relación con la influencia del clima en el crecimiento y que constaten la ventaja de las masas mixtas frente al cambio climático. En definitiva, las plantaciones mixtas presentan a priori una gran potencialidad para mejorar las producciones de estos sistemas forestales y mejorar su estabilidad y resiliencia, aspecto fundamental en el marco del cambio global. Sin embargo, las interacciones entre especies son complejas y dependen de numerosos factores, por lo que para identificar mezclas de interés es necesario establecer dispositivos experimentales que permitan detectar estas interacciones. Referencias Amoroso M.M.; Turnblom E.C.; 2006. Comparing productivity of pure and mixed Douglas-fir and western hemlock plantations in the Pacific Northwest. Can. J. For. Res. 36, 1484-1496. Bertness M.D.; Callaway R.; 1994. Positive interactions in communities. Trends in Ecology & Evolution, 9, 191-193. Condés S.; Río M.; Sterba H.; 2013. Mixing effect on volume growth of Fagus sylvatica and Pinus sylvestris is modulated by stand density. Forest Ecology and Management 292, 86–95. Dieler J.; Pretzsch H.; 2013. Morphological plasticity of European beech (Fagus sylvatica L.) in pure and mixedspecies stands. Forest Ecology and Management 295: 97-108. Griess V.C.; Knoke T.; 2011. Growth performance, windthrow, and insects: meta-analyses of parameters influencing performance of mixed-species stands in boreal and northern temperate biomes. Can. J. For. Res. 41, 1141-1159. Kelty M.J.; 1992. Comparative productivity of monocultures and mixed-species stands. In: Kelty M.J.; Larson B.C.; Oliver C.D. (Eds.), The Ecology and Silviculture of Mixed-Species Forests, Kluwer Academic Publishers, Dordrecht (The Netherlands), pp. 125-141. Forrester D.I.; Bauhus J.; Khannac K.; 2004. Growth dynamics in a mixed-species plantation of Eucalyptus globulus and Acacia mearnsii. Forest Ecology and Management 193, 81–95. Forrester D.I.; Bauhus J.; Cowie A.L.; Vanclay J.K.; 2006a. Mixed-species plantations of Eucalyptus with nitrogenfixing trees: A review. Forest Ecology and Management 233, 211-230. Holmgren M.; Scheffer M.; 2010 Strong facilitation in mild environments: the stress gradient hypothesis revisited. Journal of Ecology, 98, 1269-1275. Jose S.; William R.; Zamorac D.; 2006 Belowground ecological interactions in mixed-species forest plantations. Forest Ecology and Management 233, 231–239. Pretzsch, H.; Block, J.; Dieler, J.; Dong, P.H.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zingg, A.; 2010. Comparison between the productivity of pure and mixed stands of Norway spruce and European beech along an ecological gradient. Ann. For. Sci. 67, 1-12. Pretzsch H.; Schütze G.; Uhl E.; 2012. Resistance of European tree species to drought stress in mixed versus pure forests: evidence of stress release by inter-specific facilitation. Plant Biology, 15, 483–495. 23 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Río M.; Schütze G.; Pretzsch H.; 2013. Temporal variation of competition and facilitation in mixed species forests in Central Europe. Plant Biology. doi:10.1111/plb.12029. Vilà M.; Carrillo-Gavilán A.; Vayreda J.; Bugmann H.; Fridman J.; Grodzki W.; Haase J.; Kunstler G.; Schelhaas M.; Trasobares A.; 2013 Disentangling Biodiversity and Climatic Determinants of Wood Production. Plos One 8(2):e53530. 24 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 PRODUCTOS FORESTALES NO MADEREROS (PFNM)….UN APORTE AL MANEJO FORESTAL SOSTENIBLE Mario José Añazco Romero Universidad Técnica del Norte [email protected] Resumen La sostenibilidad tiene raíces históricas en el campo forestal, en gran parte, la silvicultura practicada por pueblos y nacionalidades indígenas de la Amazonía ecuatoriana y de otros países se preocupaba de la preservación y conservación de los bosques como reservas de flora y fauna que son la base de su seguridad alimentaria y medicina natural. De otro lado las actividades forestales se caracterizan por tener una visión de largo plazo, porque varias especies forestales requieren turnos que superan los 50 años para poder cosecharse. Esto explica quizá por qué el sector forestal abrió el camino a la elaboración del moderno concepto de sostenibilidad. No obstante de lo anterior, el Manejo Forestal Sostenible (MFS) ha sido sesgado hacia el rendimiento sostenido de madera, ignorando los Productos Forestales No Madereros (PFNM) y servicios ecosistémicos que los bosques naturales y otros ecosistemas forestales brindan. También los enfoques metodológicos utilizados para el análisis de los PFNM han sido por lo general desde el diagnóstico (el problema), lo cual también ha contribuido para que estos no sean lo suficientemente visibilizados desde el potencial que ofrecen. Existen varias fuerzas o factores que influyen para lograr la propia sostenibilidad de los Productos Forestales No Madereros y para que estos aporten al MFS, entre ellos están factores restrictivos e impulsores de orden político, institucional, legal, económico, social, ambiental, cultural y técnicos. Entre los factores restrictivos resaltan la limitada información que existe sobre los PFNM, las escasas políticas y normas que orienten el manejo y uso, y las demandas del mercado que superan la oferta. Los factores impulsores más sobresalientes son la diversidad de usos históricos y sus aportes a la economía, la cultura y la seguridad alimentaria. La tendencia actual en el país es aproximarse hacia un Manejo Forestal Sostenible encaminado al uso múltiple del bosque, asegurando que no se disminuya su estructura y funcionalidad y por ende la capacidad de provisión de bienes y servicios. En otras palabras un ecosistema forestal ordenado de esta manera proveerá de madera sobre una base sostenible y continuará ofreciendo otros productos madereros como leña y carbón; al mismo tiempo podrá aportar con PFNM como fibras, tintes, resinas, gomas, taninos, alimentos, forrajes, entre otros; y mantendrá su función en cuanto a la preservación de la biodiversidad, protección del suelo, aportando en la regulación hidrológica y al cambio climático. Palabras claves: sostenibilidad, Productos No Madereros El Problema (diagnóstico) y el Potencial (pronóstico) Por lo general, la manera de acercarse a comprender la situación de los Productos Forestales No Madereros (PFNM) es desde el problema únicamente, lo cual genera una aproximación importante, no obstante también se aporta analizando paralelamente el potencial que estos productos tienen. En el presente caso se recurre al análisis 25 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 de algunos factores o fuerzas restrictivas (equivalentes al diagnóstico) y otras impulsoras (pronóstico) referente al aporte de los PFNM al Manejo Forestal Sostenible de los bosques naturales y otros ecosistemas forestales. Objetivo: La sostenibilidad de la sostenibilidad Los primero es lograr la sostenibilidad de los PFNM para que este resultado se convierta en un insumo hacia la sostenibilidad del bosque u otro ecosistema forestal, porque la insostenibilidad de los PFNM no podría ser un aporte al manejo forestal sostenible (MFS). Factores que restringen la contribución de los PFNM al Manejo Forestal Sostenible Los PFNM pueden ser vulnerables dependiendo de varios factores como las tecnologías utilizadas para su cosecha, las partes de la planta que se utilizan, los agentes de polinización, entre otros de orden político, económico, social, cultural y ambiental que a continuación se describen: 1.- Limitada información técnico-científica La información que permita tomar decisiones desde la óptica silvicultural, ecológica y económica, es limitada en el país, los PFNM de origen silvestre son cosechados bajo criterios que ofrece la fenología (madurez de frutos y semillas), las demandas del mercado y las propias necesidades de los usuarios de los bosques. Incluso especies que se consideran domesticadas como por ejemplo Juglans neotropica cuya corteza se utiliza como tinte natural, sus hojas son de uso medicinal y la nuez que es muy apreciada en mercados locales, tiene dificultades en su plantación y manejo por la presencia de agentes biológicos como la presencia de un lepidóptero Hypsyphylla grandella que la ataca en su fase primaria de crecimiento, para lo cual no se tiene suficientes investigaciones que permitan manejar esta situación. 2.- Reducido número de individuos/especie/superficie La composición (riqueza y diversidad) y estructura de los bosques, así como la frecuencia y abundancia de varias especies que proveen fibras, látex, resinas, aceites y frutos, constituyen un factor que restringe el manejo de los PFNM, en ocasiones se encuentra un solo individuo de una especie en una hectárea. Por ejemplo especies que proveen fibras naturales como la piquigua Evodiantus funifer y la yalica Heteropsis ecuadorensis, o aquellas que producen aceites como unguragua Oenocarpus bataua, tienen riesgos si se intensifican el uso. 3.-Métodos insostenibles de recolección que incluso afectan la regeneración natural El aprovechamiento de algunos PFNM ha provocado el exterminio de la especie que lo originaba y en otros casos provocó la diminución de la misma (Hiraoka 2001). En su tiempo en la Costa el palmiche Euterpe chaunostachyis que fue cosechado para ser comercializado a industrias que a su vez lo enlataban, provocó la disminución drástica de las poblaciones. 4.-Visiones sobre bosques y ecosistemas forestales Las visiones que han prevalecidos sobre los bosques y otros ecosistemas forestales, han sido “simplistas o reduccionistas” porque solo consideraban a estos como proveedores de madera (FAO 1994), lo cual influyó en las 26 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 políticas aún vigentes como es el caso de la Ley Forestal y de Conservación de Áreas Naturales y Vida Silvestre promulgada en el año 1981 con un enfoque de aprovechamiento de madera. También esta visión condujo a una subvaloración de los PFNM que dificulta que estos reciban apoyo político que se merecen. Actualmente se está proponiendo una visión integral e integradora donde los bosques a más de madera ofrecen otros productos como los PFNM y una variada gama de servicios ecosistémicos. 5.- Migración La migración es un factor que coarta las formas tradicionales de trasmitir información-conocimiento de generación en generación, lo cual ha sido importante para el uso y manejo de algunos PFNM. Por ejemplo, en algunos lugares la cosecha de frutos del morete Mauritia flexuosa se la realizaba derribando las palmas femeninas, mientras que las masculinas permanecían en pie, esto daba lugar a una disminución de la población, lo cual deja áreas extensas sin el material genético reproductivo debido a que la proporción de individuos masculinos sobrepasa a los femeninos, en poblaciones naturales, es de 5:1. Algunos grupos étnicos amazónicos para evitar esta situación aprendieron a reconocer los frutos femeninos y evitaron extraerlos y así mantener la reproducción de la especie, lo cual sin duda se perderá el momento que las nuevas generaciones no participen de esta forma de acceder a la información-conocimiento. 6.- Escasa normativa sobre PFNM En Ecuador el Ministerio del Ambiente inició el diseño de una norma para regular el uso y manejo de PFNM el año 2006, pero aún no ha sido promulgada, en estos días se encuentra actualizando la misma para someterla a consideración de la autoridad a fin que pueda ser finalmente aprobada y entre en vigencia lo más pronto posible. 7.- Otros factores Existen otros factores importantes de ser señalados como: i) los usos sobredimensionados de algunos PFNM; ii) la comercialización solo como materias primas sin valor agregado; iii) la degradación y deforestación; iv) la presión sobre los recursos forestales (ampliación de la frontera agrícola, mercado de especies de alto valor comercial madereras y no madereras, escaso porcentaje de tierras legalizadas que contienen bosques u otros ecosistemas donde se encuentran los PFNM, incremento de la población lo cual implica mayor demanda de alimentos y recursos naturales en general); v) un obstáculo adicional es la percepción general sobre los PFNM como especies que juegan un rol ecológico distinto al de especies sin importancia económica. Factores que impulsan la contribución de los PFNM al Manejo Forestal Sostenible 1.- Uso histórico La etnobotánica y etnomedicina dan cuenta del uso de varias especies entre las que sobresalen la Cinchona spp de la cual se utilizó la corteza rica en el alcaloide “quinina” muy reconocido en el tratamiento de los síntomas febriles ocasionados por la malaria o paludismo. Antes de las conquistas de los incas y de los españoles, los Paltas que habitaron gran parte del territorio que hoy comprende la provincia de Loja ya conocían las propiedades de esta planta conocida con los nombres comunes de cascarilla, habita en los bosques andinos. Se menciona que en 1378 el paludismo diezmó los ejércitos del inca Pachacutec, posteriormente en 1635 el producto fue introducido en 27 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Europa para uso médico. Otro PFNM que contribuyó al desarrollo de la industria automotriz fue el látex del caucho Castilla elastica que se explotó principalmente de los bosques amazónicos 2.- Forman parte de la cultura A lo largo del tiempo varios PFNM han formado parte de la cultura en diferentes lugares del país, por ejemplo el yaje o ayahuasca Banisteriopsis caapi se utiliza en rituales espirituales en la Amazonía ecuatoriana. El Sombrero de paja toquilla fue declarado como Patrimonio Cultural Intangible por la UNESCO el año 2012, este sombrero se confecciona con la fibra de la palma conocida como paja toquilla Carludovica palmata. Los Municipios de Cuenca y Quito declararon a varias especies arbóreas como patrimonio, entre ellas constan Erythrina edulis y Acacia macracantha. 3.- Representan el uso de la biodiversidad La diversidad de ecosistemas, especies vegetales y animales y los recursos genéticos constituyen la fuente principal de PFNM, así la biodiversidad que se encuentra en los bosques naturales es la que da origen de los PFNM considerados silvestres y la agrobiodiversidad constituye la fuente de PFNM domesticados. El Ecuador es reconocido por su biodiversidad, el mapa actual de ecosistemas elaborado por el Ministerio del Ambiente, registra 91 ecosistemas, de los cuales 65 son boscosos donde se encuentran la gran mayoría de PFNM. En cuanto a las especies, el Ecuador alberga alrededor del 10% de especies de flora y fauna que existen a nivel mundial de ahí el gran potencial de PFNM, por ejemplo Guazuma ulmifolia Lam. que se encuentra en los bosques secos tropicales tanto natural o intervenido, su corteza es utilizada como medicina natural para la diabetes, se utilizan árboles grandes que deben tener un mínimo de 50 cm de DAP y sólo utilizar corteza del tronco (Aguirre 2012). 4.-Diversidad de usos Los PFNM de origen vegetal y animal se caracterizan por una infinidad de usos. Añazco (2004) en un estudio realizado en el Ecuador continental con 600 especies encontró que estas generan alrededor de 3.000 PFNM. Existen especies que son más versátiles que otras, por ejemplo las palmas o los bambúes, estos últimos tienen registros de más de 1500 usos documentados, incluyendo, alimentos, bebidas, papel, adornos, cestas, lámparas, instrumentos musicales, abanicos, utensilios, juguetes, sistemas de irrigación y apoyos agrícolas, todos hechos con bambúes de diferentes especies. El principal bambú nativo de Ecuador es la Guadua angustifolia que es considerada como uno de los bambúes de América más apropiados para la fabricación de papel. 5.- Los PFNM forman parte de la economía del país PFNM como el cacao Theobroma cacao que es de origen amazónico ha contribuido históricamente a la economía del país empezando a finales del siglo XIX donde la economía del Ecuador había sufrido un cambio significativo. La expansión de la Costa, alentada por el incremento del cultivo del cacao para exportación, había desembocado en un boom que se extendió por varias décadas, desde 1880 hasta inicios del siglo XX. De 1880 a 1890, el Ecuador fue el mayor exportador mundial de cacao, y su comercio internacional adquirió proporciones inéditas. Con estos recursos se crearon los principales bancos del país y fue el principal motor para dinamizar la economía y el comercio nacional. Con ello se aceleró el crecimiento económico y se precipitaron cambios políticos y sociales de gran importancia. 28 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 6.- Los PFNM aportan a la soberanía y seguridad alimentaria Partiendo de la definición simplificada que la seguridad alimentaria es el acceso físico y económico a los alimentos y que es importante conocer su origen, los PFNM constituyen aportes significativos, por ejemplo especies domesticadas como el árbol de pan Artocarpus altilis que se maneja en sistemas agroforestales de la Costa y Amazonía tiene un valor nutricional que parte de un contenido proteico del 11%, es decir mayor que la papa, la yuca, el maíz, el plátano y el arroz; es rico en niacina, calcio, potasio y hierro. En la sierra las semillas cocidas de guato Erythrina edulis contienen 22% de proteína (superior al frijol y la lenteja), y un 51% en carbohidratos. El valor biológico de la proteína es mayor que la arveja y haba, es rico en potasio. 7.- Otros factores Existen otros factores que impulsan el uso y manejo de los PFNM: contribución a una industria ecológica como el caso de los taninos de guarango Caesalpinia spinosa y Polylepis spp utilizados en la industria del cuero. Otro factor es la inclusión en la malla curricular de las escuelas de Ingeniería Forestal de algunas Universidades del país, la asignatura PFNM. Bibliografía Aguirre, Z. 2012. Especies forestales de los bosques secos del Ecuador. Quito, EC. MAE/FAO-Finlandia. 140 p. Añazco, M; Loján, L; Yaguache, R. 2004. Productos forestales no madereros en el Ecuador (PFNM)….una aproximación a su diversidad y usos. Quito, EC. MAE/FAO, Soboc Grafic. 190 p. FAO. 1994. El desafío de la ordenación forestal sostenible. Perspectivas de la silvicultura mundial. Roma, IT. 122 p. Hiraoka, M., Mora, S. comps. 2001. Desarrollo sostenible en la Amazonía ¿mito o realidad?. Quito. EC.Abya-Yala. 201 p. 29 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 DOMESTICACIÓN DE ÁRBOLES AGROFORESTALES: UN REPASO ANALÍTICO Jonathan Cornelius Centro Internacional de Investigación Agroforestal (ICRAF) [email protected] Introducción La domesticación de árboles agroforestales ha sido parte de la agenda de investigación agroforestal internacional durante más de dos décadas (Leakey et ál. 2012), aunque su avance varía de región en región. Aquí se revisan los conceptos de domesticación agroforestal con el fin de promover la maximización de su impacto, sobre todo haciendo hincapié en la necesidad de evitar conceptos restrictivos de ámbito y subestimación de retos potenciales. Posteriormente, se exponen varias prioridades de investigación. Definiciones y estudios de caso Existen varias definiciones de ‘domesticación’ y ‘domesticación agroforestal’. Por ejemplo, Simons y Leakey (2004) ofrecen la siguiente: “la domesticación de árboles agroforestales implica incorporar especies en la ampliación de cultivos... es impulsada por agricultores y pretende hacer coincidir la diversidad intraespecífica de muchas especies de árboles importantes a nivel local con las necesidades de subsistencia de los agricultores, los mercados para una amplia gama de productos y la diversidad de ambientes agrícolas”. Sin embargo, las características fundamentales de esta actividad se aprecian mejor por medio de ejemplos (abajo). Caso 1: Canarium indicum en Nueva Bretaña del Este, Papúa Nueva Guinea Históricamente, la provincia isleña de Nueva Bretaña del Este en Papúa Nueva Guinea ha dependido de la producción de cacao y copra. Ambas industrias han disminuido en los últimos años. En el caso de la copra, debido a la caída de los precios y, en el caso del cacao, debido a la irrupción del barrenedor de la mazorca del cacao (Conopomorpha cramerella: Gracillariidae) en el año 2006. Este último problema afectó de forma drástica la extensión de producción de cacao y las prácticas de manejo. Actualmente, el principal comprador de cacao promueve el concepto de cosecha de “cada árbol, cada mazorca, cada semana”. No todos los pequeños agricultores pueden sostener esta intensidad de manejo y, por lo tanto, el área con cacao se ha reducido. Este es el contexto inmediato de un esfuerzo de varias instituciones en la domesticación de Canarium indicum (Burseraceae), que produce una nuez bastante consumida (‘galip’) (Cornelius et ál. 2012; Wallace et ál. 2012; NARI 2013). El “galip” fue identificada como un posible sustituto del cacao, debido a su alta y parcialmente insatisfecha demanda interna, su potencial de exportación (comercializada por un productor en Vanuatu como “la mejor nuez suave del mundo”) y su manejo relativamente poco exigente. El programa incluye al Instituto Nacional de Investigación Agrícola (PNG National Agricultural Research Institute —NARI), en colaboración con dos instituciones australianas (James Cook University Agroforestry and Novel Crops Unit y University of the Sunshine Coast). El programa incluye caracterización de la variación intra e interespecífica en rasgos comerciales; selección participativa (con los agricultores) de fenotipos superiores y su conservación en bancos de germoplasma en el campo; desarrollo de métodos de propagación vegetativa; estudios de mercado domésticos y de exportación; métodos de procesamiento y embalaje; y desarrollo de vínculos con el sector privado. Para el año 2013, más de 60.000 plantas han sido distribuidas entre pequeños agricultores (Cornelius et ál. 2012, NARI 2013). 30 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Caso 2: Allanblackia en Ghana (Pye-Smith 2014) La Allanblackia (Clusiaceae) es un género de nueve especies que crece en África Occidental y Oriental. Aunque se ha usado tradicionalmente, las cualidades únicas del aceite de Allanblackia recién fueron reconocidas: se compone casi en su totalidad de ácidos oleicos y esteáricos y se funde a unos 34°C; es decir, permanece sólido a temperatura ambiente pero se derrite en la boca, una propiedad ideal para la producción de margarina. Como resultado, la empresa multinacional Unilever decidió cambiar una porción significativa de su producción de grasa blanca por aceite de Allanblackia. Sin embargo, las fuentes tradicionales (silvestres) son insuficientes para satisfacer la demanda. Por ello, se lanzó el Proyecto Novella, una alianza público-privada que incluye a Unilever, la Unión Internacional para la Conservación de la Naturaleza (UICN), la Organización de Desarrollo de los Países Bajos (SNV), World Agroforestry Centre (ICRAF) y socios locales en varios países, con el objetivo de establecer la Allanblackia como un cultivo para pequeños agricultores. El trabajo en investigación y desarrollo ha incluido investigación en métodos de germinación (hasta ahora un importante cuello de botella); propagación vegetativa; selección y establecimiento de bancos de germoplasma de muestras superiores; y cumplimiento de inspecciones de seguridad alimentaria europeas. Los socios de Novella planean alcanzar niveles de producción de 10.000 toneladas de aceite para el año 2020 y 60.000 toneladas para el año 2030, lo que involucraría a unos 40.000 agricultores en la siembra de 8 millones de árboles de alto rendimiento. Caso 3: Guazuma crinita en Perú La Guazuma crinita (bolaina blanca) es un árbol maderable de rápido crecimiento, nativo de la Amazonía peruana, y su madera es muy comercializada en esta zona. A mediados de los noventa, el ICRAF y sus socios en Perú llevaron a cabo un proceso de priorización, que incluyó consultas con agricultores y técnicos (ingenieros forestales, especialistas en mercadeo e investigadores) para identificar especies para la domesticación de árboles; la bolaina fue una de las especies seleccionadas. Posteriormente, se realizaron ensayos de procedencia, que condujeron a la priorización de poblaciones locales (Cuenca de Aguaytía) y a selecciones dentro de poblaciones. Se llevaron a cabo varios ensayos de descendencia en finca, diseñados para su posterior conversión en huertos semilleros de plántulas. Estos huertos semilleros ahora suministran gran parte de la semilla para las actividades de reforestación con bolaina en Perú. El Instituto de Investigaciones de la Amazonía Peruana, con el apoyo de ICRAF, está desarrollando un programa de selección clonal con base en la diversidad capturada en los huertos semilleros. El trabajo en genética ha sido complementado con investigaciones sobre efectos en el sitio, captura de carbono y transformación. Existen muchos otros ejemplos de programas de domesticación en el mundo (Leakey et ál. 2012). Los ejemplos anteriores ilustran una serie de componentes que dichos programas suelen compartir. Algunos de estos componentes son los siguientes: Por lo general, los programas de domesticación no se limitan al mejoramiento genético, aunque éste es un componente importante y esencial —la domesticación no es sinónimo de mejoramiento genético. Los programas de domesticación no se limitan a las especies silvestres, ya que existen varios grados de domesticación y los requerimientos humanos cambian. Los componentes no genéticos de los programas de domesticación incluyen el manejo (hortícola/silvícola), pero también abarcan todos los aspectos de la cadena de valor y las barreras para su eficaz funcionamiento. Estos pueden incluir, por ejemplo, métodos de procesamiento; embalaje y comercialización; formación de vínculos con el 31 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 sector privado; compensaciones de requisitos de importación y exportación; etc. Sin un enfoque amplio, la domesticación podría tener poco impacto más allá de la generación de documentos científicos. Desafíos La domesticación de árboles es una opción de investigación para el desarrollo emocionante, que parece ofrecer una oportunidad significativa para la investigación en innovación e impacto. Sin embargo, también existen desafíos. A continuación, se mencionan tres de éstos desafíos. ¿Cuándo funcionará? El ser humano depende de un grupo relativamente pequeño de plantas y animales domesticados para su sustento. Además, solo un grupo reducido se comercializa ampliamente. Esta situación ha sido una de las justificaciones para domesticar ‘nuevas’ especies. Sin embargo, se ignora el hecho de que ya hay decenas de especies que están, hasta cierto punto, domesticadas, pero para las cuales no hay mercado. Esto sugiere que es necesario pensar con cuidado antes de embarcarse en programas de domesticación de especies silvestres o, al menos, en programas cuyos productos se comercializan muy poco. Lo anterior suele indicar que los mercados actuales ya son sumamente competitivos. Por ejemplo, una ‘nueva’ especie de nuez tendría que competir en precio y calidad contra varias alternativas, ya sea en mercados de gran escala (almendra, maní) o selectos (macadamia). Del mismo modo, a veces las frutas nativas no podrían competir en precio con alternativas importadas y producidas agroindustrialmente (Clement et ál. 2007). Aunque probablemente falta experiencia a nivel mundial en programas de domesticación para poder evaluar los éxitos y fracasos a la fecha, una ayuda para la toma de decisiones con base en principios a priori podría ser útil para orientar las decisiones de inversión. Lo anterior también debería tomar en cuenta la posibilidad de que haya ‘ganadores y perdedores’ en la domesticación. Por ejemplo, la evidencia (i.e., práctica de abastecimiento de la industria) sugiere que la producción de camu-camu basada en la recolección de poblaciones naturales en Loreto, Amazonía peruana, ha disminuido significativamente debido a la producción de árboles plantados en Ucayali (Bruno Paino, ICRAF, comunicaciones personales). Por lo menos, la domesticación de los actuales productos forestales no maderables que podrían contribuir a generar ingresos para las poblaciones humanas vulnerables debe planearse cuidadosamente, para que no beneficie a los agricultores más ricos en lugar de a las poblaciones meta. Recursos genéticos agroforestales La domesticación no es sinónimo de mejoramiento genético, pero este último es un componente intrínseco y necesario de la mayoría de los programas, y el mantenimiento de la diversidad genética es importante para la producción sostenida de cualquier especie de cultivo. Algunos recursos genéticos agroforestales no están amenazados (sobre todo aquellas especies invasoras que florecen en paisajes antropogénicos dentro y fuera de su rango nativo). No obstante, muchos sí están amenazados (incluyendo variedades locales y poblaciones silvestres de especies domesticadas, y poblaciones de especies silvestres), principalmente, por la destrucción de su hábitat. Una amenaza subyacente y menos obvia es la brecha de conocimiento que existe en relación con la diversidad genética de la mayoría de las especies agroforestales más usadas; en muchos casos, se desconoce la magnitud y distribución de la variación, incluso con especies domesticadas. Enfoques para ensayos genéticos 32 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 En general, los enfoques para ensayos genéticos agroforestales (por ejemplo, ensayos de campo de procedencia y descendencia) parecen seguir la práctica forestal y hortícola tradicional, con pocos intentos por ensayar tratamientos en las condiciones ambientales y de manejo de los sistemas agroforestales en los que podrían utilizarse. Quizás esto se deba a la falta de conexión entre los investigadores involucrados en la domesticación (que suelen conocer de mejoramiento de árboles) y aquellos involucrados en los aspectos silviculturales/hortícolas. Esto también se aborda en la sección final (abajo). ¿Incorporación (“mainstreaming”) de domesticación de árboles... o “mainstreaming” de integración? ¿Podría el enfoque de domesticación de árboles ser un modelo para la investigación agroforestal orientada a la producción a nivel de parcela? Obviamente, no siempre: hay bastante investigación en ecología de producción agroforestal con base en sistemas, que no depende necesariamente de la especie, y que es primordial para la domesticación de árboles. Sin embargo, el resultado deseado de muchas investigaciones con base en la especie es mejorar la calidad o cantidad del producto final. En este caso, se desea un enfoque integrado, es decir, uno que reconozca que existen diversas maneras de lograr los resultados de investigación (por ejemplo, selección genética, manejo hortícola o silvícola, manejo poscosecha, técnicas de procesamiento) y que éstas, aunque sean complementarias, tienen distintas relaciones de costo-beneficio; por lo tanto, necesitan priorizarse y valorarse de forma distinta. En muchos casos, podría no tener mucho sentido calificar a este enfoque como ‘domesticación de árboles’; por ejemplo, con especies que ya se plantan ampliamente. Sin embargo, un enfoque que integre investigación silvícola/hortícola, información de mercado y mejoramiento genético mutuamente complementario en las prácticas de mejoramiento agroforestal (cualquiera que sea el nombre que se use) pareciera ser una opción razonable para todas las principales especies agroforestales. Actualmente, no siempre se sigue este enfoque. Referencias Clement C, Cornelius JP, Pinedo Panduro MH, Yuyuma K. 2007. Native fruit tree improvement in Amazonía: A overview. Pp 110-119 in Akinnifesi FK, Leakey RRB, Ajayi OC, Sileshi G, Tchoundjeu Z, Matakala P. y Kwesiga FR (Eds.). Indigenous fruit trees in the tropics: domestication, utilization and commercialization. CAB International Publishing,Wallingford, UK. 2008. ISBN 978-1-84593-110-0. Cornelius, JP, Leakey RRB, Minnah S Nevenimo T Page T, Pauku R, Sibaris W, Yagau K. 20120. Domestication and commercialisation of Canarium indicum in Papua New Guinea. Informe final para ACIAR (FR2012-21). ISBN 9781921962912. Leakey RRB, Weber JC , Page T , Cornelius JC, Akinnifesi FK, Roshetko JM, Tchoundjeu Z, Jamnadass R. 2012 Tree Domestication in Agroforestry: Progress in the Second Decade (2003–2012). Pp 145-173 in Nair PKR y Garrity D. (eds) 2012. Agroforestry – the future of global land use. Springer. NARI. 2013. Development of galip-nut industry. http://www.nari.org.pg/node/272. Accedido el 7 de noviembre de 2013. Pye-Smith C. 2014. Trees for Life. ICRAF, Nairobi. Simons AJ and Leakey RRB. 2004. Tree domestication in tropical agroforestry. Agroforestry Systems 61: 167-181. 33 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Wallace H, Carter J, Austin J, Poienou M, Moxon J, Walton D, Randall B, Viji I. 2012. Processing of Canarium indicum nuts: adapting and refining techniques to benefit farmers in the South Pacific. Final report to ACIAR (FR2012-12). ISBN 978 1 921962 68 4 34 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 PATRONES Y DETERMINANTES HISTÓRICOS Y GEOGRÁFICOS DE LA DEFORESTACIÓN EN EL ECUADOR, 1950-2010 Y TENDENCIAS AL FUTURO.1 Rodrigo Sierra, Ph.D. GEO-15 [email protected] Esta es una aproximación a los patrones, factores y agentes de la deforestación en el Ecuador continental que abarca la segunda mitad del siglo pasado y la primera década del presente y una exploración de sus tendencias futuras. El análisis desagrega la deforestación neta en sus componentes básicos: la deforestación bruta asociada con el establecimiento de nuevas áreas agropecuarias e infraestructura, como carreteras y asentamientos humanos, y la regeneración de bosques naturales a partir de áreas agropecuarias abandonadas o la recuperación de bosques muy degradados. En el período 1990‐2008 se perdieron aproximadamente 19000 kilómetros cuadrados del bosque nativo en el Ecuador Continental.2 Esta área equivale al 13% del área forestal remanente en 1990. El 99.4% del área deforestada entre 1990 y 2000 fue transformada a áreas agropecuarias, el 0.14% a infraestructura, principalmente áreas urbanas y asentamientos rurales densos, y 0.46% a otros tipos de cobertura. Entre el 2000 y el 2008, el 99.4% del área deforestada fue transformada a áreas agropecuarias, el 0.23% a infraestructura, principalmente áreas urbanas y asentamientos rurales densos, y 0.37% a otros. La expansión del área agropecuaria total también dependió de la deforestación: aproximadamente el 97.5% y el 95% del incremento del área agropecuaria entre 1990 y 2000 y 2000 y 2008, respectivamente, se generaron mediante la transformación de áreas de bosque a cultivos y pastos. Es razonable asumir que la relación entre la expansión del área agropecuaria y del área deforestada ha sido históricamente estable. Usadas como proxi, esta relación permite distinguir tres periodos con características de expansión del área bajo uso, y por lo tanto del área deforestada, y condiciones estructurales particulares: Un periodo inicial, desde mediados de los 1950s y hasta los primeros años de la década de los 1970s, de expansión regional del área agropecuaria, y por lo tanto de la deforestación, en la costa centro y norte y en la amazonía al pie de los andes, y de contracción del área agropecuaria, y por lo tanto presumiblemente de recuperación de bosques, en la sierra. En este periodo la densidad de población rural creció rápidamente, creando presión sobre el área productiva especialmente en la sierra, donde la disponibilidad de tierras productivas era bastante limitada. El segundo periodo comienza a inicios de la década de los 1970s, cuando se acelera significativamente la expansión del área agropecuaria en todo el país, incluyendo en la sierra, pero con densidades de población decrecientes por la caída de las tasas de crecimiento del población nacional en general y de las poblaciones rurales en particular, y por la extensificación de la producción agropecuaria mediante la transformación de espacios forestales con densidades de Este documento se basa en Sierra (2013), Castro et al. (2013) e investigaciones en proceso por el autor. Colaboraron en varias etapas de este trabajo Oscar Calva, Miguel Castro, Jaime Camacho y Fausto López. Estos estudios fueron financiados por Conservación Internacional-Ecuador, el Programa Gestión Sostenible de Recursos Naturales (GESOREN) de la Deutsche Gesellschaft für Internationale Zusammenarbeit (GIZ) GmbH, el Programa Nacional Conjunto ONU-REDD y el Ministerio del Ambiente del Ecuador. El autor agradece este apoyo. 2 Calculado en base a MAE (2012) con modificaciones por R Sierra. 1 35 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 población bajas, especialmente en áreas ocupadas pero no controladas por comunidades indígenas y negras en la costa y la amazonía. El tercer período, correspondiente aproximadamente a los últimos 10‐15 años del siglo pasado y a la primera década del presente (y posiblemente continuando en los próximos años), es un periodo caracterizado por la intensificación de la deforestación de los remanentes de bosque inmersos en las áreas agropecuarias existentes y en desarrollo, sin una expansión importante hacia nuevas áreas con cobertura forestal continua, y densidades rurales decrecientes por la concentración de la población rural en asentamientos rurales densos a lo largo de las carreteras o su migración a ciudades y al exterior, y la profundización de la transición demográfica del país. Durante este último periodo, la mayor parte de la deforestación neta, cerca del 70%, ocurrió en la década de los 1990s, apuntando a una tendencia hacia la caída de la deforestación en el país. Aproximadamente el 60% de la reducción observada se debe a un aparente incremento de la regeneración de bosques, aunque al menos una parte de las áreas de recuperación pueden corresponder a plantaciones forestales o ciertos cultivos perennes maduros, como café. La diferencia (~40%) se explica por la caída de la demanda de nuevos espacios para ser dedicados a usos agropecuarios e infraestructura (Tabla 1).3 Estudios recientes sugieren además que estas tendencias son similares a las de otros países de la región (e.g., Aide et al. 2012, Sánchez et al. 2012, INPE 2013). A nivel regional (Costa, Sierra, Oriente), el área deforestada neta cayó en todo el país. A nivel provincial, solo Morona Santiago y Bolívar experimentaron incrementos en la deforestación neta en la última década. En el resto de provincias la deforestación neta anual cayó entre el 5% (Santa Elena, Pastaza) y el 80% (Sucumbios y Orellana). De los 10 cantones con el mayor incremento absoluto del área deforestada, cuatro están en Manabí, tres en Morona Santiago, y uno en Bolívar, Esmeraldas y Guayas. De los diez cantones con mayor caída en la deforestación absoluta, cuatro están en Esmeraldas, dos en Manabí, dos en Orellana y dos en Sucumbios. Los ecosistemas forestales más afectados son los bosques húmedos de las cordilleras de la costa, donde se observan entre el 2000 y el 2008 las tasas de deforestación anual promedio más altas del país y una tendencia hacia la aceleración de la deforestación. La deforestación de los otros tipos de bosque ha bajado pero se mantiene alta en los bosques húmedos y secos y semi‐secos de la costa y los bosques del piedemonte andino en la costa. Cuatro factores estructurales son claves para entender los patrones de la deforestación en el Ecuador: 1) la intensificación de los sistemas productivos rurales y la reestructuración del empleo rural, 2) las mejoras en accesibilidad en casi todos los sectores rurales del país, 3) la caída de las tasas de crecimiento de población en general, y de las de natalidad en particular, en todos los segmentos de la población ecuatoriana, y 4) el cierre de casi todas las fronteras de colonización debido a la consolidación de los derechos de propiedad de las áreas forestales remanentes más importantes. Éstos definen los niveles de presión para crear nuevos espacios productivos y la permanencia de las aéreas bajo usos agropecuarios. La combinación de mejoras en la accesibilidad a áreas urbanas, la caída de las tasas de natalidad y el incremento de los ingresos urbanos y rurales no agropecuarios, impulsa la intensificación productiva del sector agrícola. Este análisis no evalúa la degradación de bosque, y en especial la tala comercial en bosques nativos. Sabemos que solo una pequeña fracción de los bosques explotados son luego reemplazados por áreas agropecuarias. Como resultado de esto, una fracción importante del área forestal medida corresponde a bosques degradados. 3 36 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figura 1. Patrones de deforestación provincial y regional en el Ecuador continental, 1990-2000 y 2000-2008. (Fuente: Sierra 2013; Modificado de MAE 2012) 37 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Donde ocurre, la intensificación se evidencia en el incremento del uso de insumos agropecuarios y de las productividades por unidad de área bajo uso (Figura 2). Esto ha ocurrido con los cultivos de mayor expansión en el país: arroz, maíz, cacao, palma africana y caña de azúcar. Entre 1990 y 2010 la productividad agrícola creció en forma continua sin una mayor expansión del área bajo uso.4 Donde no ocurre, como en las zonas ganaderas cuya productividad, medida como carga por unidad de área, se mantuvo estable entre 1990 y 2008, la producción total se incrementa mediante la expansión del área bajo producción. La reestructuración del empleo y redistribución de la población rural se evidencian en la variación en el crecimiento y proporción de la población económicamente activa (PEA) primaria (trabajadores que labora en agricultura, ganadería y silvicultura) y la creciente concentración de la población en asentamientos rurales densos a lo largo de carreteras y ciudades menores, sobre todo a partir de inicios de este siglo. Dentro de este contexto general funcionan factores coyunturales que afectan las dinámicas locales y a corto plazo de la deforestación. En especial, factores que modulan la demanda de nuevos espacios agropecuarios 120.00 110.00 INDICE SUPERFICIE AGROPECUARIA 100.00 90.00 80.00 ÍNDICE PRODUCCIÓN AGRÍCOLA NETO PER CÁPITA 70.00 2010 2009 2008 2007 2006 2005 2004 2003 2002 2001 2000 1999 1998 1997 1996 1995 1994 1993 1992 1991 1990 1988 1987 1974 1964 60.00 Figura 2. Índices de Superficie y Producción Agrícola per cápita de Ecuador. Fuente: FAO (2013); INEC (2009); SICA (2002) en Castro et al. 2013. están íntimamente relacionados con los ciclos económicos del país. Cuando el crecimiento económico se acelera, la deforestación y degradación de bosques repunta, y cuando cae, caen también las tasas de deforestación y degradación de bosques. La relación es clara. Cuando la economía del país crece, la población tiene recursos para comprar más productos que se producen creando nuevos espacios agropecuarios, como carne en Morona Santiago, o puertas, sillas y mesas que se producen mediante la tala de bosques naturales.5 En general, comparando las tasas de deforestación anual en el periodo 1990-2000 con el período 2000-2008, en las regiones donde aumentó o se mantuvo la tasa de deforestación se producen esencialmente productos agropecuarios destinados al consumo interno. Es decir, fue la propia demanda interna, motivada por una creciente economía, ingresos y consumo, el principal incentivo a deforestar en Ecuador. La demanda nacional de maíz, caña de azúcar, arroz y yuca fueron factores claves en la deforestación en la Amazonía Centro, las Cordilleras y Valles Semi-secos de la Costa Central, los Bosques y Valles Semisecos del Sur y la Sierra Centro. La creciente demanda interna de carne fue la principal presión en la Amazonía Sur, Manabí NorteEl área sembrada y la productividad bajó en este mismo período en algunos productos importantes, por ejemplo la papa y el fréjol, pero la producción de éstos no está relacionada con procesos de deforestación. 5 Esto ocurre a pesar de que hay suficiente evidencia como para asumir que una parte significativa de la madera consumida en el Ecuador en los últimos años provino de plantaciones forestales. 4 38 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Esmeraldas Sur y Manabí Centro. Por otra parte, una constante demanda de leche incidió en la deforestación en el Piedemonte Andino de la Costa Central, las Estribaciones Occidentales de los Andes del Sur y la Sierra Norte. La producción de cultivos permanentes como cacao, palma africana y banano, destinados mayoritariamente al comercio internacional, son los principales promotores en la Amazonía Norte, Esmeraldas Norte y Piedemonte Costa Norte y en la Cuenca del Guayas. Tanto la deforestación como la regeneración se encuentran espacialmente concentradas. La deforestación 2000-2008 ocurrió en 250 mil parches cuyos tamaños van de <1 ha hasta más de 3.000 has. Los parches de <1 ha son el 65% pero apenas representan el 4% de la superficie afectada. Los parches mayores a 50 has son menos del 2% del total pero concentran el 42% de la superficie deforestada. El 72% de los 233 mil parches de regeneración tienen menos de 1 ha pero apenas representan el 9,4% de la superficie boscosa recuperada, mientras que los parches mayores a 50 has, que son menos del 1%, concentran el 31% de esta superficie. Las proyecciones de la deforestación en la próxima década sugieren que la caída de la deforestación en el Ecuador continuará a largo plazo, aunque es razonable esperar que no sea distribuida en forma homogénea en todo el país y que no varíe a corto plazo con las fluctuaciones de la economía nacional. Los modelos proyectan una reversión del proceso en dos zonas: en Manabí Central y en los Bosques y Valles Semi-secos del Sur; en los dos casos principalmente debido a una redistribución de la población. En la primera ya se empezaron a observar indicios de una transición forestal durante la primera década de este siglo y en la segunda la deforestación cayó en más de 50% en este periodo comparado con la década anterior. En contraste, la proyección de la deforestación en el norte de la Amazonía muestra una tendencia a incrementarse, principalmente debido a la caída de los costos de transporte y mejoramiento de la renta agropecuaria. Los resultados de los modelos no deben ser tomados como medidas precisas, sino más bien como indicadores de las tendencias de presión. En estas condiciones, los mosaicos de remanentes de bosque con agricultura/ganadería que se formaron en las seis décadas pasadas pueden transformarse paulatinamente en paisajes estrictamente agropecuarios. A nivel nacional, la tendencia apunta a la formación de paisajes bipolares: unos completamente dominados por bosques, lejos de las áreas donde vive la gente, y otros completamente agropecuarios en y alrededor de donde vive la gente. En su conjunto, el análisis de promotores regionales y las proyecciones estadísticas para cada una apuntan a la importancia de las políticas de Estado en la gestión del uso y cobertura del suelo. Los planes de desarrollo del Estado, que afectan tanto a los costos de oportunidad del bosque, mediante el incremento de la demanda nacional (a su vez relacionada con los niveles de ingresos reales de la población) y la apertura potencial de mercados de productos relacionados a la deforestación, como a la dinámica demográfica del país, juegan un papel clave en la estructuración del riesgo de deforestación. Específicamente, este estudio demuestra que es posible reducir la deforestación, en especial en aquellas zonas con niveles altos de expansión del área de uso agropecuario mediante políticas que promuevan la intensificación agropecuaria en general, y la ganadería en particular, sobre todo si se considera que todavía existe una brecha entre los rendimientos agrícolas de Ecuador y los rendimientos internacionales. Políticas de estado que promuevan la concentración y estabilización de la población en asentamientos densos, especialmente rurales, mediante mejoras de viabilidad o servicios públicos, podrían reducir la presión para crear nuevos espacios productivos en áreas alejadas. Para ello el Estado debe promover la implementación de políticas públicas que incentiven el acceso 39 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 a asistencia técnica, insumos y semillas mejoradas, crédito y que promuevan la intensificación del uso eficiente del espacio agrícola. Los modelos de riesgo de deforestación demuestran además que las acciones encaminadas a la protección de los remanentes de bosque tiene un impacto clave en su conservación. El establecimiento de áreas protegidas nacionales por el Ministerio del Ambiente del Ecuador y de bosques protectores por gobiernos locales ha sido un factor de freno de la deforestación, especialmente en la región andina. Esto sugiere que una política que promueva las acciones de protección de los gobiernos autónomos descentralizados provinciales y municipales, puede dar resultados significativos en el proceso de reducir la deforestación, especialmente en zonas con pocos remanentes de bosque primario. Referencias Aide, T., M. Clark, H. Grau, D. López, M. Levy, D. Redo, M. Bonilla, G. Riner, M. Andrade, y M. Muñiz (2012). Deforestation and Reforestation of Latin America and the Caribbean (2001–2010). Biotropica 0(0): 1–10 (10.1111/j.1744‐7429.2012.00908.x) Castro, M., R. Sierra, O. Calva, J. Camacho, F. López y P. Lozano. 2013. Zonas de Procesos Homogéneos de Deforestación del Ecuador. Factores promotores y tendencias al 2020. Programa GESOREN-GIZ y Ministerio de Ambiente del Ecuador. Quito, Ecuador. FAO (Food and Agriculture Organization of the United Nations) (2013). http://faostat.fao.org/ Faostat pilot version. Disponible en febrero 12, 2013. INEC (Instituto Nacional de Estadísticas y Censos) (Varios años) Encuesta de superficie y producción agropecuaria continua (ESPAC). Instituto Ecuatoriano de Estadísticas y Censos INEC, Quito. INPE (2013). http://www.obt.inpe.br/prodes/index.php. PRODES. Disponible 01 Enero 2013. MAE (Ministerio de Ambiente del Ecuador) (2012). Mapas de Deforestación Histórica del Ecuador. Versión Mayo 2012. Ministerio de Ambiente del Ecuador, Quito. Sánchez, A., T. Aide, M. Clark y A. Etter. (2012) Land Cover Change in Colombia: Surprising Forest Recovery Trends between 2001 and 2010. PLOS 7:e43943 (SICA) Proyecto Servicio de Información y Censo Agropecuario, Instituto Ecuatoriano de Estadísticas y Censos (INEC) Ministerio de Agricultura y Ganadería (MAG) (2002). III Censo Nacional Agropecuario, in Instituto Ecuatoriano de Estadísticas y Censos (INEC), ed.: (Quito). Sierra, R. 2013. Patrones y factores de deforestación en el Ecuador continental, 1990-2010. Y un acercamiento a los próximos 10 años. Conservación Internacional Ecuador y Forest Trends. Quito, Ecuador. 40 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 BOSQUES Y CAMBIOS CLIMÁTICOS Franklin Cargua CUANTIFICACIÓN DE STOCK DE CARBONO EN UNA PLANTACIÓN DE PINO (Pinus radiata) EN ESTRATO DE PÁRAMO DE OZOGOCHE BAJO, PARQUE NACIONAL SANGAY Jesús Mª Bastida EFECTO DE LA FRAGMENTACIÓN DEL BOSQUE TROPICAL SECO SOBRE LA VIABILIDAD DE PROGENIES DEL ÁRBOL DIOICO Brosimum alicastrum (MORACEAE) Jimmy J. Cevallos Zambrano EVALUACIÓN ECOLÓGICA RÁPIDA DE LA FAUNA EXISTENTE EN EL BOSQUE TROPICAL SIEMPRE VERDE DEL CANTÓN EL CARMEN, MANABÍ ESTRUCTURA Y COMPOSICIÓN DEL BOSQUE SECO SEMIDESIDUO DE LA RESERVA DE BOSQUE SECO LALO LOOR DEL CANTÓN JAMA, MANABÍ ESTRUCTURA Y COMPOSICIÓN FLORÍSTICA DEL BOSQUE TROPICAL SIEMPRE VERDE DEL CANTÓN EL CARMEN, MANABÍ Jorge Caranqui CONTRIBUCIÓN A LA DIVERSIDAD Y SIMILITUD ARBOREA DE LOS BOSQUES MONTANOS ALTOS DEL ECUADOR CENTRAL DIVERSIDAD Y COMPOSICIÓN FLORÍSTICA EN LA ESTACIÓN CIENTÍFICA RÍO PALENQUE Fabián Salas CARACTERIZACIÓN DE LAS ÁREAS NATURALES (BOSQUES Y PÁRAMOS) COMO HERRAMIENTA DE ORDENAMIENTO TERRITORIAL EN LA PROVINCIA DE CHIMBORAZO 41 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 CUANTIFICACIÓN DE STOCK DE CARBONO EN UNA PLANTACIÓN DE PINO (Pinus radiata) EN ESTRATO DE PÁRAMO DE OZOGOCHE BAJO, PARQUE NACIONAL SANGAY. Franklin E. Cargua; Marco V. Rodríguez; Celso G. Recalde; Luis M. Vinueza Caracterización Biogeográfica de Sub-cuencas hídricas; UNACH; Av. Antonio José de Sucre Km 11/2 vía a Guano; Riobamba-Ecuador. [email protected] RESUMEN El objetivo de la presente investigación determinó el contenido de carbono (C) acumulado en una plantación de pino (Pinus radiata) y en un estrato de páramo, ubicado en la zona de amortiguamiento sur occidental del Parque Nacional Sangay; se analizó el contenido de carbono en (biomasa aérea y subterránea, sotobosque, necromasa y suelo). Se establecieron 6 parcelas de monitoreo, el contenido de carbono promedio en biomasa en la plantación fue de 14.69 Mg C/ha, para sotobosque el valor promedio fue del 0.68 Mg C/ha, en plantación y 6.55 Mg C/ha en páramo; para detritus no vivo se obtuvo un valor de 2.97 Mg C/ha en plantación y 56.09 Mg C/ha en páramo; el carbono en suelo fue de 268.83 Mg C/ha e la plantación y 215.0 Mg C/ha en páramo y el carbono fue de 287.31 Mg C/ha en la plantación y 277.81 Mg C/ha en páramo. Palabras clave: Stock de carbono, suelo, biomasa, necromasa, plantación forestal, páramo. INTRODUCCIÓN El páramo es un ecosistema neo tropical ubicado en Centro y Suramérica, en la franja entre el bosque montano y el límite superior de la nieve perpetua, es decir, aproximadamente entre 3.000 y 5.000 msnm, (Sklenář et al., 2005). Se caracteriza por ser un ambiente frío y húmedo, con temperaturas diarias que fluctúan entre los -3°C y +20°C (Mena y Balslev, 1986) y una precipitación anual que varía desde 500 hasta sobre 3.000 mm (Luteyn, 1999). En términos biológicos, los páramos constituyen una parte importante de la extraordinaria diversidad ecológica de un país relativamente pequeño como el Ecuador, (Medina et al., 1997; Medina, 2000; Medina y Mena, 2001), pero con una variedad ambiental y biológica mayor a la de países con extensiones muy superiores (Mittermeier et al., 1997). Los suelos de los páramos del Ecuador se han desarrollado esencialmente sobre los depósitos piroclásticos resultantes de las erupciones de los numerosos edificios volcánicos (Quantin y Zebrowski, 1997). Se trata en general de andosoles, pero su morfología y propiedades varían considerablemente según los principales factores de la pedogénesis, como la edad, naturaleza, composición química de los materiales y las condiciones climáticas. La acumulación de carbono orgánico en el suelo (OTCS) es un proceso importante para mitigar efectos del cambio climático (Houghton & Woodwell, 1989), ya que el suelo, además de ser un sumidero, es un reservorio de carbono estabilizado (Sánchez et al., 2011). Otro de los ecosistemas que ayudan al secuestro de carbono 42 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 son los bosques, con un manejo adecuado, pueden secuestrar más carbono que otros ecosistemas terrestres, contribuyendo a la mitigación del cambio climático, producido por la degradación y deforestación, de esta manera se considera que los bosques podrían ser considerados como sumideros de C, y los dueños podrían recibir un pago por la venta de bonos de C en base al acuerdo de los diferentes protocolos internacionales e iniciativas nacionales (Masera, 1995; Návar, 2009). Por otra parte, después del Protocolo de Kyoto (1997), la reducción de gases de efecto invernadero ha llegado a ser una prioridad para la mayoría de los países (Hernández et al., 2006), las actividades forestales como la reforestación, forestación y la protección de áreas forestales (Vesterdal et al., 2007), pueden ser implementadas para mitigar el calentamiento global (Hernández et al., 2006). Una forma de mitigar los efectos negativos que genera el CO2 es capturarlo y mantenerlo el mayor tiempo posible en la biomasa de las plantas (Pauli et al., 2003) y en el suelo (Torres et al., 2011). Por esta razón se han implementado políticas gubernamentales y los mercados de carbono están proporcionando mecanismo de flexibilidad existente; siendo uno de ellos el proyecto llamado mecanismo de desarrollo limpio (Concha et al., 2007), como forma de compensar a los países que protegen sus bosques y aquellos que reducen las emisiones (Torres et al., 2011). La estimación precisa de la dinámica de los flujos netos de carbono entre los bosques y la atmósfera (es decir, el balance emisión-captura) es uno de los problemas abiertos más importantes en la discusión sobre cambio climático (IPCC, 2005). El objetivo del presente trabajo fue estudiar el almacenaje de carbono en los cinco depósitos propuesto por el IPCC (2006), en una plantación de pino (Pinus radiata), establecida en un ecosistema de páramo (Hofstede, 2000). MATERIALES Y MÉTODOS Área de Estudio La presente investigación se llevó a cabo en la comunidad de Ozogoche Bajo, que se encuentra en el flanco nororiental de la zona de amortiguamiento del parque Nacional Sangay en la parroquia Achupallas, cantón Alausí, provincia de Chimborazo, en una plantación de pino (Pinus radiata) que abarca una superficie de 100 ha; las muestras se tomaron en rangos de altitud desde los 3792 - 4096 msnm. Los datos obtenidos de los anuarios de la estación meteorológica (M396 INHAMI 2010); presentan una temperatura media de 8 ºC, con rangos de (4 - 12) °C; una precipitación de 1077 con rangos de (1000 – 1200) mm. Los tipos de vegetación presentes en esta zona son propios de estratos de páramo (IPCC 2006), en la zona de estudio podemos encontrar especies endémicas (Cortaderia nítida), (Calamagrostis intermedia), Almohadillas (Hypochaeris sessiliflora), (Wernerianubigena), (Prieto y Paucar, 2011; Luteyn, 1999; Jorgensen & León-Yánez, 1999; Mena-Vásconez, 2001; León-Yánez, 2000), para la investigación se ha considerado la comparación del contenido de carbono en una plantación de pino (Pinus radiata), en estado fustal de 18 años de edad, establecido a una altura de 3840 msnm; y el contenido de carbono en un ecosistema de páramo conservado ubicado a una altura de 4000 msnm. 43 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Muestreo Se aplicó un diseño de muestreo de doble estratificación, en dos fases con una rejilla grande de (1x1) km con proyección geográfica UTM WGS´84 Zona 17S; path-row 10-62(glovis.usgs.gov), luego se seleccionó una muestra más pequeña en dos estratos (Fig. 1); (Cueva et al., 20012). Figura 1. Ubicación de las parcelas de monitoreo georeferenciadas Estratificación de unidades de muestreo En cada muestra seleccionada se midieron y observaron todas las variables de interés a través de las parcelas de muestreo, y a partir de ellas se derivaron las estimaciones de cada variable por estrato (Carvajal et al., 2009). Ecuación 1. Para determinar (i) intensidad de muestra. Para las mediciones de campo se utilizaron un conglomerado de parcelas en forma de L (Figura 2). Las parcelas fueron de forma cuadrada de 60 x 60 m. Esta forma de parcelas es usual para las mediciones de biomasa y la mayoría de tipos de vegetación (Ravindranath y Oswald, 2008). Tabla1. Localización de las Parcelas Permanentes de Muestreo (PPM). Conglomerado Parcela CUT`s Descripción P1 20 Páramo conservado 001 P2 20 Páramo conservado P3 20 Páramo conservado P1 10 Plantación (Pinus radiata) 002 P2 10 Plantación (Pinus radiata) P3 10 Plantación (Pinus radiata) Dónde: P= parcela 44 Este 765171 764921 764921 761931 761681 761681 Norte 9749347 9749347 9749597 9754976 9754976 9755227 Altitud (msnm) 4096 3984 4020 3796 3880 3876 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 En la parcela de 60 x 60 m, se realizó un inventario forestal, se establecieron sub-parcelas para recolectar muestras de suelo a diferentes profundidades: entre (0-10), (10-20), (20-30) cm; se implementó otra subparcela donde se registraron los pesos de la hojarasca y detritus no vivo. La tercera sub-parcela permite la medición de sotobosque. Figura 2. Diseño, distribución y tamaños de las parcelas anidadas para las mediciones. RESULTADOS Y DISCUSIÓN Stock de carbono en la biomasa aérea y subterránea en la plantación de (Pinus radiata) Para la transformación del volumen de fuste estimado mediante inventarios forestales, en valores de biomasa total son necesarios coeficientes técnicos, que representan la relación porcentual entre ambas medidas. Para determinar el contenido de biomasa aérea, se utilizando factores de expansión (Loguercio & Defossé, 2001), para el caso de la raíz 0.27, para fuste + copa 2.8 y densidad de la madera 0,54 g/cm3 (Kyrklund, 1990). A partir de ellas se ha generado mucha información que constituye una base muy útil a la hora de realizar estimaciones de la capacidad de fijación de carbono en un bosque. Se ha realizado estudios especializados que han determinado factores estándar, los cuales nos han servido para determinar el contenido de carbono fijado en la plantación (Cueva et al., 2012). La variación del contenido de carbono orgánico total está representada por clases diamétricas en la plantación de pino, con un valor promedio de 0,20 MgC/ha con niveles de confiabilidad de 0,4-0,1 y una desviación estándar de ±0,14, los valores de biomasa aérea más alta se observaron en la clase diamétrica (>20-25) cm, con un valor de 0,32 y una desviación de ± 0,04. Los valores más altos de biomasa subterránea se observaron en la clase diamétrica (>20-25) cm, con un valor medio de 0,09 con una desviación estándar de ±0,01; otros estudios sobre la biomasa de raíces de coníferas muestran valores entre 0,1 y 0,25 del árbol (Guerra et al., 2005) 45 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Tabla 3. Contenido de carbono aéreo en la plantación de Pino (Pinus radiata) Conglomerado Parcela Carbono (Mg/ha) P1 16,33 002 P2 11,03 P3 16,72 Tabla 4. Relación entre biomasa aérea y subterránea de la plantación de Pinus radiata. Clase diamétrica Biomasa Biomasa Biomasa N0 Carbono Total (cm) aérea subterránea Total 1 10 – 15 0.09 0.02 0.11 0.06 2 >15 – 20 0.22 0.06 0.28 0.14 3 >20 – 25 0.36*±0.04 0.10*±0.01 0.46*±0.05 0.23*±0.02 4 >25 – 30 0.62 0.17 0.78 0.39 * Media ± Coeficiente variación. DAP: Diámetro a la altura del pecho (1.30m) (Lazore y Gilabert, 2007), menciona dentro de sus estudios la relación entre raíz/fuste fue de 20,3 y 79,7% respectivamente, además se hace énfasis en el desarrollo de la plantación que varía con el tiempo pero a determinada edad el incremento se detiene. Figura 4. Relación entre biomasa aérea y biomasa subterránea. Carbono presente a diferentes profundidades en las parcelas permanentes de muestreo (PPM). El contenido de carbono en el suelo fue mayor a una profundidad de (20-30) cm; en la plantación de Pinus radiata con un valor medio de 85.54 MgC/ha con desviación estándar de ± 13.28; mientras que el valor más bajo se observa en el estrato de páramo con un valor de 65.01 MgC/ha y una desviación estándar de 9,22; estudios similares (Ibrahim et al., 2007) , reportan valores entre 52,3 ± 9,8 y 81,3 ± 3,6 Mg/ha en pasturas, lo que difiere de los ecosistemas de páramo por ser un sumidero de carbono. 46 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Tabla 5. Contenido de carbono en el suelo Conglomerado 001 002 Parcela Profundidad Profundidad Profundidad (0-10 cm) (10-20 cm) (20-30 cm) P1 73.15 73.76 71.70 P2 71.67 74.61 69.95 P3 71.41 73.76 65.01 P1 85.55 83.45*±10.30 85.54*±13.28 P2 94.35 99.04 98.19 P3 80.25 *±9.22 88.90 91.22 * Media ± Desviación estándar. Profundidad de suelo. Figura 5. Concentración de carbono acumulado en suelo a diferentes profundidades. Carbono orgánico total en parcelas permanentes de muestreo (PPM). El contenido promedio de carbono en la biomasa aérea en la plantación forestal fue de (14.69 Mg C/ha), sus límites de confianza + 18.63; -10.7. En la tabla 6 muestra la cantidad de carbono presente en los CUTs, evaluados. Como se esperaba en la plantación de pino fue donde se logró el mayor secuestro de C con, (287.31 MgC/ha) en comparación con el observado en el páramo (277.82 MgC/ha). 47 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Conglomerado 001 002 Tabla 6. Contenido de carbono orgánico total presente en los cinco depósitos Biomasa Carbono aérea SOTB DNV Raicillas Suelo CUTs Parcela Total plantación MgC/ha MgC/ha MgC/ha MgC/ha Mg*/ha MgC/ha P1 6,48±0.31 50,46 0,15 218,61 275,70±4.72 Páramo P2 6,29 51,84 0,17±0.02 216,23±4.35 274,53 P3 6,89 65,96±8.58 0,19 210,17 283,22 * Media ± Desviación estándar. Carbono orgánico total en estrato de páramo P1 16,33±3.18 0,39 3,52 0,16±0.04 254,54 274,93 P2 11,03 0,93 2,66 0,10 291,59 306,31 P3 16,72 0,71±0.27 2,72±0.48 0,17 260,37±19.92 280,69±16.7 * Media ± Desviación estándar. Carbono orgánico total en plantación forestal Plantación forestal El contenido promedio de carbono en la biomasa aérea (Ruiz-Peinado et al., 2013), en la plantación forestal fue de 14.69 MgC/ha, sus límites de confianza +18.63 y -10.7; los mismos que están dentro del rango 20,3 y 79,7% respectivamente (Lazore y Gilabert, 2007), además se hace énfasis en el desarrollo de la plantación que varía con el tiempo pero a determinada edad el incremento se detiene. En potreros de Oaxaca se encontraron de 99 a 181 MgC/ha (Etchevers et al., 2002). Para el depósito de sotobosque se obtuvo el valor más alto en el estrato de páramo con una media de 6.55 Mg C/ha y una desviación estándar de 0.31; mientras que para la plantación forestal se obtuvieron los valores de 0.68 Mg C/ha con una desviación estándar de 0.27; reportes similares (Ruiz, 2002), mencionan valores de (12.5±3.6 MgC/ha); la cantidad de carbono acumulado en la materia muerta o detritus no vivo obtuvo el valor más alto en el estrato de páramo con un valor medio de 56.09 Mg C/ha, con una desviación estándar de 8.58, mientras que la plantación forestal tiene un valor medio de 2.97 Mg C/ha, con una desviación estándar de 0.48; (Gasparri y Manghi, 2004), reporta valores de 6.92 Mg C/ha para madera muerta y 2.80 Mg C/ha, para hojarasca. La taza de eficiencia en el ecosistema de páramo fue de 75% de CO2 de la atmósfera (Houghton, 1996), esto implica que la pérdida de biodiversidad florística se afecte al momento de establecer plantaciones exóticas en ecosistemas que son fuente de varios servicios ecosistémicos como agua, carbono, etc. Como se esperaba en la plantación de pino fue donde se logró el mayor secuestro de carbono total con un valor medio de, (287.31 MgC/ha), con un coeficiente de variación de 5.81 y un error relativo del 5%, mientras que los datos obtenidos en el estrato de páramo fueron de (277.82 MgC/ha), con un coeficiente de variación de 4.71 y un error relativo del 1.49%, observando el aporte relativo que prestan las plantaciones exóticas las cuales son aprovechadas en la industria. Los resultados obtenidos en páramo y plantación forestal fueron similares a los obtenidos por (Gayoso & Schlegel, 2001), quien registró estimaciones de 389 y 212 Mg/ha, en bosques ubicados en la Cordillera de los Andes y de la Costa de Chile respectivamente. 48 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figura 6. Relación del C en SOTB/DNV Figura 7. Carbono orgánico total Varios estudios han estimado la acumulación de biomasa total aérea y la consecuente fijación de C en bosques no intervenidos (Krankina et al., 1996), plantaciones forestales y bosques secundarios usando o desarrollando modelos estadísticos, (Brown y Lugo, 1982; Brown et al., 1989; Brown and Lugo, 1990; Brown, and Lugo, 1992; Brown, 1997; Nelson & Sommers, 1996; Dauber, 1995; Schlesinger, 2000). El C orgánico del suelo (Martínez et al., 2008), se encuentra en forma de residuos orgánicos poco alterados de vegetales, animales y microorganismos, en forma de humus y en formas muy condensadas de composición próxima al C elemental (Jackson, 1964; Céspedes, 2012). Como se puede observar, los cálculos de la captura de carbono en plantaciones forestales son muy sensibles a los cambios en la productividad. En el segundo grupo de variables en cuanto a su sensibilidad están la cantidad del carbono en el suelo, la densidad de la madera y el factor de expansión para calcular la biomasa total del árbol. Los suelos ricos en materia orgánica tienen mayor porosidad y por lo tanto retienen un mayor volumen de agua; a medida que la densidad aparente aumenta, se reduce la porosidad del suelo y por lo tanto el contenido orgánico disminuye (Chen y Twilley, 2005; Cuevas et al., 2006). El presente estudio muestra una mayor concentración de carbono en el suelo, estos resultados concuerdan con lo reportado por (Swift, 2001), en la que menciona que en este tipo de depósito contienen más C que la suma existente en la vegetación y en la atmósfera. CONCLUSIONES La proporción entre la biomasa aérea con respecto a la biomasa subterránea muestra una tendencia proporcional del crecimiento de la (raíz/fuste) es del 21 y 79% respectivamente presente en todas las clases diamétricas el crecimiento dependerá de factores externos ya mencionados, y del manejo técnico que se implemente en la plantación. El contenido de carbono acumulado en suelo no es inversamente proporcional con respecto a la profundidad, debido a que en el páramo el ciclo de transformación de los nutrientes es lento debido a las bajas temperaturas, por esta razón en los estratos de páramo existen grandes depósitos de “turba” o materia orgánica en descomposición esto permite considerar que el ecosistema de páramo es un gran depósito de carbono. 49 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 El contenido de biomasa y detritus no vivo es mayor en el ecosistema de páramo, debido a que en estos lugares existe un ciclo continuo que se regenera de forma natural, mientras que al momento de implementar una plantación la pérdida de biodiversidad es alta por la perturbación del medio. El contenido total de carbono es mayor en la plantación, debido al aporte de biomasa, esto contrasta con la pérdida de la biodiversidad en ecosistemas que generan servicios ecosistémicos como es la regulación hídrica. LITERATURA CITADA Brown S., 1997. Los bosques y el cambio climático: el papel de los terrenos forestales como umideros de carbono. In Actas del XI Congreso Mundial Forestal: Recursos Forestales y Arboles. Vol 1. Antalya, Turkia 13–22 October 1997. Brown, S. and A. E. Lugo, 1992. Above ground biomass estimates for tropical moist forests of the Brazilian Amazon. Interciencia 17:8-18. Brown, S. y A.E. Lugo. 1982. The storage and production of organic matter in tropical forests and their role in the global carbon cycle. Biotropica. 14: 161-187. Brown, S., A. J. R. Gillespie, and A. E. Lugo, 1989. Biomass estimation methods for tropical forests with applications to forest inventory data. Forest Science 35:881-902. Brown, S., and A. E. Lugo. 1990. Tropical secondary forest. Journal of Tropical Ecology 6: 1-32. Carvajal A., Feijoo, A., Quintero, H., y Rondón, M., 2009. Carbono orgánico del suelo en diferentes usos del terreno de paisajes Andinos Colombianos.Rev. Cienc. Suelo Nutr. / J. Soil. Sci. PlantNutr. Céspedes Flora E., Fernández, J., Gobbi J., y Bernardis A., 2012. Reservorio de carbono en suelo y raíces de un pastizal y una pradera bajo pastoreo. Facultad de Ciencias Agrarias. Universidad Nacional del Nordeste (UNNE). Pcia. Chaco, República Argentina. Chen, R., y Twilley, R. 2005. A simulation model of organic matter and nutrient accumulation in mangrove wetland soils. Biogeochemistry. 44(1): 93-118. Concha, J., Alegre, J. y Pocomucha V. 2007. Determinación de las reservas de carbono en la biomasa aérea de sistemas agroforestales de teobroma cacao l. en el departamento de San Martìn, Perú Ecología Aplicada, vol. 6, núm. 1-2, diciembre, 2007, pp. 75-82. Cueva K., Añasco M., Ordoñez L., Salazar X., Sánchez G., Cisneros C. y Segura D. 2012 Manual de Campo proyecto Evaluación Nacional Forestal y el programa nacional conjunto ONU – REDD+ del Ministerio del Ambiente del Ecuador. Bajo la cooperación del Programa “Manejo Forestal Sostenible ante el Cambio Climático” FAO Finlandia y el componente ONU REDD FAO. Cuevas, J., Seguel, O., Ellies A., Dorner J. 2006. Efecto de las enmiendas orgánicas sobre las propiedades físicas del suelo con especial referencia a la adición de lodos urbanos. Revista de la Cencia del Suelo y Nutrición Vegetal. 6(2): 1-12. 50 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Dauber, E. 1995. Guía práctica y teórica para el diseño de un inventario forestal de reconocimiento. Santa Cruz, Bol, Proyecto BOLFOR. s.p. Etchevers, J. Vargas, M. Acosta, y A. Velázquez. 2002. Estimación de la Biomasa Aérea Mediante el Uso de Relaciones Alométricas en Seis Especies Arbóreas en Oaxaca, México. Agrociencia Vol. 36 N° 006. pp. 725 – 736. Gasparri, I. y E. Manghi. 2004. Estimación de volumen, biomasa y contenido de carbono de las regiones forestales argentinas. Dirección de Bosques, Secretaría de Ambiente y Desarrollo Sustentable. Unidad de Manejo del Sistema de Evaluación Forestal. Argentina. 26 pp. Gayoso, J. y Schlegel, B. 2001. Proyectos forestales para la mitigación de gases de efecto invernadero. Ambiente y desarrollo 17 (5): 14 – 16. Santiago de Chile, Chile. Guerra C, Javier; Gayoso, Jorge; Schlatter, Juan y Nespolo, Roberto. 2005. Análisis de la biomasa de raíces en diferentes tipos de bosques: Avances en la evaluación de Pinus radiata en Chile. Bosque (Valdivia). 2005, vol.26, n.1, pp. 5-21. Hernández, G., Izquierdo, I., Alguacil, M., Caravaca F., y Roldán A., 2006. Captura de C e indicadores de rehabilitación de suelos de áreas degradadas por la minería en MOA (Cuba). TOMO II, 505-514, Diputación de Badajoz, España. Hofstede, R. 2000. Aspectos técnicos ambientales de la forestación en los páramos. En: La forestación en los páramos. Serie Páramo 6: 41-66. GTP/Abya Yala. Quito. Houghton, R.A. y C.M. Woodwell. 1989. Global climate change. Scientific American 260(4):36-40. Houghton, RA., 1996. Converting terrestrial ecosystems from sources to sinks of carbon. Ambio 25: 267-272. http://www.fao.org/forestry/fma/openforis/en/ 2012, 05/02/2013. http://www.glovis.usgs.gov2012, 16/02/2013. Ibrahim, M., Chacón, M., Cuartas, C., Naranjo, J., Ponce, G., Vega, P., Casasola, F. & Rojas, J. 2007. Almacenamiento de carbono en el suelo y la biomasa aérea en sistemas de uso de la tierra en paisajes ganaderos de Colombia, Costa Rica y Nicaragua. Agroforestería en las Américas, 45: 27–36. Jackson, M. L., 1964. Análisis químico de suelos (Traducido por J. Beltrán). Ediciones Omega, S. A. Barcelona, España. 662 p. Jorgensen, P. & S. León-Yánez. 1999. Catalogue of the vascular plants of Ecuador. Missouri Botanical Garden. San Luis. Lægaard, S. 1992. Influence of fire in the grass paramo vegetation of Ecuador. En: Balslev, H. y J. Luteyn (Eds.). Paramo: an Andean ecosystem under human influence. Academic Press. Londres. Krankina ON, Harmon ME, Winjum K (1996) Carbon storage and sequestration in Russian Forest Sector. Ambio 25: 284-288. Kyrklund, B. 1990. Cómo pueden contribuir los bosques y las industrias forestales a reducir el exceso de anhídrido carbónico en la atmósfera. Unasylva 163(41):12-14. Lazore, A., Gilabert, P. 2007. Estimación del Carbono Capturado en las Plantaciones de Pino Radiata y Eucaliptos Relacionadas con el DL-701. 51 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 León-Yánez, S. 2000. La flora de los páramos ecuatorianos. En: La biodiversidad de los páramos. Serie Páramo 7: 5-21. GTP/AbyaYala. Quito. Loguercio G.A. and Defossé G., 2001. Ecuaciones de biomasa aérea, factores de expansión y reducción de la lenga Nothofagus pumilio (OPEP et Endl.) Krasser en el SO de Chubut, Argentina. Simposio Internacional sobre Medición y Monitoreo de la Captura de Carbono en Ecosistemas Forestales. Valdivia, Chile 18–20 de Octubre de 2001. Luteyn, J. 1999. Páramos, a checklist of plant diversity, geographical distribution, and botanical literature. New York Botanical Garden Press. Nueva York. Martínez E., Fuentes J., Acevedo E., H. 2008. Carbono Orgánico y Propiedades del suelo. Universidad de Chile. Facultad de Ciencias Agronomicas y Forestales. Plant Nutr. 8 (1) (68-96). Mascha Jacob, Claudia Bade, Hector Calvete, Sebastian Dittrich, ChristophLeuschner, and Markus Hauck 2013. Significance of Over-Mature and Decaying Trees for Carbon Stocks in a Central European Natural Spruce Forest. Ecosystems 16: 336–346. Masera, O. 1995. Carbon mitigation scenarios for Mexican forests: methodological considerations and results. Interciencia 20(6): 388-395. Medina, G .y D. Ortiz.2001. Políticas nacionales y plan de acción para la conservación y manejo del ecosistema páramo en el Ecuador. En: Mena V.,G.Medina y R.Hofstede (Eds.).2001. Los Páramos del Ecuador.Particularidades, Problemas y Perspectivas. Abya Yala/Proyecto Páramo.Quito. Medina, G. 2000. Los páramos: opciones para el futuro. Terra Incognita 2(9): 32-33. Medina, G. y P. Mena. 2001. En: P. Mena, G. Medina y R. Hofstede (Eds.). Los Páramos en el Ecuador. Particularidades, problemas y perspectivas. Abya Yala/Proyecto Páramo. Quito. Medina, G., J. Recharte, E. Suárez y F. Bernal. 1997. Perspectivas para la conservación de los páramos en el Ecuador. Informe final de proyecto de EcoCiencia y el Instituto de Montaña a la Embajada de los Países Bajos. Quito. Mena. P. Y H. Balslev. 1986. Comparación entre la vegetación de los páramos y el cinturón afroalpino. Reports from the Botanical Institute, University of Aarhus 12:1-54. Mena-Vásconez, P. 2001. La Biodiversidad de los páramos en el Ecuador. En: P. Mena Vásconez, G. Medina y R. Hofstede (Eds.). Los Páramos del Ecuador. Particularidades, problemas y perspectivas. Proyecto Páramo/Abya-Yala. Quito. Mittermeier, R., P. Robles y C. Göttsch-Mittermeier. 1997. Megadiversidad. Los países biológicamente más ricos del mundo. CEMEX S.A. y Agrupación Sierra Madre. México. Návar, J. 2009. Allometric equat0.ions for tree species and carbon stocks for forests of northwestern Mexico. Forest Ecology and Management 257: 427-434. doi:10.1016/j.foreco.2008.09.028 Nelson DW, Sommers LE (1996) Total carbon, organic carbon, and organic matter. In: Sparks DL (ed) Methods of soil analysis. Part 3. Chemical methods; SSSA Book Series No. 5. Soil Science Society ofAmerica,Madison,Wisconsin, pp 961–1010 52 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Panel Intergubernamental de Expertos sobre Cambio Climático – IPCC 2005. La captación y el almacenamiento de dióxido de carbono (en línea). Suiza. Disponible en: http://arch.rivm.nl/env/int/ipcc/pages_media/SRCCS-final/IPCC%20Sp.pdf Panel Intergubernamental de Expertos sobre Cambio Climático – IPCC - 2006. Directrices del IPCC de 2006 para los inventarios nacionales de gases de efecto invernadero. Volumen 4: Agricultura, Silvicultura y otros usos de la tierra. Publicado por IGES, Japon. Pauli, H., Gottfried, M., and Grabherr, G. 2003. Effects of climate change on the alpine and nival vegetation of the Alps. J. M. E. Journal of Mountain Ecology 7: 9-12. Prieto F. y Paucar G., 2011. Diversidad de flora y fauna en los páramos y bosques montanos del sur oeste del Parque Nacional Sangay (Provincias de Chimborazo y Cañar). Protocolo de Kyoto de la convención marco de las naciones unidas sobre el cambio climático. 1997. Unfccc.int/resource/docs/convkp/kpspan2007. Quantin (Q.), Zebrowski (C.). 1997. Caractérisation et formation de la cangahua en Equateur. IN : Suelos Volcánicos endurecidos, pp.29-47. Memorias del III Simposio Internacional sobre Suelos volcánicos endurecidos, Quito, Equateur, decembre 96. Editions Orstom Quito - CEE - UCE Quito - UC FCA Quito. Ravindranath N. H. y Oswald M. 2008. Carbon Inventory Methods: Handbook for Greenhouse Gas Inventory, Carbon Mitigation and Roudwood Production Projects. Springer. Ruiz, A. 2002. Fijación y almacenamiento de carbono en sistemas silvopastoriles y competividad económica en Matiguás, Nicaragua. Turrialba, CR, CATIE. 111 p. Ruiz-Peinado R. Bravo-Oviedo A., Lopez-Senespleda E., Montero G., Rio M. 2013. Do thinnings influence biomass and soil carbon stocks in Mediterranean maritime pinewoods. Eur J Forest Res 132:253–262. Sánchez-Hernández, R., Ramos-Reyes, R., Geissen, V., Mendoza-Palacios, J., De la Cruz-Lázaro, E., Salcedo-Pérez, E., Palma-López D. Contenido de carbono en suelos con diferentes usos agropecuarios en el trópico Mexicano. Terra Latinoamericana, vol. 29, núm. 2, abril-junio, 2011, pp. 211-219. Schlesinger, W. H., 2000. Carbon sequestration in soils: some cautions amidst optimism. Agriculture, Ecosystems and Environment, 82, 121–127. Sklenář, P., J.L. Luteyn, C. Ulloa, P.M. Jørgensen y M.O. Dillon. 2005. Flora Genérica de los Páramos. Guía Ilustrada de las Plantas Vasculares. Memoirs of the New York Botanical Garden 92: 3-499. Swift, R. S., 2001. Sequestration of carbon by soil. Soil Sci., 166, 858-871. Torres-Rivera, J., Espinoza-Domínguez, W., LaskmiReddiar-Krishnamurthy, Vázquez- Alarcón, A,. Secuestro de carbono en potreros arbolados, potreros sin árboles y bosque caducifolio de Huatusco, Veracruz Tropical and Subtropical Agroecosystems, vol. 13, núm. 3, 2011, pp. 543-549, Universidad Autónoma de Yucatán México. Vesterdal L., et al. 2007. Carbon sequestration in soil and b iomass following afforestation: experiences from oak and norway spruce chronosequences in Denmark, Sweden and the Netherlands. G.W. Heil et al. (eds.). Environmental Effects of Afforestation in North-Western Europe, 19–51. Springer. 53 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 EFECTO DE LA FRAGMENTACIÓN DEL BOSQUE TROPICAL SECO SOBRE LA VIABILIDAD DE PROGENIES DEL ÁRBOL DIOICO Brosimum alicastrum (Moraceae) Jesús Mª Bastida1,2, Fernando L. Rosas3 & Mauricio Quesada2 1Escuela Superior Politécnica del Chimborazo, Facultad de Recursos Naturales, Riobamba (Chimborazo, Ecuador). Investigador Prometeo-SENESCYT. 2Centro de Investigaciones en Ecosistemas, Universidad Nacional Autónoma de México, Morelia (Michoacán, México). 3Centro de Investigaciones Biológicas, Universidad Autónoma del Estado de Hidalgo, Pachuca (Hidalgo, México) [email protected] Introducción La fragmentación y pérdida de hábitats naturales, junto con el cambio climático global, son los principales motores de la actual crisis de biodiversidad (Sala et al. 2000). Particularmente alarmantes han sido las elevadas tasas de pérdida de hábitats naturales ocurridas en numerosos paises de Suramérica, Africa occidental o del Sureste asíatico durante las últimas décadas fruto de la expansión de la frontera agrícola, infraestructuras, industrialización y complejos turísticos (FAO 2010, Lambin y Meyfroidt 2011). En primera instancia, el efecto más evidente de la fragmentación y consecuente pérdida de hábitats es la creación de fragmentos de hábitat natural inmersos en una matriz, en mayor o menor medida, antropizada y “hostil” (McInyre y Hobbs 1999, Ewers y Didham 2006). Como consecuencia directa de la fragmentación se reducen los tamaños poblacionales, aumenta el aislamiento, se reduce la conectividad e incrementan los efectos de borde (McGarigal y Cushman 2002; Fahrig 2003). Estos cambios a nivel de paisaje tienen profundas implicaciones en la estructura y composición de especies entre y dentro de los fragmentos, condiciona la abundacia y diversidad de interacciones mutualistas y/o antagonistas (i.e. polinización, dispersión, predación o herbivoría) (Aizen y Feinsinger 1994; Aguilar et al. 2006; Kolb 2008); a la vez que modifica variables físicas (i. e. temperatura del aire y suelo, pH, humedad ambiental, humedad edáfica, disponibilidad hídrica, flujos de radiación) responsables del deterioro de microclimático dentro de fragmentos (Didham 1998; Mix et al. 2006), así como la reducción del nicho de regeneración de múltiples especies (Benítez-Malvido 1998). La fragmentación del hábitat en términos generales tiene un fuerte efecto negativo sobre el éxito reproductivo y diversidad genética de numerosas especies de plantas (Aguilar et al. 2006, 2008). Además, es sabido que las plantas de ambientes fragmentados producen en promedio menor cantidad de propágulos (i.e. menor cuajado de frutos y producción de semillas por fruto) que los procedentes de ambientes continuos (Aguilar et al. 2006). A pesar de esto, pocos estudios han valorado el efecto de la fragmentación sobre el desempeño de las progenies, p. ej. peso de la semilla, éxito de germinación o vigor (Rocha y Aguilar 1998; Ashworth y Martí 2010; Aguilar et al. 2012). Siendo la germinación y establecimiento de nuevas progenies estadíos críticos para la persistencia y regeneración a largo plazo de cualquier población de plantas (Kitajima y Fenner 2000). Por tanto, la evaluación de la variabilidad y viabilidad de las progenies de ambientes fragmentados constituye un 54 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 paso esencial para contar con una idea más completa de las consecuencias de la fragmentación sobre la viabilidad de poblaciones de plantas en generaciones futuras. Recientemente, Aguilar et al. (2008) encontraron que los árboles adultos en ambientes fragmentados mostraron tasas de exo-cruzamiento menores que se relacionaron con mayores índices de endogamía en las progenies estudiadas. Esto sugiere que ya sea por limitación de polen, aumento de la tasa de autogamia o apareamineto entre individuos emparentados se están produciendo cambios en los patrones de apareamiento. Tales cambios, en última instancia puede comprometer el desempeño temprano de las progenies (Eckert et al. 2009). Particularmente, rasgos de historia de vida (e. g. peso de la semilla, germinación, tiempo de emergencia, nivel de ploidía, tasas de crecimiento, longevidad foliar), por estar estrechamente relacionados con el desempeño, pueden pueden verse profundamente afectados a causa de depresión por endogamía (Young et al. 1996, Eckert et al. 2009). Por lo tanto, mientras la detección de la erosión de la variabilidad genética producto de la fragmentación puede tardar varias generaciones, en especial entre especies de larga vida, los efectos de la endogamía en progenies (i. e. depresión por endogamía) se pueden sentir en periodos de tiempo considerablemente más cortos y sobre diferentes componentes del desempeño de las progenies (Young et al. 1996; Leimu et al. 2006). Adicionamente, otro factor a tener en cuenta, dada su ubicuidad en el reino vegetal, son los efectos marternos sobre el desempeño de las progenies. La calidad y cantidad de semillas producidas por una árbol pueden verse profundamente afectados por las cambios en las condiones microclimáticas de su entorno inmediato, tanto abióticas (alta intensidad lumínica, baja humedad relativa, mayor desecación) como bióticas (densidad de conespecíficos, disrupción de interacciones mutualistas y antagosnistas), ya que éstas pueden limitar la provisión de recursos destinados al desarrollo y maduración de semillas (Stephenson 1992). Dada la compleja naturaleza de los efectos y procesos ligados a la fragmentación, así como la variabilidad de las respuestas de cada especie (p. ej. tiempo en fragmentación, configuranción especial, sistema de apareamiento, rasgos de vida). Cada vez se hace más apremiante tener una idea más completa del efecto de la fragmentación sobre la viabilidad y persistencia a largo plazo de poblaciones de plantas, para lo cual es necesario evaluar el impacto de la fragmentación sobre la calidad y cantidad de propágulos (Ashworth y Martí 2010). En el presente estudio se evalua el efecto de la fragmentación del bosque tropical seco sobre la viabilidad y desempeño de progenies del árbol dioico Brosimum alicastrum (Moraceae). Basado en los estudios que han demostrado el aumento de los coeficientes de endogamía en progenies procedentes de ambientes fragmentados, así como el deterioro del medio abiótico en los ambientes fragmentados; predecimos que las progenies procedentes de bosque continuo presentaran una mayor viabilidad y desempeño que las progenies procedentes de fragmentos de vegetación 2ª y zonas antropizadas. Material y Métodos Sistema de estudio (Brosimum alicastrum) Brosimum alicastrum (Moraceae) es una especie perennifolia, de lento crecimiento (alcanza la reproducción aprox. A los 20 años) y larga vida (aprox. 150 años) que puede alcanzar hasta los 45 m. de altura. Su distribución va desde México hasta el norte de Perú y las islas del Caribe, habitando tanto selvas secas como húmedas. Respecto a su expresión sexual Br. alicastrum puede presentar poblaciones dioicos (generalmente en zonas de bosque tropical seco) mientras que en lugares más húmedos (como puede ser en Los Tuxlas, Veracruz o en Calakmul, Campeche) las poblaciones son eminentemente monoicos. Estudios sobre la 55 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 polinización de Br. alicastrum sugieren que es polinizado tanto por viento (Berg 2001) como por insectos (Croat 1978, Berg 1990). Generalmente, fructifica y es dispersado en épocas secas, lo que unido a las abundantes cosechas que produce, es un árbol que atrae y mantiene a un sinfín de especies de aves y mamíferos. Entre las especies que dispersan sus frutos se encuentran una gran variedad de mamíferos y aves (chachalacas, monos, murciélagos entre otros), así como es dispersado de forma secundaria por los roedores (Croat 1978). Es una especie con buena regeneración y que puede ser facilitada por ciertos arbustos en ambientes perturbados o pastizales (Sánchez-Velázquez et al. 2004). De forma adicional, es un árbol frecuentemente usado como cercas vivas y potreros donde puede formar bosquetes prácticamente monoespecíficos. Por estas características, Br. alicastrum puede considerarse una especie clave para el mantenimiento de la conectividad y funcionalidad a nivel de paisaje y ecosistema. Área de estudio Las poblaciones de árboles evaluados en este estudio se encuentran dentro y alrededor de la Reserva de la Biosfera Chamela-Cuixmala (ca. 19 ° 30.0 N, 105 ° 0.30 W), que abarca 13.200 hectáreas de bosque tropical seco conservado. La reserva se ubica en la costa del Pacífico en el estado de Jalisco, México; y se caracteriza por una marcada estación seca de noviembre a mayo, con un 80 % de la precipitación que cae entre julio y octubre (707 mm/año). El tipo de vegetación es una densa selva tropical caducifolia, compuesta por dos principales hábitats: bosque tropical seco y la vegetación de ribera que se encuentra a lo largo de los grandes ríos y arroyos estacionales (Bullock, 1986; Lott, 1993). Los bosques tropicales secos son uno de los ecosistemas más amenazados del mundo. Los estudios indican que han sido las zonas preferidas para el desarrollo de la agricultura (debido a la mayor fertilidad de sus suelos respecto a los bosques tropicales húmedos) y asentamientos humanos (complejos turísticos) en el neotrópico (Murphy y Lugo, 1986) y están entre los más utilizados, perturbados, y menos conservados de los grandes ecosistemas tropicales (Quesada y Stoner, 2004). Una reciente evaluación de la cubierta vegetal y patrones de fragmentación en el área de estudio (Sánchez-Azofeifa et al. 2009, Portillo-Quintero y Sánchez-Azofeifa, 2010) indica que la cobertura de la tierra dentro de la reserva incluye: el 92,1 % de bosque seco, el 2% de los bosques de ribera, y el 3,7 % de la superficie transformada (campos agrícolas o pastos). Por el contrario, las mismas categorías de cobertura de la tierra fueron, respectivamente, 79,85 %, 1,65 % y 16,55 % en un área de 10 kilómetros que rodea la reserva, donde se encuentran rodales y árboles aislados de Br. alicastrum en parches de tamaño medio que van desde 8.5 hasta 19.17 ha (Tabla 1). Muestreo de poblaciones de Brosimum alicastrum Entre diciembre de 2010 y Octubre de 2011 se realizaron muestreos destinados a la localización del mayor número de poblaciones dentro del área de estudio. Se georeferenciaron entre el 70%-100% de ejemplares en 10 localidades de Br. alicastrum a lo largo de la costa de Jalisco (México), que tomando como centro la Reserva Biológica de Chamela-Cuixmala progresivamente se alejan cumbriendo un amplio mosaico de ambientes y estados de fragmentación. En cada población se realizaron muestreos extensivos perimetrales destinados a delimitar y confirmar la extensión de cada población a fin evitar dejar fuera posibles árboles reproductores. A cada árbol georreferenciado se determinó el sexo mediante observación directa de flores y el diámetro a la altura del pecho. Para cada localidad se generaron archivos gpx para su posterior manejo y análisis con el software QGIS 1.7 (ref.). La base de datos georreferenciados así obtenida permite caracterizar cada localidad a distintas escalas espaciales, así como evaluar la fragmentación y la matriz ambiental en la que se hayan embebidas. Cada población se delimitó a partir de la identificación de agrupaciones discretas de 56 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 individuos en el paisaje, i. e. cada deme o subpoblación, que vendría definida por cada unidad de agrupación de individuos que apareciesen discretas tras la aplicación de un buffer de agregación de 50 m. a cada individuo georreferenciado. Tras esto, cada población fue definida como el polígono mínimo convexo que agrupa a todos las agregaciones discretas de individuos de un área (tras uso de buffer 50m). Se utilizaron mapas de fragmentación en la costa de Jalisco 1973-1992; mapas de uso de suelo en México 2005 y 2010; (CONABIO) para asignar a cada árbol madre una estado de fragmentación. Experimento de viabilidad y desempeño de progenies de Brosimum alicastrum Con la finalidad de explorar el efecto de la fragmentación sobre las progenies del Br. alicastrum se seleccionaron 45 madres (Tabla 1). Las madres seleccionadas representan los distintos estados de perturbación del hábitat en la región de Chamela (bosque continuo-conservado, fragmentos de vegetación secundaria y zonas antrópizadas (cultivos, potreros, zonas urbanas)). El desempeño de las progenies, medido como el éxito de germinación, crecimiento y supervivencia, fue evaluado a través de un experimento en invernadero de siembra de progenies aleatorizado donde se mantuvo la identidad de cada árbol madre y de las semillas. En junio de 2011, un total de 875 semillas de 45 madres (15-20 semillas por madre) procedentes de condiciones (Tabla 1) distintas fueron pesadas y sembradas individualmente. Cada bolsa de siembra se rellenó con una mezcla de turba, agrolita y arena (2:1:4) y cada semilla se sembró a una profundidad de 0.5 cm. Las siembras fueron regadas dos veces en semana. La germinación y mortalidad de plántulas se monitoreó dos veces en semana hasta el final del experimento. Para cada plántula se midió el éxito de germinación, número de días en germinar (tiempo de emergencia) como un inverso de la velocidad de germinación y supervivencia despues de un año. Tambien se midió el número de hojas producidas transcurridos un mes de la germinación. Al final del experimento (Junio de 2012) cada plántula fue cosechada y se les midió el número de hojas totales, altura, longitud de la raiz, diametro basal, área foliar media, area foliar total y tasa de producción foliar. Finalmente, cada plántula fue secada a 60º por 72 horas y su biomasa total medida. Análisis estadísticos El desempeño de las progenies se evaluó mediante MIXED y GLIMMIX procedures en SAS 9.3. Debido a la proximidad de las localidades estudiadas y a la probable conexión vía flujo de polen o en décadas pasadas, el presente diseño incluye el efecto “Estado de fragmentación” (Bosque continuo, fragmentos de bosque y zonas antropizadas) como factor fijo y el efecto “Madre” como factor aleatorio anidado dentro de “Condición del hábitat” (Tabla 2). Para cada variable se usaron distribución binomial (Germinación y Supervivencia), poisson (Días en emerger, Nº de hojas tras un mes, Nº de hojas totales) o normal (biomasa, altura de la plantula, área foliar, longitud de la raíz). El efecto del peso de la semilla (Sm), así como el tiempo de emergencia (Em), en las variables medidas fue explorado para su inclusión en el modelo como covariables. Para los modelos MIXED la covariable fue incluida de acuerdo al Criterio de Información de Akaike (AIC) (i.e. AIC con covariable < AIC sin covariable), de modo que la covariable fue incorporada si mejoraba el modelo. Para GLIMMIX, si se detectaron efectos significativos entre las covariables y las variables respuestas (vía regresión logística), éstas fueron incluidas en el modelo. Resultados 57 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 En general, tanto el peso de la semilla como el tiempo de emergencia, mejoraron los modelos y por tanto fueron incluidas como covariables (excepto para la variable Nº de hojas totales). El peso de la semilla y el tiempo de emergencia afectaron a la viabilidad (excepto para la supervivencia) y desempeño de las progenies (excepto Nº de hojas totales) (ver Tabla 2). En términos de viabilidad, i. e. éxito de germinación, tiempo de emergencia y supervivencia, las progenies no mostraron diferencias estadísticamente significativas respecto al estado de fragmentación (Tabla 2). En cambio, el desempeño de las progenies sí mostró diferencias relacionadas con el estado de fragmentación (Tabla 2). Concretamente, las progenies procedentes de madres ubicadas dentro bosque conservado continuo mostraron mayor número de hojas totales, area foliar total y biomasa que las procedentes de fragmentos de vegetación 2ª y zonas antropizadas (Figura 1-3). Por el contrario, las progenies procedentes de fragmentos de vegetación 2ª y zonas antropizadas mostraron mayores diámetros basales; y en caso de progenies de madres de zonas antropizadas también mayor longitud de la raíz (Figura 4-5). Además, se detectó un importante y generalizado efecto materno en las variables exploradas atribuible a diferencias en germinación, tiempo de emergencia y desempeño de las progenies de las madres dentro de un mismo estado de fragmentación (Tabla 2). En resumen, las semillas procedentes de bosque conservado continuo produjeron plántulas más grandes y con mayor area foliar, i.e. relacionado con la capacidad competitiva por recursos que las de fragmentos de vegetación 2ª y zonas antropizadas. Además llama la atención, la respuesta fenotípica de las plántulas de fragementos de vegetación 2ª y zonas antropizadas, al desarrollar diámetros basales y raices mayores, i. e. relacionado con estrategias de tolerancia a estres ambiental. Discusión La reducción del número de posibles donadores de polen, de la diversidad y abundancia de polinizadores o el aumento de endogamía y paternidad correlacionada tienen un efecto negativo sobre la cantidad y calidad de semillas que produce un determinado árbol (Aguilar et al. 2006; Quesada et al. 2004; Eckert et al. 2009). En consecuencia, se predice que la fragmentación tendrá un efecto negativo sobre la viabilidad y desempeño de las progenies. Sin embargo, los estudios realizados al respecto en otras especies de bosque tropical seco han mostrado distinto apoyo esta hipótesis (Rocha y Aguilar 1998; Ashworth y Martí 2010; Aguilar et al. 2012, Cascanta et al. 2002). En terminos generales nuestros resultados siguen la linea de estudios anteriores y corroboran parcialmente la hipótesis inicial. Así, no se detectaron efectos negativos significativos de la fragmentación sobre la germinación, tiempo de emergencia y supervivencia. Por ejemplo, Ashworth y Martí (2010) y Aguilar et al. (2012) tampoco detectaron efectos negativos de la fragmentación en la germinación de cuatro de cinco especies del Chaco argentino, de las cuales 4 son especies de árboles. Sin embargo, Aguilar et al. (2012) si detectó diferencias en la supervivencia de las plántulas procedentes de distintos estados de fragmentación. Por el contrario, nuestros resultados indican que el desempeño de las progenies sí es afectado por la fragmentación. Las progenies procedentes de bosque continuo presentaron mayor tamaño y area foliar que el resto. Esto sugiere que las plántulas de esta procedencia puede presentar ciertas ventajas competitivas respecto a la captura y ocupación de recursos (p.ej. luz, espacio, agua). Similares resultados fueron encontrados para Enterolobium ciclocarpum (Rocha y Aguilar 1998) y Prosopis caldenia (Aguilar et al. 2012), plántulas de ambientes continuos fueron mayores (biomasa, desempeño acumulado, RGR). Entre las principales causas relacionadas con un pobre desempeño de las progenies provenientes de ambientes fragmentados se encuentra el incremento en los niveles de endogamía (Aguilar et al. 2008; Eckert et al. 58 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 2009). Alternativamente, se ha demostrado que en ciertas condiciones de fragmentación árboles anemófilos han incrementado la extensión del flujo de polen evitando así los efectos negativos como la depresión por endogamía (Wang et al. 2010). En otros casos es la propía distribución espacial de los árboles aislados y fragmentos los que mantienen la conectividad vía polen (Lander et al. 2010). También llama la atención que las plántulas de ambientes fragmentados mostraron mayor diámetro basal y longitud de la raíz, lo cual puede resultar ventajoso bajo las condiciones estresantes propias de los ambientes fragmentados. En términos de resistencia o tolerancia al estrés ambiental cambios en el reparto de biomasa entre la parte aéreo y subterranéo, pueden implicar reflejar una mayor sistema radical que relaciona con la capacidad de obtener recursos hídricos. López-Mata (1987) también encontró variación en el peso seco de las raices de plántulas Br. alicastrum de distintas procedencias, lo cual se relacionó con la economía hídrica. Sin embargo, el valor adaptativo de estos rasgos en ambientes fragmentados o pastizales, por ejemplo contribuyendo en el mantenimento de la biomasa, queda más hallá de los objetivos de este estudio. En resumen para el caso de Br. alicastrum, proponemos que, ya sea por el corto número de generaciones en fragmentación, por un posible flujo de polen extensivo a escala de paisaje, así como por su conservación en potreros y zonas agrícolas los efectos negativos de la fragmentación sobre la viabilidad y desempeño de las progenies pueden haber sido parcialmente mitigados. El estudio de la variación en desempeño en relación a la fragmentación de progenies provenientes de madres en distinto estado de fragmentación aporta información relevante sobre la persistencia a largo plazo de la población que puede ser de utilidad para planes de manejo, programas de reforestación o restauración in situ. Bibliografía Aguilar, R., Ashworth, L., Galetto, L. y Aizen, M. A. (2006) Determinants of plant reproductive susceptibility to habitat fragmentation: Review and synthesis through a meta-analysis. Ecology Letters 9, 968–980. Aguilar, R., Quesada, M., Ashworth, L., Herrerias-Diego, Y. y Lobo, J. (2008) Genetic consequences of habitat fragmentation in plant popula- tions: Susceptible signals in plant traits and methodological approaches. Molecular Ecology 17, 5177–5188. Aizen, M. A. y Feinsinger, P. (1994) Habitat fragmentation, native insect pollinators, and feral honey bees in Argentine Chaco Serrano. Ecological Applications 4, 378–392. Benítez-Malvido, J. (1998) Impact of forest fragmentation on seedling abundance in a tropical rain forest. Conservation Biology 12, 380–389. Berg CC (1990) Differentiation of flowers and inflorescences of Urticales in relation to their protection against breeding insects and to pollination. Sommerfeltia, 11, 13–24. Berg CC (2001) Moreae, Artocarpeae, and Dorstenia (Moraceae) with Introductions to the Family and Ficus and with Addi- tions and Corrections to Flora Neotropica Monograph 7. Bullock, S.H., (1986) Climate of Chamela, Jalisco, and trends in the south coastal region of Mexico. Archives for Meteorology, Geophysics, and Bioclimatology 36, 297–316. Croat T (1978) Flora of Barro Colorado Island. Stanford Univer- sity Press, Stanford, California. 59 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Didham, R. K. (1998) Altered leaf-litter decomposition rates in tropical forest fragments. Oecologia 116: 397– 406. Eckert, C.G., Kalisz, S., Geber, M.A., Sargent, R., Elle, E., Cheptou, P.O., Goodwillie, C., Johnston, M.O., Kelly, J.K., Moeller, D.A., Porcher, E., Ree, R.H., Vallejo-Marín, M., Winn, A.A., (2009) Plant mating systems in a changing world. Trends in Ecology and Evolution 25, 35–43. Ewers, R. M. y Didham, R. K. (2006) Confounding factors in the detection of species responses to habitat fragmentation. Biological Review 81: 117–142. Fahrig, L. (2003) Effects of habitat fragmentation on biodiversity. Annual Review of Ecology and Systematics 34, 487–515. Kitajima, K. y Fenner, M. (2000) Ecology of seedling regeneration. In: Fenner M, editor. Seeds: the ecology of regeneration in plant communities. 2nd edn. Wallingford, UK: CAB International; 2000, 331-359. Kolb, A. (2008) Habitat fragmentation reduces plant fitness by disturbing pollination and modifying response to herbivory. Biological Conservation 41, 2540–2549. Lander, T. A., Boshier, D. H. y Harris, S. (2010) Fragmented but not isolated: Contribution of single trees, small patches and long-distance pollen flow to genetic connectivity for Gomortega keule, an endangered Chilean tree. Biological Conservation 143, 2583-2590. Lambin, E.F. y Meyfroidt, P. (2011) Global land use change, economic globalization, and the looming land scarcity. Proc. Natl. Acad. Sci. USA 108, 3465–3472. Leimu, R., P. Mutikainen, J. Koricheva & M. Fischer. 2006. How general are positive relationships between plant population size, fitness and genetic variation? Journal of Ecology 94, 942–952. López-Mata, L. (1987) Genecological Differentiation in Provenances of Brosimum aUcastrum - a Tree of Moist Tropical Forests. Forest Ecology and Management 21, 197-208. Lott, E.J., 1993. Annotated checklist of the vascular flora of the Chamela bay region Jalisco, Mexico. Occasional Papers of the California Academy of Sciences 148, 1– 60. Mcgarigal, K. y Cushman, S. A. (2002) Comparative evaluation of experimental approaches to the study of habitat fragmentation effects. Ecological Applications 12, 335–345. McIntyre, S. y Hobbs, R. (1999) A framework for conceptualizing human effects on landscapes and its relevance to management and research models. Conservation Biology 13(6): 1282‐1292. Mix, C., Pico, F. X., Van Groenendael, J. M. y Ouborg, N. J. (2006) Inbreeding and soil conditions affect dispersal and components of performance of two plant species in fragmented landscapes. Basic and Applied Ecology 7, 59–69. Sánchez-Azofeifa, G.A., Quesada, M., Cuevas-Reyes, P., Castillo, A., Sánchez- 60 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Montoya, G. (2009) Land cover and conservation in the area of influence of the Chamela-Cuixmala Biosphere Reserve, Mexico. Forest Ecology and Management 258, 907–912. Murphy, P.G., Lugo, A.E., 1986. Ecology of tropical dry forest. Annual Review of Ecology and Systematics 17, 67–88. Portillo-Quintero y Sánchez-Azofeifa (2010). Quesada, M., Stoner, K.E., Lobo, J.A., Herrerías-Diego, Y., Palacios-Guevara, C., Mungía-Rosas, M.A. et al. (2004) Effects of forest fragmentation on pollinator activity and consequences for plant reproductive success and mating patterns in bat pollinated bombacaceous trees. Biotropica, 36, 131–138. Quesada, M. y Stoner, K.E. (2004) Threats to the conservation of the tropical dry forest in Costa Rica. In: Frankie, G.W., Mata, A., Vinson, S.B. (Eds.), Biodiversity Conservation in Costa Rica: Learning the Lessons in a Seasonal Dry Forest. University of California Press, Berkeley, California, pp. 266–280. Sánchez-Velázquez, L. R., Quintero-Gradilla, S., Aragón-Cruz, F. y Pineda-López, Ma R. (2004) Nurses for Brosimum alicastrum reintroduction in secondary tropical dry forest. Forest Ecology and Management 198 (1), 401-404. Sala, O.E., Chapin III, F.S., Armesto, J.J., Berlow, E., Bloomfield, J., Dirzo, R., HuberSanwald, E., Huenneke, L.F., Jackson, R.B., Kinzig, A., Leemans, R., Lodge, D.M., Mooney, H.A., Oesterheld, M., Poff, N.L., Sykes, M.T., Walker, B.H., Walker, M., Wall, D.H. (2000) Global biodiversity scenarios for the year 2100. Science 287, 1770–1774. Stephenson, A. G. (1992) The regulation of maternal investment in plants. In C. Marshal and J. Grace (Eds.). Fruit and seed production: Aspects of development, environmental physiology and ecology, pp. 151–171. Cambridge University Press, Cambridge, London. Wang, J., Kang, M., Gao, P. y Huang, H. (2010) Contemporary pollen flow and mating patterns of a subtropical canopy tree Eurycorymbus cavaleriei in a fragmented agricultural landscape. Forest Ecology and Management 260, 2180-2188. Young, A., Boyle, T. & Brown, T. (1996) The population genetic consequences of habitat fragmentation for plants. Trends Ecol. Evol. 11: 413–418. 61 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Tabla 1. Localidades de Brosimum alicastrum muestreadas a lo largo de la costa del estado de Jalisco. Localidades Nº indivs. (madres Razón sexual focales) Dbh (cm) Chamela 22(8) 1.625 228 Careyes 204(14) 0.81 Caiman 161(10) Zapata medio Densidad (indiv/ha) Área (ha) % Área Nº aislados ocupada 500m) 0.132 166.4 16.95 0 229.1 1.236 165 19.8 0 1.0 205.1 0.3356 479.6 5.75 2 14(5) 0.33 399.5 0.2208 63.40 15.75 1 A.Seco 50(5) 1.888 226.2 0.5534 90.35 10.14 0 El Tecuan 109(4) 1.25 190 0.3783 288.1 7.11 2 La Mesa 22 1.333 240 2.5821 8.52 27.5 0 Aguacatillo 28 - 247.8 1.4605 19.17 45.4 2 Mazanilla 30 - 360.8 1.7729 16.92 29.7 1 62 árboles (> PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Tenacatita 7 - 345 0.4208 16.63 25.2 1 Tabla 2. Resultados de los efectos del estado de fragmentación (Bosque continuo/ Fragmento de vegetación 2ª/Áreas antropizadas) sobre la germinación, supervivencia y crecimiento de porgenies de Brosimum aliscastrum. 63 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Covariables en el modelo Estado de fragmentación Madre(Estado de fragmentación) d.f. F P d.f. F P Germinación 2, 826 0.0 0.99 42, 826 2.48 <.0001 Sm Días en emerger 2, 826 0.05 0.95 42, 826 3.78 <.0001 Sm Surpervivencia después de 1 año 2, 646 0.00 1.00 42, 646 0.25 1.00 Sm Nº hojas tras de un mes 2, 613 1.46 0.23 42, 613 3.34 <.0001 Sm Nº hojas tot. 2, 613 7.23 0.0008 42, 613 18.21 <.0001 Area Foliar tot. 2, 613 10.99 <.0001 42, 613 26.26 <.0001 Sm, Et Area Foliar media 2. 613 0.75 0.47 42, 613 6.73 <.0001 Sm, Et Longitud de la raiz 2, 613 13.92 <.0001 42, 613 6.24 <.0001 Sm, Et Diámetro basal 2, 613 3.38 0.03 42, 613 5.51 <.0001 Sm, Et Altura de la plántula 2, 613 2.43 0.08 42, 613 40.71 <.0001 Sm, Et Biomasa tot. 2, 613 3.91 0.02 42, 613 33.98 <.0001 Sm, Et Medidas de desempeño Germinación y supervivencia Crecimiento 64 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figura 1. Rasgos relacionados con el desempeño con diferencias significativas respecto al estado del fragmentación (bosque continuo, fragmentos de vegetación 2ª y zonas antrópizadas). Las letras indican diferencias estadísticamente significativas. 65 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 EVALUACIÓN ECOLÓGICA RÁPIDA DE LA FAUNA EXISTENTE EN EL BOSQUE TROPICAL SIEMPRE VERDE DEL CANTÓN EL CARMEN, MANABÍ Jimmy J. Cevallos Zambrano Universidad Laica Eloy Alfaro de Manabí [email protected] Problema La tala indiscriminada de los relictos existentes de bosque siempre verde en la costa ecuatoriana avanza en sus etapas finales, al borde de la extinción de miles de especies de fauna existentes en ello, el bosque de la Universidad Laica Eloy Alfaro de Manabí en el Cantón el Carmen mantiene un gran número de especies de fauna muy importantes que no están siendo protegidas en la actualidad, lo que podría lograr su exterminio y con ello muchos impactos negativos. La declaratoria de reserva vegetal o área protegida de esta zona es urgente y por ello se ha emprendido con el presente estudio. Resumen Realizar un Inventario básico de Fauna existente en este bosque identificando las especies y estudiando sus relaciones ecológicas fue el principal objetivo de este estudio. El bosque mantiene una diversidad faunística muy importante demostrado aplicando la metodología de Evaluación Ecológica Rápida, realizando un conteo de individuos, de indicios encontrados y, de indicios sonoros durante un determinado tiempo; para el efecto de esta investigación se consideró una duración de 16 días consecutivos y de manera casi permanente estando en los puntos de conteo durante más de 17 horas al día. Se establecieron 3 puntos de conteo que estuvieron vigilados por este tiempo en el cual se registró todos y cada uno de los individuos que lleguen al lugar; además se establecieron 3 transeptos equidistantes de una dimensión de 200 metros en los cuales se hacían recorridos constantes durante el día y la noche, como complemento a la metodología de estudio se desarrolló en la finca de la granja experimental El Carmen, de la Universidad Laica Eloy Alfaro de Manabí; Extensión el Carmen. Tiene una superficie de más de 31 Has. Es importante mencionar que los sitios de conteo y transeptos se establecieron específicamente en la zona del bosque en estudio. Entre los resultados obtenidos se pueden mencionar que se determinó la presencia de 3 clases taxonómicas que son: mamíferos, aves, y reptiles; dentro de las cuales se identificó 19 órdenes, 34 familias, 42 géneros y 48 especies. Es importante señalar que se registró un número de 216 avistamientos u observación con un promedio de 54 observaciones por día, también se registraron 38 indicios que determinan la presencia de ciertas especies. Objetivos El presente trabajo conlleva como objetivo general la determinación de la biodiversidad faunística del bosque en estudio mediante la realización de un Inventario de fauna del bosque utilizando la metodología de evaluación ecológica rápida a través de puntos de conteo y de manera cuantitativa. Procedimiento La metodología utilizada fue mediante puntos de conteo u observación o también llamada como “Evaluación Ecológica Rápida” que consistía en el establecimiento de 3 puntos de conteo con observadores durante 17 horas al día, por 5 días consecutivos. 66 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Se estableció también 3 transeptos de 200 metros en los cuales se monitorearon y registraron todos los individuos en diferentes horas del día y la noche. De manera complementaria también se realizaron recorridos de observaciones más amplias a través de los predios de la finca. Para el establecimiento de los puntos de conteo y transeptos se procedió previamente a realizar varios recorridos por todo el predio del bosque y de la finca en general, en estos recorridos se determinó el área más representativa para el estudio a realizar. El establecimiento de una parcela permanente de una hectárea, en la misma que se procedió de la siguiente manera: Se realizaron monitoreo en cada uno de los puntos de conteo desde las 6:00H hasta las 22:00h, de manera intercalada entre dos investigadores cada 6 horas. Durante el día y la noche dos investigadores realizaban recorridos constantes a través de los transeptos registrando todos los avistamientos posibles así como indicios o huellas encontradas. El monitoreo en general consistía en la toma de datos sonoros, visuales, indicios, huellas, capturas fotográficas y registros de referencias del guía de campo. Resultados 3 clases taxonómicas 19 órdenes 34 familias 42 géneros 48 especies 216 avistamientos – Individuos 54 observaciones por día 38 indicios de mamíferos mayores. (tigrillo, tejón, venado. Etc.) Tabla 1. Descripción taxonómica básica de los individuos inventariados No Clase Orden Familia Género 25 42 3 2 5 22 Coraciformes Passeriformes Squamata Squamata Xenarthra (Edentata) Accipitriformes Alcedinidae vireonidae Boidae Crotalidae Bradypodidae Accipitridae Alcedo bellii Boa Bothrops Bradipus Buteo 5 4 1 1 1 1 39 Aves Caprimulgiformes Caprimulgidae Caprimulgus 4 C. ruficollis 8 23 15 17 7 6 11 41 9 Chiroptera Apodiformes Piciformes Cuculiformes Rodentia Rodentia Cingulata Passeriformes Didelphimorphia Phyllostomidae Trochilidae Picidae Cuculidae Agoutidae Dasyproctidae Dasypodidae Parulidae Didelphidae Carollia chlorostibon Colaptes Crotophaga Cuniculus Dasyprocta Dasypus Dendroica Didelphis Piciformes Picidae Dryocopus Aves Aves Reptilia Reptilia Mammalia Aves Mammalia Aves Aves Aves Mammalia Mammalia Mammalia Aves Mammalia 14 Aves 67 Ind. Especie 1 25 6 16 1 3 1 6 2 Alcedo atthis bellii pusillus B. Constrictor Bothrops asper B. variegatus Buteo platypterus C. perspicillata. C. Swainsonii C. melanolaimus Crotophaga ani Cuniculus paca D. puntata D. Novemcinctus Dendroica petechia D. marsupialis 3 Dryocopus lineatus N. Vulgar Martin pescador vireo belli Mata Caballo Culebra x, Cascabel Peresoso Gavilan común huevo arrastradochotacabras murcielago pardonegro chupa flor carpintero real común Garrapatero Guanta Guatuzo armadillo vireo amarillo Zorro - zariguella carpintero de copete rojo PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 47 12 40 19 Mammalia Mammalia Aves Aves Carnivora Rodentia Psittaciformes Passeriformes Mustelidae Gliridae Psittacidae Furnariidae Eira Eliomys Forpus Furnarius 32 Aves Passeriformes Icteridae Icterus 4 Icterus cucullatus 33 Aves Passeriformes Icteridae Icterus 7 icterus nigrogularis 16 Aves Charadriiforme Jacanidae Jacana 6 jacana jacana 28 Aves Passeriformes Furnariidae Lepidocolaptes 9 L. leucogaster 21 Aves Accipitriformes Accipitridae Leucopternis 1 L. occidentalis 30 Aves Passeriformes Tyranidae Machetornis 9 M. rixosus 31 Aves 37 Aves 45 Aves Passeriformes coraciiformes Apodiformes Icteridae Momotidae Strigidae Molothrus Momotus Otus 8 Molothrus aeneus 5 M. momota 3 Otus nudipes Squamata Squamata Colubridae. Colubridae. Oxibelis Oxibelis 1 Oxibelis fulgidus 1 Oxibelis aeneus 13 Aves 18 Aves 20 Aves Piciformes Passeriformes Gruiformes Picídae Tyrannidae Rallidae Picus pitangus Porphyrula 6 Picus viridis 7 p. sulphuratus 6 P. martinicase 10 Mammalia Carnívoro Procyonidae Potos 2 Potos flavusus 48 44 24 38 Carnivora Piciformes Apodiformes coraciiformes Procyonidae Ramphastidae Trochilidae Thraupidae Procyon Ramphastos Ramphomicron thraupis 29 Aves Passeriformes Cotingidae Tityra 5 Tityra semifasciata 26 Aves Trogoniformes Trogonidae Trogon 2 Trogon citreolus 27 46 35 36 43 34 Trogoniformes Strigiformes Passeriformes Passeriformes Passeriformes Passeriformes Trogonidae Tytonidae vireonidae vireonidae vireonidae vireonidae Trogon Tyto vireo vireo vireo Vireo 1 Reptilia 4 Reptilia Mammalia Aves Aves Aves Aves Aves Aves Aves Aves Aves 68 1 2 7 14 2 5 14 16 2 1 7 5 4 18 Eira barbara Eliomys quercinus F. xanthopterygius Furnarius rufus Procyon spp. R. sulfuratus R. microrhynchuym thraupis episcopus Trogon elegans Tyto alba V. philadelphicus vireo leucophrys Vireo solitarius Vireo gilvus cabeza de mate zorrillo- rata de monte perico de escoba hornero - oyerita linchero- cacique platanero cacique común gorrion Gallareta de aguajacana cucarachero trepatroncos gavilán negro platanero- atrapa moscas tordo ojirrojo- negro tilingo tordote - pedrote lechuza común CULEBRA DE VEJUCO C. Vejuco Parda carpintero verde amarillo benteveo- sota gavilán gallineta - gallito azul cuzumba - mico de noche zorra patona- mapache - tejón tucán – pilí chupa flor verde Azulejo verraco - puerquito blanco trogón- maría tonta celeste trogón – maría tonta roja lechuza pecho blanco vireo. Chicle verdoso vireo - amarillo verdoso. vireo solitario vireo - chicle marron PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Imagen No. 1.- La Guanta. Cuniculus paca, especie poco abundante y muy consumida por los pobladores de exquisita carne es una de las especies más importantes del bosque. Imagen No. 2.- El Guatuzo. Dasyprocta puntata, especie también poco abundante y muy consumida por los pobladores, excelente carne y representa una fuente de alimento muy saludable. 69 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Imagen No. 3.- Gallareta de agua, Porphyrula martinicase, se la vio varios individuos en las lagunas del bosque. Con presencias en todas las horas del día. Imagen No. 4.- El cacique. Icterus nigrogularis, ave muy importante conocida y famosa por sus silbidos y colores. Abundantes en el bosque. 70 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Imagen No. 4.- El Perico Ligero, Bradipus variegatus, especie en peligro de extinción, pocos individuos en el bosque. Discusión El número de individuo encontrado por unidad de superficie en el bosque es muy importante para tomar la decisión de conservación de esta superficie. Los resultados del presente trabajo han sido muy considerables ambientalmente, el bosque en estudio mantiene una diversidad faunística muy importante. Es muy considerable manifestar que se estima en base a la información proporcionada por los directivos de la Universidad, Extensión El Carmen; que existen una variedad de anfibios dentro del bosque ya que se cuenta con tres lagunas que abarcan una superficie total de más de 6000m2 lo cual mantiene un importante nicho biológico para múltiples especies que no han sido inventariadas en este estudio. Los resultados obtenidos en la presente evaluación están por encima de la media nacional en gran parte de los datos obtenidos lo que hace que sea muy importante para considerar su protección desde ya. En el caso de los indicios encontrados se manifiesta que se deben en su gran mayoría a mamíferos como tejones, tigrillos, venados, cabeza de mate, entre otros lo cual se puede mencionar que estos animales circundan por las noches estas tierras y que pueden o no vivir en este bosque ya que pueden movilizarse grandes extensiones fácilmente, pudiendo venir de bosques vecinos sin embargo esto implica la bondad del bosque en estudio. Dentro de la familia de los reptiles vale destacar que no se hicieron trabajos encaminados directamente a la captura y evaluación de estos ya que se realizó de manera general, por tanto y según criterios de los pobladores se pueden encontrar dentro del bosque una gama de reptiles como son: iguanas, culebra ciega, culebra de bejuco, culebra de mono, culebra sayama, lagartos o tulicios, entre otros. Los mismos que tampoco han sido inventariados con exactitud. Se debe considerar que la presente investigación ha sido una metodología de evaluación rápida, por tanto es fundamental tener en cuenta que no se han considerado modelos ni análisis estadísticos respectivos concernientes a fauna silvestre. 71 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Bibliografía Alcívar A. 2012. Taller sobre procedimientos veterinarios para fauna silvestre. Dirección de ambiente de manabi infomanabi. Disponible en línea. Infomanabi.wordpress.com/category Diario La Hora. 2003. Bosques de Manabí. Biodiversidad de bosque húmedo. (en línea) Portoviejo Ecu. 2003 www.dlh.lahora.com Diario Manabita. 2002. Suplemento Manabí por cantones. El Carmen, (en línea) 2003, www.eldiario.com FAO. 2012. La fauna silvestre www.fao.org/docrep/017/i2498s/i2498s00.htm en un clima cambiante. Disponible en línea. Grupo de estudios de fauna silvestre 2012. Disponible en línea. Https://es-es.facebook.com/faunasilvestreurp Monografías.com. 2013. Evaluación de fauna silvestre altoandina. Disponible en línea. www.monografias.com/...faunasilvestre.../evaluacion-poblacional-fauna. Neill. 2003. Curso-pasantía pre profesional en Botánica y conservación de la biodiversidad. Missouri Botanical Garden, Herbario nacional del Ecuador. Quito- Napo, Ecu. p. 35. Universidad Laica “Eloy Alfaro” de Manabí. 2012. Repositorio.uleam.edu.ec/bitstream. Lugares con menos estudios sobre la fauna silvestre. 72 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 ESTRUCTURA Y COMPOSICIÓN DEL BOSQUE SECO SEMIDESIDUO DE LA RESERVA DE BOSQUE SECO LALO LOOR DEL CANTÓN JAMA. MANABÍ Jimmy J. Cevallos Zambrano Jardin Botánico de Missouri (MO) EE. UU. Universidad Laica Eloy Alfaro de Manabí [email protected] Problema La declaratoria de área protegida del bosque seco semi deciduo Lalo Loor demandó la realización de un estudio técnico para el diagnóstico de la flora existente lo cual se traduce en la necesidad de un inventario florístico y determinación de las dinamicas del bosque y por ende de los indicadores de vegetación existentes como el índice de valor de importancia y la dominancia de la vegetación. El desconocimiento de la composición florística de este bosque era eminente por lo que se hacía cada vez más importante un estudio de esta naturaleza. Resumen El presente trabajo se desarrolló en la Reserva de Bosque Seco Lalo Loor en las siguientes coordenadas: 0º 4’ 48” (S) 80º 09’ 07” (W) en la comunidad Camarones del Norte del Cantón Jama, fue un estudio cuantitativo de la estructura y composición de bosque seco semidesiduo. Identificar taxonómicamente mediante la utilización de claves y estudios de morfología y fisiología a todas y cada una de las especies encontradas, determinar cuál es la especie dominante en el bosque a estudiarse, determinar si existen nuevos géneros o especies vegetales en el mencionado ecosistema y analizar su estructura y composición general fueron los principales objetivos. Estableciéndose dos parcelas permanentes de una hectárea cada una, en las siguientes coordenadas. Parcela # 1: 00º 05’ 07” (S) y 80º 09’ 05” (W), Parcela # 2: 00º 05’ 03” (S) y 80º 08’ 54” (W). Se registraron un total de 900 individuos distribuidos en 33 Familias, 65 Géneros y 216 especies. Obteniéndose un área basal promedio de 18.10 m2 / Ha. El mayor DAP lo obtuvo el árbol 8.03 de la parcela # 2, Spondias mombin de la Familia Anacardiaceae con un díametro de 122.5 cm y una AB de 1.18. La especie dominante fué Brosimum alicastrum de la familia Moraceae con AB de 2.59 m2 una Dm.R de 14.21 y un IVI promedio de 29.10. La especie que predomina con el mayor número de individuos es la especie Guettarda acreana de la familia Rubiaceae. Con un total de 96 individuos. La familia más importante del bosque fué la Rubiaceae con 207 individuos, con una AB de 4.65, una Dm.R. de 30.20 y un IVI de 48.65. Seguida de la Familia Moraceae con 64 individuos, con una AB de 2.77 una Dm.R. De 15.23 y un IVI de 44.85, concluyéndose que en ambas parcelas existe gran diferencia de especies e individuos, una diferencia de 33 especies, 2 géneros, y 84 individuos de la parcela # 1 en relación a la # 2, mostraron mayor diversidad de especies en la parcela # 1. lo que se atribuyó a la presencia de una zona de transición climática y ecosistémica. Objetivo Identificar taxonómicamente mediante la utilización de claves y estudios de morfología y fisiología a todas y cada una de las especies encontradas en el bosque, determinando cual es la especie dominante y la presencia de nuevas especies. Procedimiento 73 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 En el trabajo de campo fue imprescindible la toma de un sinnúmero de datos, que se concretaron o certificaron en el laboratorio. Relacionado a la ecología y ambiente.Altitud, latitud, humedad, hábito y hábitat de la planta, precipitación, longitud. Se consideró todo árbol con un DAP mayor a los 10 cm. y se caracterizó a las plantas por su morfología principalmente, luego se trató de llegar a las identificaciones taxonómicas en las siguientes jerarquías: Especie, género, familia, orden y clase, altura, diámetro, tipos de corteza, raíz, hojas, tallo y ramas., tipo de sustrato, tipo de látex, resinas, exudados, tipo y color de frutos, tamaño. Además se registrará un número para cada árbol analizado. En el campo, visualmente se determinó características de la planta tales como hábitat, hábito, altura, tipos de estructuras, para la altitud, coordenadas, diámetro, se utilizaron sistemas tecnológicos como altímetros, GPS y cintas diamétricas respectivamente. En el laboratorio para el reconocimiento taxonómico se utilizaron las colecciones del herbario nacional del Ecuador y también sus laboratorios para llegar a determinar todos los individuos en lo posible hasta especie y se llevó un libro de campo con un registro y forma de evaluación para el posterior proceso de datos. El establecimiento de las parcela se inició al levantar una perpendicular de 100 m con relación a un punto cardinal. Siguiendo la dirección de la brújula plantamos jalones cada 10 m medidos con la cinta métrica. Se dejó una piola entre los jalones. Se consideró el punto de inicio como el punto (0) cero. Un giro de 90º y se continuó formando la otra perpendicular del mismo modo y así hasta cerrar el cuadrado de 100 x 100 m. Se procedió a trazar perpendicularmente cada 20 m partiendo desde la línea base del cuadrado y luego las otras líneas horizontales paralelas a la línea base cada 20 m logrando así cuadrar las subparcelas de 20 x 20 m. Todas las subparcelas al igual que las parcelas quedaron demarcadas con piola y jalones. Se procedió a plantar tubos de PVC de 2” de 1,60 m de largo en cada esquina de las subparcelas siendo un total de 36 tubos. Se pintó la parte superior de los tubos con rojo para mayor visualización. Se prepararon las placas numeradas de aluminio con martillos y clavo se inició en la subparcela uno numerando a los árboles que tengan un diámetro la altura del pecho (DAP) mayor a 10 cm previo a esto se tomó el DAP y datos del árbol. Marcado el árbol se procedió a la recolección de muestras principalmente fértiles, es decir, con flores o frutos a tal punto de poder identificar la especie. Se colectaron cuatro duplicados de cada individuo como mínimo, de allí se pudo colectar más según el número de herbarios que va a dársele un duplicado. En el libro de campo se registró el número de colección de cada árbol que no va a coincidir con el número de árbol de las subparcelas. Se dió una preidentificación a la muestra y se depositaron en una funda plástica para ser trasladadas al campamento. En el campamento se dió un tratamiento a las muestras dándole un baño con alcohol ya estando prensadas en periódicos. Una vez terminado de muestrear y coleccionar todos los árboles de las subparcelas se procedió a retirar las piolas y jalones dejando simplemente los tubos PVC pintados. El tratamiento de secado y curación de las muestras se lo realizó en las instalaciones del herbario nacional del Ecuador (QCNE) en Quito. Aquí, llegadas las muestras pasaron a las secadoras permaneciendo 24 horas, para el proceso de identificación en laboratorio y colección del herbario. La utilización de las claves taxonómicas se las realizo según la tecnica del Missouri Botanical Garden. Resultados En una superficie de 20000 m2 se registraron 900 individuos distribuidos en 33 familias, 65 géneros y 216 especies, en el cuadro # 7 se pueden observar las familias y géneros y en el cuadro # 11 constan todos los registros encontrados en las dos parcelas. Un resultado de gran magnitud ha sido el hallazgo de tres nuevos registros para el Ecuador como son las siguientes especies: Amyris pinnata Kunth. Especie de la Familia RUTACEAE, de la cual solo se han encontrado individuos en Centro América, especificamente en Costa Rica y en América del Sur en la región de Tumbez en Perú; Exothea paniculata de la Familia SAPINDACEAE, especie de la cual tambien solo se cuenta con registros en Centro América, convirtiéndose asi en un nuevo registro para el Ecuador; Guettarda acreana de la Familia RUBIACEAE, tambien se convierte en un nuevo registro para la botánica ecuatoriana debido a que no se conocia en nuestros inventarios. 74 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 El área basal promedio de los árboles de este bosque es de 18.10 m2 / hectárea, el registro del mayor DAP lo obtuvo el árbol 8.03 de la parcela # 2, Spondias mombin de la Familia Anacardiaceae con un diámetro de 122.5 cm y una AB de 1.18. La especie dominante en el bosque es Brosimum alicastrum de la familia Moraceae con una AB de 2.59 m2 una Dm. R de 14.21 y un IVI promedio de 29.10. Esto debido a que son árboles de mayor diámetro, sin embargo la especie que predomina con el mayor número de individuos es la especie Guettarda acreana de la familia Rubiaceae. Con un total de 96 individuos. La familia más importante del bosque es la Rubiaceae con un total de individuos de 207, con una AB de 4.65, una Dm. R. de 30.20 y un IVI de 48.65. Seguida de la Familia Moraceae con 64 individuos, con una AB de 2.77 una DmR. De 15.23 y un IVI de 44.85, Tabla 1. Familias y géneros presentes en la superficie de investigación. FAMILIAS ANACARDIACEAE ANNONACEAE ASTERACEAE BIGNONIACEAE BOMBACACEAE BOMBACACEAE BOMBACACEAE BORAGINACEAE CAPPARIDACEAE CARICACEAE CELASTRACEAE CHRYSOBALANACEAE ERYTHROXILACEAE EUPHORBIACEAE EUPHORBIACEAE EUPHORBIACEAE EUPHORBIACEAE EUPHORBIACEAE FABACEAE FABACEAE FABACEAE FABACEAE FABACEAE FABACEAE FABACEAE FABACEAE FABACEAE FABACEAE FLACOURTIACEAE LECYTHIDACEAE MALPIGHIACEAE RHAMNACEAE URTICACEAE # de arbol 8.03 GÉNEROS Spondias Annona Fulcaldia Tabebuia Ceiba Pachira Pseudobombax Cordia Capparis Carica Maytenus Licania Erythroxylum Acalypha Alchornea Phyllanthus Hyeronima Croton Geoffroea Swartzia Mimosa Machaerium Brownea Centrolobium Leucaena Inga vera Erythrina Lonchocarpus Casearia Gustavia Bunchosia Zizyphus Urera FAMILIAS ELEOCARPACEAE MELIACEAE MELIACEAE MORACEAE MORACEAE MORACEAE MYRISTICACEAE MYRTACEAE MYRTACEAE NYCTAGINACEAE NYCTAGINACEAE OLACACEAE PHYTOLACACEAE PHYTOLACACEAE POLYGONACEAS POLYGONACEAS RUBIACEAE RUBIACEAE RUBIACEAE RUBIACEAE RUBIACEAE RUBIACEAE RUTACEAE RUTACEAE SAPINDACEAE SAPINDACEAE SAPINDACEAE SAPOTACEAE SAPOTACEAE STERCULIACEAE VERBENACEAE VERBENACEAE Tabla 2. Los 5 individuos con mayor DAP encontrados. Familia Nombre Cientifico DAP Anacardiaceae Spondias mombin 122.5 75 GÉNEROS Sloanea Trichilia Guarea Brosimum Ficus Sorocea Conpsoneura Eugenia Psidium Neea sp. Pisonia Heisteria Achatocarpus Gallesia Tryplaris Coccoloba Alseis Simira Guettarda Randia Coussarea Faramea Zanthoxylum Amyris Allophylus Exothea Talisia Pouteria Pradosia Guazuma Vitex Aegiphila A. B 1.18 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 18.20 9.12 1.05 12.02 Verbenaceae Moraceae Phytolacaceae Rubiaceae Vitex sp. Brosimum alicastrum Gallesia integrifolia Alseis eggersii 120 99.7 85.9 78.6 Tabla 3. Las 5 familias más importantes encontradas. Familia #individuo A. Basal Dm. R RUBIACEAE 207 4.65 30.20 MORACEAE 64 2.77 15.23 FABACEAE 179 2.85 14.00 SAPINDACEAE 72 1.46 8.04 EUPHORBIACEAE 54 0.41 2.20 1.13 0.78 0.58 0.49 IVI 48.65 44.85 34.25 15.77 8.24 Tabla 4. Las 10 especies más importantes encontradas. # # arbol col. 25.08 249 25.08 276 25.18 468 23.15 402 25.14 207 25.07 274 10.01 383 11.27 275 25.10 452 24.08 303 23.08 340 Familia MORACEAE RUBIACEAE RUBIACEAE FABACEAE RUBIACEAE SAPINDACEAE FABACEAE PHYTOLACACEAE EUPHORBIACEAE FABACEAE SAPOTACEAE Nombre cientifico Brosimum alicastrum Guettarda acreana Simira rubescens Brownea coccinea Alseis eggersii Exothea paniculata Swartzia littlei Gallesia integrifolia Hyeronima sp machaerium millei Pouteria brevipetiolata Ab/sp DmR 2.59 14.21 1.89 10.42 1.26 6.80 0.75 4.00 1.43 7.91 1.26 6.98 1.07 5.88 1.32 6.22 0.34 1.83 0.35 1.95 0.34 1.84 #arb./sp 30 96 49 71 47 46 29 14 41 36 25 IVI 29.10 20.82 13.00 13.00 12.71 11.79 9.12 7.79 6.54 5.67 4.84 Discusión En cuanto a densidad y diversidad de especies en ambas parcelas podemos notar claramente la diferencia de especies e individuos encontrados en ellas, una diferencia de 33 especies, 2 géneros, y 84 individuos de la parcela # 1 en relación a la # 2, hacen pensar fácilmente en una mayor diversidad de especies vegetales presentes en la parcela # 1. Lo que se debe seguramente a la presencia en esta parcela de una zona de transición entre el bosque seco desiduo y el bosque seco siempre verde dentro de la misma reserva. La parcela # 1 se encuentra con un 50 % de superficie en un bosque totalmente seco que no permite la permanencia de las hojas durante todo el año, está ubicada justo en la cima de una colina en donde por el extremo Norte se encuentra una garúa permanente todos los días mientras que por el extremo Sur está permanentemente en contacto con las condiciones áridas causadas por la corriente fría del Humboldt, en esta zona de toda la reserva. Esta área constituye un medio ideal para la variación de especies debido a los siguientes factores: Todos los árboles en esta zona son de baja altura, en promedio aproximado de 15 m. De hojas caducas y semi caducas y de menor diámetro del tallo. Esto permite que la superficie del suelo se mantenga con mayor penetración de luz solar durante el año, a la vez esto conlleva a facilitar la germinación y desarrollo de muchas especies; lo que ha causado que la parcela # 1 cuente con una mayor diversidad y densidad. Referente al mismo fenómeno la parcela # 2 cuenta con una vegetación mucho más exuberante siempre verde, con diámetros mayores a los 100 cm. y con alturas hasta más de 60 metros, de copas muy amplias y una superficie totalmente cubierta de materia vegetal en descomposición y con una 76 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 nula penetración de los rayos solares lo que impide en gran parte la diversificación de las especies a corto tiempo. Ya que la mayor diversidad de la parcela # 2 se las encuentra en las Sub-parcelas en donde existen individuos muertos. Por otro lado a pesar que la parcela # 1 contiene mayor número de individuos, el Área Basal de ambas parcelas se mantiene alrededor de 18.10.m2 / Ha. Esto se debe al aumento del diámetro de los árboles en la parcela # 2; que hacen equilibrar el cálculo de AB. En cuanto a la especie Dominante encontrada el resultado deja muchas incógnitas y que tiene relación directa con lo antes mencionado. Podemos ver claramente en el cuadro # 3 que la especie dominante de la parcela # 1 es Alseis eggersii de la Familia Rubiaceae y a solo mil metros de distancia terrestre, en la parcela dos no encontramos más que un solo individuo de esta especie, lo que hace que pierda la dominancia dentro del bosque. El género Alseis de la Familia Rubiaceae se caracteriza por poseer árboles y arbustos, con estipulas interpeciolares; flores dispuestas en espigas, posee 20 especies en todo el mundo y 2 especies en el Ecuador, de las cuales 1 es endémica; ambas se las encuentra en todo el territorio Nacional. La especie dominante de la parcela # 2 como se ve en el cuadro # 6 es Brosimum alicastrum de la Familia Moraceae. Este en cambio se mantiene en ambas parcelas con menor número que la especie dominante de la parcela # 1, sin embargo obtiene la dominancia gracias a que son árboles de gran diámetro del tallo y por ende de una mayor Area Basal. Lo que le ayuda a convertirse en la especie dominante y más importante dentro del bosque. El género Brosimum se caracteriza por poseer latex blanco acuoso, fuste grisaceo, contiene 14 especies para America Tropical con 8 especies en Ecuador. La familia más grande e importante es la Rubiaceae con un total de 207 individuo, se mantiene presente en las dos parcelas con un AB promedio entre ellas de 4.65m 2/ Ha. Un promedio de IVI de 48.65 y una DmR. De 30.20 %. Lo que nos dice que esta Familia tiene el 48.65 % de importancia en el bosque, que el 30.20 % del total de individuos encontrados en el bosque son especies de esta familia. Y que de cada 10000 m 2, 4.65 m2 están cubiertos por especies de esta familia. La familia Rubiaceae es una de las más grandes contiene 630 géneros con 10400 especies en todo el mundo, 212 géneros con 4500 especies neotropicales, 78 géneros con 499 especies para Ecuador, de las cuales 100 son Endémicas. Se caracterizan por tener hojas simples usualmente enteras, opuestas, verticiladas en algunos géneros, las estipulas interpeciolares son la principal fuente de diagnóstico de esta familia, estan ubicadas una a cada lado del tallo, entre las hojas opuestas, aún después de caer las estipulas se puede observar las cicatrices que dejan en el tallo entre las hojas, existen unas especies excepcionales con estípulas axilares. La flor tiene una corola en forma de tubo con el ovario ínfero. El fruto es usualmente una baya, muy rara vez es capsula o drupa; el fruto tiene una cicatriz circular en el ápice dejado por el caliz. La Rubiaceae es abundante en los bosques húmedos, aunque se encuentran también en menos húmedos y andinos, la mayoría son árboles pequeños. La Familia incluye especies de importancia económica, como el café y la quina. Por otro lado el haber encontrado 33 familias en una superficie de 20000 m2 y con una diferencia de 4 familias entre las parcelas nos dice claramente que en toda la superficie del bosque existen muchas más familias asi como de especies. En la parte ecológica se puede notar fácilmente que 900 individuos en una superficie de 20000 m2 son sumamente significativos aunque no es comparable con los resultados obtenidos en otros bosques de donde se obtienen hasta más de 600 individuos en una sola hectárea. Sin embargo esto nos dice que en esta región de la Costa ecuatoriana existe una gran diversidad de especies, seguramente debido a la influencia que ejercen sobre esta zona las corrientes marinas, lo que hace que se presenten variados microclimas durante el día y en el mismo bosque, esto facilita la adaptabilidad de cualquier especie vegetal del mundo, por las variaciones climáticas permanentes. Bibliografía Bonner, J. y Galston, AW. 1961. Principios de fisiología vegetal. Federico Portillo. Instituto tecnológico de California. Madrid Esp. 2da Ed. Aguilar. p, 13,14 77 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Ceron, MCE. 1993. Manual de Botánica Ecuatoriana. Sistemática y métodos de estudios. Universidad Central del Ecuador. Escuela de Biología. Quito Ecu. p,156. Cevallos, ZJ. 2003. Inventario de la familia Meliaceae en diferentes altitudes de la cordillera galeras. Universidad Laica Eloy Alfaro de Manabí. Quito Ecu. Herbario Nacional del Ecuador. p, 6 Cuamacas, D. 2003. Composición florística de una parcela permanente en la cordillera de Galeras. Universidad Técnica del Norte. Quito Ecu. Herbario Nacional del Ecuador. p,3. Diario La Hora. 2003. Bosques de Manabí. Bosque húmedo. ( en línea) Portoviejo Ecu. 2003 www.dlh.lahora.com Diario Manabita. 2002. Suplemento Manabí por cantones. Pedernales, (en línea) 2003, www.eldiario.com. Hurtado, F; Neill, D; Alvarado, A. s f. inventario cuantitativo de una hectárea de bosque pluvial premontano en la región del volcán sumaco. MO Missouri Botanical garden. Quito Ecu. p, 5. Lopez, VH. 2003. Distribución altitudinal de inga en la reserva de biosfera Sumaco. Herbario Nacional del Ecuador. Quito Ecu. p, 4. Lozano, P; Delgado, T; y Aguirre Z. 2002. Estado actual de la flora endémica exclusiva y su distribución en el occidente del parque nacional podocarpus. Funbotanica, Herbario y jardín botánico Reinaldo Espinoza. P, 9,97. Neill, D. A. 1999. Catálogo de las Plantas vasculares del Ecuador, vegetación. Missouri Botanical Garden, Herbario Nacional del Ecuador, Pontificia Universidad Católica del Ecuador. USA. P, 27. ------- y jorgensen, PM. 1999. Catalogue of the vascular plants of Ecuador. Climas. Missouri Botanical Garden, Herbario Nacional del Ecuador, Universidad Católica del Ecuador. Missouri USA. p, 27. -------. 2003. Curso- pasantia preprofesional en Botánica y conservación. Missouri Botanical Garden, Herbario nacional del Ecuador. Quito- Napo, Ecu. p. 35. Stern, M. 2003. Curso-pasantia preprofesional en Botánica y conservación. Impacto ambiental del cultivo de la naranjilla Solanum quitoense en la zona del sumaco. Missouri Botanical Garden, Herbario Nacional del Ecuador. Quito Ecu. p, 8. 78 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 ESTRUCTURA Y COMPOSICIÓN FLORÍSTICA DEL BOSQUE TROPICAL SIEMPRE VERDE DEL CANTÓN EL CARMEN, MANABÍ Jimmy J. Cevallos Zambrano Universidad Laica Eloy Alfaro de Manabí [email protected] Problema La tala de los bosques es preocupante y avanza a pasos gigantescos lo que conlleva a la extinción de las especies vegetales existentes, la Universidad Laica Eloy Alfaro de Manabí en el Cantón el Carmen mantiene un remanente de bosque siempre verde el mismo que posee una diversidad florística que era hasta hace poco desconocida lo cual era una desventaja para el proceso de protección y conservación de estas especies. Resumen Inventario Botánico del bosque, Determinar la familia botánica dominante, Obtener la densidad de población y Determinar la especie más importante del bosque; fueron los principales objetivos de este trabajo. Los resultados han sido muy considerables ambientalmente, el bosque en estudio mantiene una diversidad florística muy importante abarcando a más de 24 familias botánicas, 41 géneros y 45 especies, esto hace que se considere un bosque muy diverso florísticamente, mantiene un dosel con una altura promedio de 35 m con un sotobosque semidiferenciado y a una altura de 10 m se cuenta con una densidad de población de 546 individuos por hectárea lo cual es considerada una densidad abundante. Cabe destacar que el área basal total del bosque es de 30,1 % y que la familia más abundante y por ende más importante es la Moraceae. Las especies más importantes son: Chivin con un valor de importancia del 45,49 con un área basal por especie de 8,14 y una densidad de 101 árboles por hectárea , una segunda especie en importancia es el jaguillo con una importancia de 41,59 un área basal de la especie de 5,47 y con un número de árboles de 128 por hectárea; una tercer especie en importancia es el pechuga de gallina con un total de 11 árboles por hectárea, con una importancia de 17,40 y un área basal por especie de 4,64; la cuarta especie en consideración es el aspan con 70 árboles por hectárea, un área basal por especie de 1,44 y una importancia de 17,58. La metodología utilizada en este trabajo fue parcelas permanentes de una hectárea considerando a todos los árboles con un DAP superior a 10 cm. Se concluye que el bosque es significativamente importante para su conservación por la abundancia e índice de importancia que representan sus especies. Objetivos La determinación de la biodiversidad del bosque en estudio mediante la realización de un Inventario Botánico del bosque, Determinando la familia botánica dominante, Obtener la densidad de población y a la vez determinar la especie más importante del bosque. Procedimiento A metodología utilizada fue el establecimiento de una parcela permanente de una hectárea, en la misma que se procedió de la siguiente manera: Se procedió a un reconocimiento del área del bosque, con la finalidad de determinar cuál era la zona más idónea en regularidad dendrológica como para desarrollar el presente estudio. Se delimitó una hectárea de terreno con la señalización de piola y jalones. 79 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Una vez establecido el cuadrante se procedió a la subdivisión de parcelas de 20 x 20 m. obteniendo 25 subparcelas de 400m2/u Se desarrolló el inventario botánico en cada subparcela considerando todos los árboles que tenían un DAP superior a los 10 cm. Se dejó una numeración por árbol adherido con cinta y un clavo con la finalidad de garantizar la veracidad de los datos mientras tarde el proceso de tabulación y análisis de los resultados. Se realizó el proceso de los resultados en las oficinas del departamento de medio ambiente en la ciudad de Manta. Para el desarrollo de las actividades se contó con el apoyo de los estudiantes de la extensión, y los técnicos del departamento de ambiente y un guía de campo. Resultados Es importante destacar que el estudio desarrollado se lo hizo dentro del cronograma establecido, logrando determinar la presencia de 546 individuos distribuidos en 24 familias botánicas las cuales engloban a 41 géneros y 45 especies. Entre estas las siguientes: FAMILIAS Verbenaceae Anonaceae Anonaceae Arecaceae Arecaceae Moraceae Moraceae Caricaceae Moraceae Moraceae Cecropiaceae Meliaceae Moraceae Boraginaceae Bombacaceae Fabaceae Bignoniaceae Moraceae Moraceae Meliaceae Tabla 1. Principales familias, Géneros y Especies encontradas. GÉNEROS y ESPECIES GÉNEROS y ESPECIES PRINCIPALES FAMILIAS PRINCIPALES Aegiphila alba Malvaceae Hibiscus elatus Annona cherimola Euphorbiaceae Hieromina sp. Annona muricata mimosaceae Inga edulis Astrocaryum vulgare Arecaceae Iriartea deltoidea bactris gasipaes Lauraceae nectandra acutifolia Brosimum sp. Lauraceae nectandra reticulata Brosimum utile Arecaceae Oenocarpus bataua carica papaya Arecaceae Oenocarpus bataua castilla elástica Mirysticaceae Otoba parvifolia castilla tunu Theaceae Pelliciera rhizophorae cecropia obtusifolia Lauraceae Persea americana Cedrella odorata Cecropiaceae Pourouma sp. Clarisia biflora Sapotaceae Pouteria capacifolia Cordia alliodora Moraceae Pseudolmedia rigida cotostemma comune Araliaceae schefflera actinophylla Erythrina smithiana Ulmaceae Sparrea schippii Exarata chocoensis meliaceae Swietenia macrophylla King Ficus benjamina Poligonaceae Triplaris americana Ficus obtusifolia Asteraceae Vernonia baccharoides Guarea Rutaceae Xanthoxilum tachuelo El bosque mantiene un dosel con una altura superior de 35 m con un sotobosque semidiferenciado y a una altura superior a 8 m. Se determinó una densidad de población de 546 individuos por hectárea la cual es considerada abundante. El área basal total del bosque es de 30,1 % La familia más abundante y por ende más importante es la Moraceae. Las especies más importantes son: Chivin (castilla tunu) con un valor de importancia del 45,49 con un área basal por especie de 8,14 una densidad de 101 árboles por hectárea y una dominancia relativa de 26,99. Una segunda 80 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 especie en importancia es el jaguillo *(Pouteria capacifolia pitz) con una importancia de 41,59 un área basal de la especie de 5,47 y con un número de árboles de 128 por hectárea y un 18,14 de dominancia relativa. Una tercer especie en consideración es el Aspan (Swartzia littlei) con 70 árboles por hectárea, un área basal por especie de 1,44 y una importancia de 17,58 y una dominancia de 4,76. Una cuarta especie en importancia es el pechuga de gallina *(Aegiphila alba) con un total de 11 árboles por hectárea, con una importancia de 17,40 y un área basal por especie de 4,64 y una dominancia relativa de 15,39. Imagen No. 1.- Vista del interior del bosque. Imagen No. 2.- toma de DAP en un representante de la familia Moraceae. 81 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Imagen No. 3.- Arbol de Chivin o cauchillo. castilla tunu, especie dominante del bosque. Discusión Los resultados del presente trabajo han sido muy considerables ambientalmente, el bosque en estudio mantiene una diversidad florística muy importante lo que ha permitido mantener la biodiversidad de fauna abundante asi como de muchos otros organismos. La cantidad de individuo encontrado por unidad de superficie en el bosque es muy importante para tomar la decisión de conservación y emprender nuevas prácticas de sustentabilidad del bosque que permitan a la comunidad vecina suministrarse ciertos productos de manera controlada a través de un plan de manejo. Cabe destacar que en el bosque existe también gran cantidad de caña guadua y otras especies vegetales que no se encontraban dentro de la parcela pero que son muy significativas dentro de esta jurisdicción. El área basal determinada es mayor que la mayoría de los bosques de la costa. La abundancia determinada es superior a la mayoría de los bosques de la costa y con esto se puede ver directamente que su importancia por especie se mantiene sobre la media de la costa también lo cual indica que existe una considerable abundancia y densidad de 456 individuos por hectárea. Se pudo identificar la altura promedio del dosel a unos 35- 45 metros en su mayoría con un subdosel poco diferenciado de manera iregular a unos 10 metros. El sotobosque se encontraba aproximadamente a unos 3-4 metros considerando arbustos en su totalidad, la presencia de malezas fue nula totalmente puesto que estaba dominado por el bosque y sotobosque, la presencia de algunas lianas es también preponderante puesto que se manifestaron de manera continua a través del bosque sin embargo la presencia en la parcela fue muy escaza. Bibliografía Ceron, MCE. 1993. Manual de Botánica Ecuatoriana. Sistemática y métodos de estudios. Universidad Central del Ecuador. Escuela de Biología. Quito Ecu. p, 156. Cevallos, ZJ. 2003. Inventario de la familia Meliaceae en diferentes altitudes de la cordillera galeras. Universidad Laica Eloy Alfaro de Manabí. Quito Ecu. Herbario Nacional del Ecuador. p, 6. Diario La Hora. 2003. Bosques de Manabí. Bosque húmedo. (En línea) Portoviejo Ecu. 2003 www.dlh.lahora.com Diario Manabita. 2002. Suplemento Manabí por cantones. El Carmen, (en línea) 2003, www.eldiario.com. 82 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Hurtado, F; Neill, D; Alvarado, A. s f. inventario cuantitativo de una hectárea de bosque pluvial premontano en la región del volcán sumaco. MO Missouri Botanical garden. Quito Ecu. p, 5. Neill, D. A. 1999. Catálogo de las Plantas vasculares del Ecuador, vegetación. Missouri Botanical Garden, Herbario Nacional del Ecuador, Pontificia Universidad Católica del Ecuador. USA. P, 27. -------. 2003. Curso- pasantia preprofesional en Botánica y conservación. Missouri Botanical Garden, Herbario nacional del Ecuador. Quito- Napo, Ecu. p. 35. Oceano Centrum. sf. Enciclopedia práctica de la agricultura y la ganadería, nociones de Botánica y agro ecología americana. Grupo editorial océano. Madrid Esp. P, 11-36. 83 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 CONTRIBUCIÓN A LA DIVERSIDAD Y SIMILITUD ARBÓREA DE LOS BOSQUES MONTANOS ALTOS DEL ECUADOR CENTRAL Jorge Caranqui Aldaz Herbario Escuela Superior Politécnica del Chimborazo CHEP [email protected] Resumen El presente estudio pretende relacionar la composición arbórea de siete transectos de 0.1 Ha de bosque montano alto en la Sierra central en el Ecuador, 5 transectos en la provincia de Chimborazo y 2 en la provincia de Tungurahua. Se utilizaron las frecuencias y los índices de valor (IV) de las especies encontradas en cada uno de los transectos, con las cuales se obtuvo los índices de diversidad y similitud con el respectivo análisis. Se concluyó que los bosques son heterogéneos y probablemente la altitud sea un patrón de diversidad entre ellos. Hay gran diferencia en la composicion de especies entre los bosques estudiados. Además recomendar las especies abundantes del estudio para los planes de reforestación en la región. Palabra clave: bosque montano alto, abundancia, similitud, diversidad, reforestación, composicion de especies Introducción Los bosques de montaña se caracterizan por una enorme diversidad biológica (tan diversa quizás como la famosa selva tropical lluviosa), pero también por regular los importantes caudales hídricos de los ríos que atraviesan el continente y por sobre todo, por compartir una historia de uso y de oferta de recursos en forma interrumpida con la humanidad, durante por lo menos la última decena de miles de años (Talbot et al.1986). Sin embargo, hoy se presentan como uno de los sistemas más frágiles a la intervención humana y sobre el cual está cayendo con inusual fuerza los procesos de degradación por sobre-utilización y conversión en sistemas agrícolas y campos de pastoreo (Brown & Kappelle 2001). El bosque montano, un tipo de vegetación donde se esperan altos niveles de diversidad, debido a las barreras biogeográficas que son tan frecuentes en las montañas. La diversidad y composición florística son los atributos más importantes para diferenciar o caracterizar cada complejo y/o comunidad vegetal (Araujo-M. 2005). Por otro lado, la composición florística de un bosque es determinada por el conjunto de especies de plantas que lo componen y es tradición medirla considerando la frecuencia, abundancia o dominancia de las especies (Whittaker, 1975 ). Los bosques montanos altos del centro del Ecuador carecen de estudios especialmente florísticos (Caranqui 2011), En su estudio Jørgensen et al. (1996) manifiestan que entre los bosques del norte y del centro del Ecuador hay similitud florística, no así con los del sur del Ecuador. Por lo manifestado anteriormente, el objetivo de este trabajo es contribuir al conocimiento de la diversidad y similitud de la vegetación mayor de 5 cm de diámetro a la altura del pecho (DAP), encontrada en 7 transectos de bosque montano en el Centro del Ecuador. Métodos Área de estudio Este estudio se realizó en siete bosques montanos (Sierra 1999), localizados en las provincias de Chimborazo y Tungurahua (cuadro 1): 84 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Cuadro 1. Ubicación de los transectos en el bosque ceja de montaña en el centro del Ecuador LOCALIDAD Tambopalictahua 1 Tambopalictahua 2 ALTITUD (msnm) 3410 2780 COORDENADAS CANTÓN PROVINCIA 01.30.25S, 78.27.04W 01º30'25"S 78º27'04"W Penipe Chimborazo Penipe Chimborazo San Francisco 3600 01.47S, 78.34W Chambo Chimborazo Llucud 3400 Chambo Chimborazo Bacun 3180 01.43.12S, 78.32.32W 02.17’S, 78. 53W Chunchi Chimborazo Quimbana 3750 01.10S,78.28W Pillaro Tungurahua Leito 3300 01°19'42"S 78°27'47"W Patate Tungurahua En el cuadro 1 se encuentran las zonas de muestreo localizadas en la cordillera oriental y están ubicadas a altitudes superiores de 3000 msnm., a exepción de Tambopalictahua 2 (2780m.). Ninguno de estos bosques se encuentran dentro del Sistema de Área Protegidas del Ecuador y no sobrepasan las 80 Ha de extensión cada una, y todos en mayor o menor proporción existen fragmentación por estár amenazados por actividades humanas como cultivos y zonas de pastoreo. Toma de datos En cada sitio se instaló una área muestreal de 0.1 hectárea, divididos en 5 transectos de 50 x 4 m que es variante de los transectos realizados de Gentry (Phillips & Miller 2002, Cerón 2003), ya que estos eran de 10 juegos de transectos de 50 x 2m. Utilizando transectos amplios reduce los efectos de micro hábitats en los resultados generales, y tomando en cuenta la fragmentación del bosque que en la actualidad ocurre, los transectos de 50 x 4m nos ayudan a estandarizar el área de muestreo. Los siete transectos son del mismo tamaño lo que permite una comparación entre ellos. El trabajo de campo se realizó en varias fechas del 2009, 2010 y 2011. Se tomó el DAP (diámetro a la altura del pecho) de las especies mayores de 5 cm. Se colectaron especímenes botánicos de la mayoría de los individuos marcados (incluyendo todas las especies no definitivamente identificados en el campo) un duplicado para muestras infértiles y tres para muestras fértiles. Las muestras están depositadas en el Herbario de la Escuela Superior Politécnica de Chimborazo (CHEP). Para mayor información de las especies encontradas se revisó el Catálogo de Plantas Vasculares (Jørgensen & León-Yánez, 1999) y la actualización se consultó en la base de datos Trópicos (www.tropicos.org) del Missouri Botanical Garden. Análisis de datos Se realizaron los siguientes cálculos (Cerón 2003): AB= π(D)²/4 85 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Donde AB=Área basal D= DAP [Diámetro a la altura del pecho (1.30m.)] Π= 3.1416(constante) Densidad = Número de árboles en la parcela Densidad Relativa (DR)= (número de árboles de una especie/número de árboles en la parcela)*100 Dominancia Relativa (DMR)= (Área basal de una especie/Área basal total de todos los árboles en la parcela)*100 Índice de Valor (IV)= DR + DMR Además se generó un listado general de especies con sus respectivas frecuencias en cada uno de los transectos, con los cuales se obtuvo: riqueza, diversidad (índice de Simpson), similitud (índice de Bray Curtis), calculados en el programa PAST. Los análisis que se presentan a continuación fueron tomados de los resultados de estudios de composición y estructura realizados por Caranqui (2011), y las tesis realizadas por Ortiz (2010) y Paucar (2011). Resultados Cuadro 2. Listado de especies con sus respectivos IV en los 7 transectos Familias Especies ACTINIDACEAE Saurauia tomentosa (Kunth) Spreng. ARALIACEAE ARALIACEAE ASTERACEAE ASTERACEAE ASTERACEAE ASTERACEAE Oreopanax ecuadorensis Seem. Oreopanax seemannianus Adenostema harlingii R.M.King & H.Rob. Aequatorium asterotrichum B.Nord. Baccharis latifolia (Ruíz & Pav.) Pers. Critoniopsis sodiroi (Hieron.) H.Rob. ASTERACEAE Dendrophorbium tipocochensis (Domke) Nord. ASTERACEAE ASTERACEAE ASTERACEAE ASTERACEAE ASTERACEAE BERBERIDACEAE BERBERIDACEAE Grosvenoria campii R.M.King & H.Rob. Gynoxys sp Munozia jussieui (Cass.) H.Rob. & Brettell Verbesina cf. nudipes S.F.Blake Verbesina latisquana S.F.Blake Berberis glauca DC. Berberis sp BETULACEAE BORAGINACEAE BUXACEAE Alnus acuminata Kunth Tournefortia fuliginosa Kunth Styloceras laurifolium (Willd.) Kunth 86 TP 1 TP 2 SF LL LE QUI BA 16,2 0 4 0 0 0 0 0 13,5 3,39 4 0,41 3,28 1,38 0 1,26 0 0 0 0 0 1,13 0 0 0 0 0,82 0 0 0 0 0 0 0 0 5,96 0 0 0 0 0 0 1,36 0 0 0 0,41 0 0,3 0 7,72 12,0 0,52 0 0 0,82 8 0 0 11,4 12,1 0 0 8,08 8 0 0,95 4 0 0 1,74 4,1 0 4,19 0 0 0 0,58 0 0 0,71 0 0 1,17 0 0 0 0 0 0 0 4,61 0 2,94 0 0 0 0 0 0 0 0 0,39 0 0 0 0 0 0,22 0 45,2 0,31 0 0 0 3 0 0 0,52 0 0 0 5,56 0 1,01 0 0,65 0 0 0 0 10,9 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 CANNABACEAE Trema micrantha (L.) Blume CELASTRACEAE Maytenus verticillata (Ruíz & Pav) DC. CHLORANTHACEA E Hedyosmun cumbalense H.Karst CUNNONIACEAE CUNNONIACEAE CYATHEACEAE ELAEOCARPACEA E ERICACEAE 0 0 0 0 0,39 0 0 0 5 0 2,66 0 0 0 0 0 0 0 0 0 0 0 Weinmania mariquitae Szyszyl. Weinmania pinnata L. Cyathea cuatrecasana(Klotzch) Domin 0 0 0 0 17,7 8 0 69,6 5 0 0 2,1 0 2,54 Vallea stipularis L.f. Macleania cordifolia Benth 3,01 0,26 0 0 1,25 1,64 0 0 0 0 0 0 0 0 0 1,04 0 1,7 0 0 Miconia jahnii Pittier 0 0 1,23 0 42,1 18,0 6 3 36,1 26,2 5 3 0 5,27 0 Miconia latifolia (D.Don) Naudín 0 0 0 0 0 2,54 Miconia pseudocentrophora Cogn. 0 0 0 0 5,91 0 0 12,6 5 2,93 0 0 7,37 0,67 0 0 0,31 0 0 0 0 0 0,41 0 0 2,52 0 0 7,52 0,81 0 0 0 0 0 0 0 1,25 0 0 0 0 0,86 8,2 0 0 0 12,3 0 0 0 2,46 0 0 0,84 0 0 0 0 0 0,3 0,6 0 0 0 0,62 0 0 2,89 0 0 0,82 0 14,4 5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2,38 0 0 0 3,26 0 0 0 0 0,65 0 0 0 0 0 0 2,3 0,44 0 0 1,22 0 0 0 0 0 0,32 0 0 68,1 0,52 0 0,83 2,46 0 5 0 0 0 0 0 0,91 0 0 40,6 0 2,84 0 0 0 0 5 24,2 0 3 0 0 0 0 0 ESCALLONIACEAE Escallonia myrtilloides L.f. LAURACEAE Nectandra sp. LAURACEAE MELASTOMATACE AE MELASTOMATACE AE MELASTOMATACE AE MELASTOMATACE AE MELASTOMATACE AE MELASTOMATACE AE MELASTOMATACE AE MELIACEAE MYRTACEAE ONAGRACEAE PIPERACEAE PRIMULACEAE PRIMULACEAE PRIMULACEAE PRIMULACEAE PRIMULACEAE ROSACEAE ROSACEAE ROSACEAE RUBIACEAE RUBIACEAE S/F Ocotea floribunda (Sw.) Mez SABIACEAE Meliosma arenosa Idrobo & Cuatrec. Axinaea quitensis Benoist Brachyotum ledifolium (Desr.) Triana Miconia bracteolata (Bonpl.)DC. Miconia theazaens (Bonpl.) Cogn. Cedrela montana Moritz ex Turcz. Myrcianthes rhopaloides (Kunth) Mc Vaugh Fuchsia loxensis Kunth Piper nubigenum Kunth Ardisia sp. Geissanthus ecuadorensis Mez Geissanthus pichinchae Mez Myrsine andina (Mez) Pipoly Myrsine coriacea (Sw.)R.Br. Ex Roem. & Schult. Hesperomeles ferruginea (Pers.) Benth Hesperomeles obtusifolia (Pers.) Lindl. Prunus huantensis Pilg. Notopleura sp. Palicourea amethystina (Ruíz & Pav.) DC. Indeterminada 87 0 0 0 0 0 0 0 0 1,06 0 0 0 1,18 0 0 0 0 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 SIPARUNACEAE SOLANACEAE SOLANACEAE Siparuna muricata (Ruíz & Pav.) A.DC. 0 1,89 0 0 0 0 Cestrum peruvianum Willd. Ex Roem. & Schult. 0 0,58 0 0 0 0 Iochroma fuchsioides (Bonpl.) Miers 0 1,29 0 0 0 0 Sessea corymbiflora Goudot ex. Rich. Taylor & SOLANACEAE R.Phillips 0 2,75 0 0 0 0 SOLANACEAE Sessea vestita (Hook.f.) Miers. 0 0 0 0,82 0 0 SOLANACEAE Solanum oblongifolium Dunal 0 0 0 0 0 0 SOLANACEAE Solanum sp1 0,26 0 0 0 0 0 SOLANACEAE Solanum sp2 0 0 0 0 0 0 SOLANACEAE Solanum stenophyllum Dunal 0 0 0 0 0 4,43 SOLANACEAE Solanum venosum Dunal 0,52 1,78 0,42 6,56 1,7 4,65 URTICACEAE Boehmeria ramiflora 0 1,82 0 0 0 0 URTICACEAE Phenax rugosus (Poir.) Wedd. 0 0 0 0 0 0 VERBENACEAE Aegiphila monticola Moldenke 0 2,75 0 0 0 0 TP1:Tambopalictahua1,TP2:T.Palictahua2,SF:San Francisco,LL: Llucud, LE: Leito, QUI:Quimbana,BA:Bacún. 0 0 0 0 0 0,37 0 0,33 0 1,07 0 2,28 0 En el cuadro 2 se encuentra 65 especies distribuidas en los 7 transectos. De las cuales solo Solanum venosum tiene presencia en todos los transectos pero que sus IV no son altos, esto puede indicar que esta especie se encuentra en parches alterados, le sigue Oreopanax ecuadorensis con presencia en 6 de 7 transectos y Miconia bracteolata, Grosvenoria campii, Escallonia myrtilloides y Vallea stipularis con presencia en 4 de los 7 transectos, el resto de especies tienen menor presencia en los 7 transectos. De la tabla 2 es evidente que la especie dominantecon respecto a IV es diferente en cada parcela. Además, cada parcela típicamente tiene solamente una especie con un IV de más de 20. La excepción a esto es la aparición de dos especies de Miconia en el SF y LL con alto IV. Cuadro 3. Datos de los siete transectos de taxones, abundancia y diversidad. Familias Géneros Especies Individuos Indice Simpson Tambo Tambo San Palictahua 1 Palictahua 2 Francisco Llucud Leito Quimbana Bacún 13 17 9 9 14 6 15 13 23 13 12 17 10 15 14 22 15 15 18 13 17 194 85 121 122 175 236 155 0,69 0,91 0,79 0,85 0,84 0,73 0,86 En el cuadro 3 se aprecia los taxones de familias, géneros y especies, varían especialmente con relación a la altitud ya que los máximos valores recae en T. Palictahua 2 que está a 2780m, y los menores valores corresponden a la localidad de Quimbana con la altiud mas alta que es de 3750m. El número de individuos al contrario tiene un crecimiento con la altitud, pero en los transectos intermedios no hay una tendencia marcada. 88 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figura 2. Fenograma del índice de Bray Curtis con los 7 transectos La figura 2 indica que solo Llucud y San Francisco son afines por que fueron realizadas en la misma zona y tienen un rango igual de altitud (±200m.), el resto de transectos no tienen afinidad entre sí. Tambo Palictahua 1 es el que menos afinidad tiene tal vez es porque es el único transecto de menos de 3000 m. Y en este caso no tienen relación a T.Palictahua 2 de la misma localidad como el caso anterior, ya que los dos transectos de Tambo Palictahua tienen diferente altitud y eso puede influir en la diferente composición de especies, ya que comparten solo dos especies y por eso los resultados. Discusión El índice de diversidad de Simpson (Cuadro 2) indica la relación entre riqueza o número de especies y la abundancia o número de individuos por especies en cualquier sitio dado (Smith 2001). En los transectos encontré 14 a 22 especies en cada 0,1 Ha. coincidiendo con otros estudios en donde se han encontrado de 14 a 23 especies (Araujo-Murukami 2005). Gentry en sus transectos de 0.1 Ha a más de 3000m en Ecuador encontró 40 especies en Corazón-Carchi y 37 en Pasochoa, Pichincha (Phillips 2002). Sin embargo, Gentry utilizó un diseño de corte transversal más disperso (dos transectos de 2x50 m para cada transecto individual de 4x50 m en el presente estudio), lo que explicaría al menos en parte, algunos de los mayores niveles de diversidad reportados en sus estudios. El patrón general, sin embargo, de una menor diversidad en estos bosques es coherente con Jorgensen (1995), que manifiesta según sus parcelas instaladas en el norte de los Andes son pobres en especies. La altitud puede ser considerada como el principal factor que influye la diversidad pero la falta de estudios de ese factor en los bosques montanos no se ha podido confirmar con certeza (Gentry 1995, Lieberman 1996). Por tal razón, enunciamos que en nuestro caso hay una tendencia, ya que el menor número de especies corresponde a la localidad de Quimbana que se localiza a 3750m (altitud alta), y el mayor número de especies corresponde a Tambopalictahua2 con 2780m (altitud baja), lo que coincide con la regla RAPOPORT¨S (Bhattarai 2006), que indica que a menor altitud mayor número de especie y a mayor altitud menor número de especies. Pero por tener solo 1 localidad debajo de los 3000m, no podríamos asegurar completamente este enunciado. Kessler & Beck (2001) mencionan a Clethra, Clusia, Escallonia, Freziera, Gaultheria, Hedyosmum, Hesperomeles, Miconia, Morella, Oreopanax, Podocarpus, Prumnopitys, Symplocos y Weinmannia como los géneros dominantes de bosque montano neotropical entre 2.500 y 3.500 m. En nuestro estudio tenemos 9 de 14 géneros enunciados, también se debería incluir en la lista de géneros importantes a Myrsine y también incluir elementos importantes como por ejemplo Schefflera, Chusquea, Berberis, Ilex, Brunellia, Buddleja y Vallea , en este caso tenemos 4 de los 7 géneros. Es relevante mencionar que la especie Miconia theizans es muy importante en los bosques montanos desde Ecuador (Pasochoa) hasta el norte de Bolivia. En nuestro caso tenemos a Miconia bracteolata con buena presencia en los transectos de San Francisco y Llucud, que no se tiene mayores datos de esta especie. Es interesante observar que a nivel de familia de plantas, sólo Melastomataceae se representa una de las especies dominantes en más de un transecto, con especies de Miconia y Axinaea dominantes en los sitios ubicados en elevaciones inferiores (TP2) y superior (SF, LL) . 89 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 En cuanto a familias corroboramos también con Jorgensen (1995), tenemos Asteraceae con el 15,4% de las especies, Solanaceae con el 13,8%, Melastomataceae con el 10,8% y Primulaceae con el 7,7%; es decir estas 4 familias contienen el 48% del total de familias de las 29 en estudio. Por otro lado, las familias representadas por las especies dominantes son en su mayoría diferentes de las familias con mayor diversidad de especies. Las familias con las especies dominantes son: Cunnoniaceae (TP1: Weinmania mariquitae, IV = 69,65), Escalloniaceae (QUI: Escallonia myrtilloides, IV = 68,15), Betulaceae (LE: Alnus acuminata IV = 45.23), Lauraceae (BA: Ocotea floribunda, IV = 40,65), y Melastomataceae (SF: Miconia bracteolata, IV = 42,16; SF & LL: Miconia jahnii, IV = 36,15 y 26, 23; TP2: Axinaea quitensis, IV = 24,23). Sólo Melastomataceae está representada tanto entre las más altas familias diversidas de especies que de las familias de especies dominantes. En la zona central del Ecuador tienen especies representativas tomando en cuenta sus IV (cuadro 1), como Miconia bracteolata (1,04; 0; 42,16; 18,03; 1,7;0;0), Miconia jahnii (0; 0; 36,15; 26,23; 0; 5,27; 0), Escallonia myrtilloides (0,52; 0; 0,83;2,46; 0; 68,15;0) , Weinmania mariquitae (69,65; 0; 0; 0; 2,3; 0,44; 0), Alnus acuminata (0,31; 0; 0; 0; 45,23; 0; 0), Ocotea cf. floribunda (0; 2,84; 0; 0; 0; 0; 40,65), Axinaea quitensis (0; 24,23; 0; 0; 0; 0;0), que están presentes pero no significativamente en la mayoría de los transectos, talvez por la fragmentación que sufren los bosques montanos podría ser otra razón de aislamiento de las especies dominantes de los bosques (Kattan & Alvarez 1996). Que la especie dominante en cada transecto tenga un alto IV y no se repite la especie mayoritaria entre transectos estudiados, nos hace relacionar a la teoria de biogeografía de islas sostenidad por McCartur & Wilson en 1963, ya que cada isla, en este caso, cada bosque tiene sus especies dominantes características, y en nuestro estudio, especialmente por factores antropogénicos pueden en un futuro disminuir los valores encontrados o cambiar de especies dominantes. Probablemente en los bosques montanos arriba de los 3000 m., en la zona central del Ecuador tienen varias especies representativas, además por la fragmentación de bosques que sufren los bosques montanos podría ser otra razón de aislamiento de las especies dominantes de los bosques (Kattan & Alvarez 1996). En nuestro estudio no se diferenciaría si las especies son producto de un bosque secundario maduro o de un bosque primario, para eso se necesitaría seguimientos de largo plazo para saber exactamente la dinámica de los bosques montanos del centro del País (Caranqui 2011). Parece probable que los remanentes del verdadero bosque primario en los Andes del norte y centro de Ecuador son extremadamente raros. Conclusiones Por los datos aquí presentados, podemos manifestar que los bosques nativos son recursos importantes para la reforestación por tener una diversidad de especies nativas, en lugar de lo que se realiza actualmente, usar especies exóticas que nunca estuvieron en nuestro paisaje y además causa el desconocimiento en el uso de especies nativas (Brandbyge 1991), y lo confirma Suarez (2008), ya que manifiesta que, la recuperación de los bosques de Ceja Andina entre los fragmentos que actualmente existen es difícil. Hasta la fecha no se tiene suficiente información para hacerlo y gente tomadora de decisión, continúan con grandes campañas de reforestación, que están formando cualquier cosa menos bosque andino, en las cuales incluso en su planificación, no se está pensando en los remanentes existentes. Además estos fragmentos de bosque andino son cada uno muy diferente, y relativamente única en la dominancia de las especies y diversidad. Por lo tanto, para fines de conservación es fundamental que se proteja el máximo número posible de fragmentos de bosque, ya que cada fragmento puede representar distintos elementos de la biodiversidad regional. Por otra parte, los esfuerzos para restablecer las especies nativas en proyectos de reforestación deben hacer hincapié en la reintroducción de muchas especies diferentes de árboles, a fin de reflejar la gran diversidad de especies dominantes representadas en diferentes fragmentos de bosque. Además de los daños que puedan causar en la retención de humedad y fertilidad del suelo como la biodiversidad (Farley 2004; van Wesenbeek 2002) pero de este tiempo a esta parte no se han realizado estudios de largo plazo para utilizar especies nativas, y prácticamente del discurso no ha pasado. Agradecimientos 90 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Tim McDowell, David Suarez, Carmen Ulloa , Michael Melampy, Issau Huamantupa y Elsa Toapanta por la revisión del documento y sus importantes aportes. Además a los estudiantes de Ingeniería Forestal de la ESPOCH, por las salidas de campo. Bibliografía Araujo-Murakami, A. 2005. Estructura y diversidad de plantas leñosas en un bosque amazónico preandino en el sector del Río Quendeque, Parque Nacional Madidi, Bolivia. Ecología en Bolivia, Vol. 40(3): 304-324 Bhattarai, K. & Vetaas, O. 2006. Can Rapoport´s rule explain tre species richness along the Himalayan elevation gradient, Nepal?. Diversity and distributions. 12,373-378. www.blackwellpublishing.com/ddi. Brown, A. y M. Kappelle 2001 Introducción a los bosques nublados de Latinoamérica. Una Síntesis regional. Pp. 25–40. En: Kappelle, M. & A. D.Brown (eds.) Bosques Nublados del Neotrópico. Instituto Nacional de Biodiversidad (INBio), Santo Domingo de Heredia. Brandbyge, J. and L. B. Holm-Nielsen. 1991. Reforestación en los Andes ecuatorianos con especies nativas. CESA. Quito, Ecuador. Caranqui, J. 2011. Estudios básicos de bosques montanos en el centro del Ecuador. Editorial Académica Española. 67 páginas. Publicado en Alemania Ceron, C. 2003. Manual de Botánica, Sistemática, Etnobotánica y Métodos de Estudio en el Ecuador. Herbario “Alfredo Paredes” QAP, Escuela de Biología de la Universidad Central del Ecuador. Farley, K.A., E.F.Kelly., R.G.Hofstede., 2004. Soil organic carbon and water retention following conversion of grasslands to pine plantations in the Ecuadorian Andes. Ecosystems 7 (7) Gentry, A.H. 1995. Patterns of diversity and floristic composition in neotropical montane forest. Pp. 103-126. In: Churchill, S.P., H. Balslev, Forero,E., & LuteynJ.L. (eds), Biodiversity and conservation of neotropicals montane forests. The New York Botanical Garden, Bronx Jørgensen, P.M. y S. León-Yánez (Eds.) 1999. Catalogue of the Vascular Plants of Ecuador. Missouri Botanical Garden. Kattan G., H. Alvarez. 1996. Preservation and management of biodiversity in fragmented landscape in the Colombian Andes, p.3-18. En: J. Schelhas & Greenberg (eds.) Forest patches in tropical landscapes Island Press Washington D.C. Kessler, M. & S.G. Beck. 2001. Bolivia. Pp. 581–622. En: M. Kappelle & A.D. Brown (eds.)Bosques nublados del Neotrópico. Instituto Nacional de Biodiversidad, InBio, Santo Domingo de Heredia. Larrea M. 1997. Respuesta de las Epifitas vasculares a diferentes formas de manejo de bosque nublado. En Mena, P.A., A. Soldi, R. Alarcón, C. Chiriboga & L. Suarez (Eds.) Estudios Biológicos para la Conservación . Diversidad, Ecología y Etnobiología. Ecociencia. Quito. Pp. 321-346. Lieberman, D., Lieberman, M., Peralta,R. & Hartshorm, G.S. 1996. Tropical Forest structure and composition on a largescale altitudinal gradient in Costa Rica. J.Ecol. 84: 137-152. Ortiz, M. L. 2010. Estructura y composición en el bosque montano de Quimbana, Tungurahua. ESPOCH. Tesis de grado. Escuela de Ingeniería Forestal. Riobamba – Ecuador. Paucar, M.G. 2011. Estructura y composición en el bosque montano de Leito, Tungurahua. ESPOCH. Tesis de grado. Escuela de Ingeniería Forestal. Riobamba – Ecuador. Phillips, O. y J. S. Miller. 2002. Global patterns of plant diversity: Alwyn H. Gentry's forest transect data set. Monographs in Systematic Botany from the Missouri Botanical Garden 89: 1–319. 91 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Valencia, R. & P. M. Jorgensen. 1992. Composition and structure of a humid montane forest on the Pasochoa volcano, Ecuador. Nordic J.Bot. 12: 239- 247. Sierra, R. 1999. Propuesta preliminar de un sistema de clasificación de vegetación para el Ecuador continental. Quito – Ecuador. Suarez D. 2008. Formación de un corredor de hábitat de un bosque montano alto en un mosaico de páramo en el norte del Ecuador. Ecología Aplicada, 7 (1,2). Departamento académico de Biología, Universidad Nacional Agraria La Molina, Lima-Perú. Smith L. & T. Smith. 2001. Ecología. Pearson Education. Madrid. Pag. 664. Talbot, R. W., R. C. Harris. E. V. Browell, G. L. Gregory, D. I. Sebachery, S. M. Beck, 1986. Distribution and Geochemistry of Aerosols in the Tropical North Atlantic Troposphere: Relationship to Saharan Dust. Journal of Geophysical Research 91:5173-5182. Trópicos base de datos. Missouri Botanical Garden. [Consulta de internet 31 Mar. 2011] http://www.tropicos.org van Wesenbeeck, B.K., T. van Mourik, J.F. Duivenvoorden and A.M. Cleef. 2003. Strong effects of a plantation with Pinus patula on Andean subpáramo vegetation: a case study from Colombia. Biol. Conserv. 114:207–218. Whittaker, R.H. 1975. Communities and ecosystems. MacMillan Publishing. Nueva York. 385 p. 92 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 DIVERSIDAD Y COMPOSICIÓN FLORÍSTICA EN LA ESTACIÓN CIENTÍFICA RÍO PALENQUE Jorge Caranqui Aldaz Herbario Escuela Superior Politécnica del Chimborazo CHEP, Universidad Técnica de Quevedo UTEQ [email protected] RESUMEN La presente investigación se llevó a cabo en el bosque siempreverde de tierras bajas, ubicado en la Estación Científica Río Palenque propiedad de la Fundación Wong. El objetivo de este trabajo es contribuir al conocimiento de la diversidad y composición florística de plantas arboreos mayor de 10 cm de diámetro a la altura del pecho (DAP), además establecer el estado de los bosques base de la composición y del número de especies con sus frecuencias encontradas en 5 transectos de 1000 m² de bosque. Se encontraron 78 especies, 58 géneros y 29 familias distribuidas en los 5 transectos, en cada transecto varía de 16 a 26 especies por transecto, el índice de diversidad de Simpson se aproxima a 1(0.91-0.94), en este caso no nos permite distinguir que transecto es el más diverso. La mayoría de las especies no están presentes en todos los transectos, el número de individuos en cada transecto no es representativo. En cuanto al índice de similitud el transecto 5 es diferente que los otros 4, con esto se puede inferir que el transecto 5 pertenece a un bosque alterado y los otros a bosques maduros homogeneos. Palabra claves. Rio Palenque, bosque siempreverde, biodiversidad, similitud INTRODUCCION Según Dodson y Gentry (1987), el bosque húmedo Tropical de la Costa es altamente heterogéneo y diverso, con un dosel que alcanza los 40 m de altura y árboles emergentes que superan los 55 m o más de altura. El Centro Científico Río Palenque, es parte de los bosques húmedos occidentales ecuatorianos, ricos en especies, poseen alto endemismo local y regional. La biota comparte grandes afinidades además con los bosques Tropicales de hoja ancha conocidos como la Ecoregión del Chocó-Darien rico en diversidad y endemismo (Dinerstein et al., 1995). Desde su establecimiento como Centro Científico, Río Palenque ha sido objeto de múltiples estudios e investigaciones con el propósito de ampliar los conocimientos de su fauna y flora. Dodson and Gentry (1987) publicaron "Flora de Río Palenque", donde se identificaron 1100 especies de plantas vasculares de diversas especies, de las cuales 30 son endémicas de este bosque. Por lo manifestado anteriormente, el objetivo de este trabajo es contribuir al conocimiento de la diversidad y composición florística de la vegetación mayor de 10 cm de diámetro a 1.30 m sobre el suelo (DAP), encontrada en 5 transectos (unidades muestreales) del bosque húmedo Tropical en el Centro Científico Río Palenque y establecer los estados de bosques que pueden existir en base del número de especies y sus abundancias. MATERIALES Y MÉTODOS La presente investigación se llevó a cabo en el bosque siempreverde de tierras bajas (Sierra, 1999), ubicado en la estación científica Río Palenque (Figura 1), propiedad de la Fundación Wong la misma que se encuentra ubicada en el km 55 de la vía Quevedo – Santo Domingo cerca del límite entre las provincias de Los Ríos y Santo Domingo de Los Tsachilas, siendo su ubicación geográfico 00°35´19” de latitud Sur y 79°21´45” longitud Oeste a una altitud de 196 msnm, con una temperatura media anual que oscila entre 23 y los 25.5º C, recibe una precipitación promedia anual entre 2000 a 2500 mm, el suelo posee un horizonte B de color blanco amarillo muy seco. La Estación Río Palenque 93 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 comprende un parche de alrededor de 115 ha de bosque siempreverde de tierras bajas, y 10 ha de macadamia y árboles frutales. Figura 1. Ubicación de la zona de estudio (marca roja) Fuente: Cartografía Forestal de REYBANPAC. C.A. Los equipos y materiales utilizados fueron: piola, machete, libreta de campo, computadora, software PAST, papel periódico, secadora de plantas, etc. En el mes de agosto del 2012, se procedió a inventariar 5 transectos de 1000 m 2, en forma de zig-zag con cinco líneas de 50x4 m cada una con rumbo al Norte (Caranqui, 2011a; Caranqui, 2011b; Cerón, 2003; Phillips, 2002). De las cuales 4 están en bosque maduro y una en alterado. Se midieron todos los árboles mayores a 10 cm de DAP. Se colectaron especímenes de la mayoría de los individuos marcados. Las muestras se prensaron y fueron secadas e identificadas en el Herbario de la Escuela Superior Politécnica de Chimborazo (CHEP), y verificado la información en Jorgensen (1999) y en la base de datos del Missouri Botanical Garden (2013) A partir del inventario se consiguieron las especies presentes en cada uno de los transectos con sus respectivas frecuencias con los que se obtuvieron: riqueza, diversidad (índice de Simpson), índice de similitud (Bray Curtis), calculados en el programa PAST. Según Golicher (s/f), la fórmula para el índice de Simpson es: Donde S es el número de especies, N es el total de organismos presentes (o unidades cuadradas) y n es el número de ejemplares por especie. El índice de Bray y Curtis (1957), es uno de los más ampliamente utilizados en la ecología cuantitativa actual y sus expresiones de similitud y disimilitud son: Sjk = 2 Σ min (Xij , Xik)/ Σ (Xij +Xik)Djk = Σ |Xij - Xik|/ Σ (Xij + Xik) Algunos datos fueron tomados de los trabajos de Composición y Estructutra en Río Palenque realizado por Mata et al. (2012); Salazar et al. (2012); Barragán et al. (2012) y Aguilar et al. (2012), dentro de la cátedra de Dendrología en la Maestría de Manejo y Aprovechamiento Forestal. Cuadro 1. # Listado de especies con sus respectivas abundancias en los 5 transectos Especies T1 T2 T3 94 T4 T5 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 Rollinia mucosa (Jaqc.) Baill. Dendropanax umbellatus (Ruíz & Pav.) Decne & Planch. Synechantus warscewiczianus H.Wendl. Caparis ecuadorica H.H.Iltis Cyathea carecasana (Klotzsch) Domin Acalypha alopecuroides Jacq. Browneopsis disépala (Little) Klitgaard Inga sp. Abarema macradenia (Pittier) Barneby & J. W. Grimes Caryodaphnopsis theobromifolia (A.H. Gentry) van der Werff & H.G.Richt. Grias sp. Ossaea micrantha (Sw.) Macfad. Ex Cogn. Guarea kunthiana A.Juss. Perebea guianensis Aubl. Ficus tonduzii Standl. Ficus sp. Clarisia sp. Virola sp. Otoba novagranetensis Moldenke Zyzyphus sp. Pentagonia macrophylla Benth. Chrysophyllum venezuelanense (Pierre) T.D.Penn Pouteria sp. Lycianthes inaequilatera (Rusby) Bitter Cecropia obtusifolia Bertol. Cecropia sp. Alchornea leptogyna Diels Bactrys coloniata L.H.Bailey Brosimum utile (Kunth) Pittier Carapa guianensis Aubl. Carica pubecens Lenné & C.Koch Clarisia racemosa Ruíz & Pav. Erytrina edulis Triana ex Micheli Maclura tinctoria (L.) Steud. Miconia barbinervis (Benth.) Triana Naucleopsis sp Ochroma pyramidale (Cav. ex Lam.)Urb. Piper obtusilimbum C.DC. Piper sp Randia sp. Stylogyne serpentina Mez Tabernaemontana amigdalifolia Jacq. Wettinia sp Brownea sp Castilla elástica Sessé Ficus macbride Standl 95 1 1 1 1 2 1 1 1 1 1 1 1 1 3 4 2 5 1 2 2 1 1 2 1 1 3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 2 0 2 0 0 0 0 2 0 0 0 0 0 1 1 5 0 4 3 1 2 1 1 2 1 2 1 2 1 1 1 2 1 1 1 2 0 0 0 0 0 0 0 0 2 0 1 0 0 0 0 0 0 2 1 0 0 0 0 0 0 0 1 4 0 2 0 0 0 0 0 0 2 0 0 0 0 0 0 0 0 0 1 4 4 2 0 0 1 0 0 0 1 0 0 0 0 0 0 2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 3 0 3 0 0 1 0 0 0 0 1 0 0 9 0 0 0 2 0 0 0 0 0 0 0 0 0 3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 3 7 0 0 0 0 0 0 0 0 0 0 0 0 0 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 Matisia grandifolia Little Otoba gracilipes (A.C.Sm.) A.H.Gentry Pleurothyrium sp. Tetrorchidium andinum Mull.Arg. Trema micranta (l.) Blume Cecropia sciadophylla Mart. Pouroma sp. Nectandra reticulata (Ruíz & Pav.)Mez Grias neubertii J.F. Macbr. Guarea sp. Trichilla pallida Sw. Parkia sp. Otoba sp. Indeterminada Pouteria capacifolia Pilz Trema integérrima (Beurl.) Standl. Coussapoa herthae Midbr. Acalypha sp. Anthurium sp. Bauhinia sp. Cecropia sp2 Crepidosperma sp. Dacryodes peruviana (Loes.) J.F.Macbr. Ficus sp2. Guadua angustifolia Kunth Urticaceae Inga sp2 Lozanella Piper sp2 Piper fuliginosum Sodiro Pouteria sp2 Tapirira guianensis Aubl. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 4 1 1 4 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2 0 0 0 0 7 1 1 1 2 1 1 1 5 1 4 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 6 2 2 9 7 1 1 1 1 3 1 2 1 4 3 El listado de especies (cuadro 1), se ingresó en el programa PAST generándose el índice de diversidad de Simpson (cuadro 2), el cual resulta que los valores son altos en base al número de especies e individuos en cada transecto ya que todos los valores son cercanos a 1, el índice de Simpson indica la relación entre riqueza o número de especies y la abundancia o número de individuos por especies en cualquier sitio dado (Smith 2001); excepto para el caso de unas pocas especies de una comunidad no hay ninguna relación entre la riqueza de una comunidad (número de individuos) y su diversidad. Cuadro 2. Datos de los cinco transectos con datos de abundancia y diversidad Transecto 1 Transecto 2 Transecto 3 Transecto 4 Especies 26.00 27.00 16.00 22.00 Individuos 42.00 45.00 35.00 51.00 Índice de Simpson 0.94 0.95 0.91 0.91 96 Transecto 5 20.00 60.00 0.91 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Los estudios comparativos son escasos en los bosques húmedos tropicales (Campbell et al., 1986), más aun en el litoral ecuatoriano, y no hay muchos estudios disponibles para comparar menos aun con la metodología presentada. El Cuadro 3 indica que los transectos con más afinidad son del T1 al T4 por lo que se supone que pertenecen a bosque maduro (homogéneo) y el T5 que tiene valores menores y diferentes a los otros 4 pertenece a un bosque alterado o en proceso de regeneración y corrobora también la composición de especies que son diferentes que los otros 4 transectos (cuadro 1). De la especie # 64 al 78 del T5 (Cuadro 1) pertenece a bosque alterado y es por esa razón no comparte muchas especies (3 especies) con los otros transectos que pertenecen a bosque maduro. Cuadro 3. Indice de similitud de Bray-Curtis entre los 5 transectos en estudio 0 T1 T2 T3 T4 T5 T1 1 0,32 0,18 0,11 0,03 T2 0,32 1 0,22 0,16 0,07 T3 0,18 0,22 1 0,23 0 T4 0,10 0,16 0,23 1 0,04 T5 0,03 0,06 0 0,04 1 CONCLUSIONES Y RECOMENDACIONES La biodiversidad de los transectos según el índice de diversidad de Simpson es alto (0.91-0.94). Pero al comparar los transectos con las especies y sus frecuencias el transecto 5 es diferente por la composición de especies que el resto de transectos. Se recomienda hacer seguimientos periódicos y a largo plazo porque hace 34 años se realizó el primer estudio por parte de C. H. Dodson & A. H. Gentry haciéndose necesario la instalación de parcelas permanentes para su respectivo seguimiento y evaluación. AGRADECIMIENTOS La presente investigación fue realizada en la cátedra de Dendrología de la Maestría de Manejo y Aprovechamiento Forestal de la UTEQ en el periodo junio- agosto 2012, por la cual agradezco a los estudiantes, al Blgo. Fredy Villao y al Dr. Bryan Maclaren, que ayudaron tanto en la fase de campo como en la obtención de los datos preliminares. BIBLIOGRAFÍA Bray, J. R., & Curtis, J. T. (1957). An ordination southern Wisconsin. Ecological monographs, 27(4), 325-349. of the upland forest communities of Campbell. D.G., D.C. Daly, G.T Prance, U.N. Maciel. 1986. Quantitative ecological inventory of terra firme and várzea tropical forest on the Río Xingu, Brazilian Amazon. Brittonia 38:369-393. Caranqui. J. 2011a. Estructura y composición de un bosque siempreverde montano bajo en Río Negro (Baños. Tungurahua). Tesis digitales ESPOCH. (en línea). Consultado 18Nov. 2012. Disponible en http://dspace.espoch.edu.ec/handle/123456789/509. Caranqui. J. 2011b. Composición y estructura del bosque húmedo tropical en la estación experimental Pastaza. Tesis digitales Dspace ESPOCH. (en línea). Consultado 18 Nov. 2012. Disponible en http://dspace.espoch.edu.ec/handle/123456789/510 Cerón. C.E. 2003. Manual de Botánica ecuatoriana: Sistemática y Métodos de estudio. Facultad de Filosofía. Letras y Educación. Universidad Central del Ecuador. Quito - Ecuador. 97 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Dodson. C.H. and A.H. Gentry. 1987. Contribution of Nontrees to Species Richness of Biotropica 19(2):149-159. Tropical Rain Forest. Dinerstein E., D.M. Olson, D.J. Graham, A.L. Webster, S.A. Primm, M.P. Bookbinder y G. Ledec. 1995. Una evaluación del estado de conservación de las ecoregiones terrestres de América Latina y el Caribe. WWF y Banco Mundial. Washington, DC. USA. Golicher, D s/f. ¿Cómo cuantificar la diversidad de especies?. http://www.dfpd.edu.uy/cerp/cerp_norte/cn/Biologia/BIODIV/Como%20cuantificar%20la%20diversidad,%20algunos%20 ejercicios.pdf Hubell. S.P. and R.B. Foster. 1992. Short-term dynamics of a neotropical forest: whith ecological research matters to tropical conservation and management. Oikos 63:48 Jørgensen, P. M. and S. León-Yánez. 1999. Catalogue of the Vascular Plants of Ecuador. Missouri Botanical Garden. Missouri Botanical Garden. 2013. Base de datos trópicos. (En línea).. Disponible en htpp://www.tropicos.org. Phillips. O. y J. S. Miller. 2002. Global patterns of plant diversity: Alwyn H. Gentry's forest transect data set. Monographs in Systematic Botany from the Missouri Botanical Garden 89:1–319. Sierra. R. 1999. Propuesta preliminar de un sistema de clasificación de vegetación para el Ecuador continental. Quito – Ecuador. Smith L. and T. Smith. 2001. Ecología. Pearson Education. Madrid. Pag. 664. 98 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 CARACTERIZACIÓN DE LAS ÁREAS NATURALES (BOSQUES Y PÁRAMOS) COMO HERRAMIENTA DE ORDENAMIENTO TERRITORIAL EN LA PROVINCIA DE CHIMBORAZO. Wilfrido Haro1, Jorge Caranqui2, Diana Bustamante1, Fabián Salas1. 1Gobierno Autónomo Descentralizado de la Provincia de Chimborazo. (GADPCH) CHEP - Escuela Superior Politécnica de Chimborazo. (ESPOCH) 2Herbario [email protected] Palabras claves: Bosque, ecosistema, cobertura vegetal, ordenamiento territorial, composicion de especies Antecedentes Para concertar la territorialización de la provincia, desde una perspectiva participativa, se planteó en el año 2011 la elaboración de un Plan de Ordenamiento Territorial (PDOT) el cual está en vigencia desde el año 2012, este plan reúne en macrozonas las áreas de influencia que son competencia del Gobierno Provincial, permitiendo advertir la necesidad de disponer de una herramienta que zonifique el territorio más específicamente, de allí nace la propuesta de realizar una zonificación económica – ecológica, que busca identificar áreas homogéneas de administración del territorio a menor escala (parroquias y/o comunidades), que incluyan áreas antrópicas, productivas, seminaturales y áreas de conservación. El presente estudio pretendió afianzar la planificación de una forma más estratégica donde se articulen a todos los actores involucrados, con los beneficios ambientales que brinda principalmente el páramo y el bosque como sistemas de vida, a través de un eficaz ordenamiento territorial provincial. Objetivo Reconocer y valorar con los actores sociales, económicos e institucionales, la importancia de los ecosistemas, paisajes y otros servicios asociados a éstos, para un modelo eficiente de gestión territorial del GAD Provincial de Chimborazo. Criterios de Trabajo 1. Zonificación con articulación basado en el mapa de ecosistemas, cobertura vegetal, uso de suelo y un análisis espacial de condiciones particulares de la provincia. 2. Consideraciones para una estrategia de fomento productivo, centrado en aspectos como clima, riego y factores culturales. 3. Clasificación del uso del suelo, con referencia a los sistemas actuales y sus problemas hoy y en perspectiva. 4. Estrategia social para impulsar cambio de usos del suelo o sistemas de producción, en el marco de la ZEE y PDOT, para lo que se levantarían en la línea base, variables culturales como sistemas de representación o autoridad, capital social o juventud. 5. Zonificación en base a criterios altitudinales para la zona baja, media y alta, considerando los limitantes que plantean las pendientes. 6. Compatibilizar, mediante el debate informado con actores territoriales, la propuesta de zonificación y regulación enfocados en los PDOT’s parroquiales y cantonales. Metodología La primera fase fue la recopilación de información de campo, iniciando con una reunión entre la parte botánica y geográfica, para establecer en base a mapas, imágenes satelitales, etc., los sitios de interés para el estudio botánico. A través de imágenes satelitales (RAPIDEYE 2009-2011) se determinan zonas con particularidades ecológicas diferenciables en las imágenes. Se realizó una supervisión de campo para corroborar los puntos previamente 99 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 señalados. Los mismos, se escogieron definitivamente, a juicio del equipo de trabajo conformado por los técnicos del GADPCH y el Curador del Herbario CHEP, de la Escuela Superior Politécnica de Chimborazo. Es importante mencionar que los sitios seleccionados para el muestreo que estuvieron ubicados en áreas con varios niveles de intervención; se mantuvo una distancia mínima de 200 metros, perpendicular a la línea divisoria de cambio de uso de suelo, para tener una intervención antrópica mínima. Para la evaluación cualitativa y cuantitativa se utilizó dos metodologías: para los bosques, se instaló una área de muestreo de 0.1 hectárea, divididos en 5 transectos de 50 x 4 m que es variante de los transectos realizados de Gentry (Phillips & Miller, 2002; Cerón, 2003), ya que estos eran de 10 juegos de transectos de 50 x 2m. Utilizando transectos amplios se reduce los efectos de micro hábitats en los resultados generales, y tomando en cuenta la fragmentación del bosque que en la actualidad ocurre; los transectos de 50 x 4m nos ayudan a estandarizar el área de muestreo. Los siete transectos son del igual tamaño, permitiendo una comparación entre ellos (Caranqui, 2013). Se tomó el DAP (diámetro a la altura del pecho) de las especies mayores a 5 cm para bosques montanos y 10 cm para bosques premontanos. Se colectaron especímenes botánicos de la mayoría de los individuos marcados (incluyendo todas las especies no identificadas en el campo), un duplicado para muestras infértiles y tres para muestras fértiles (Caranqui, 2013), esto con el objetivo de realizar una posterior identificación a nivel de especie. Para los páramos la metodología utilizada fue la propuesta por Pauli et al. (2003), para el proyecto GLORIA de la región europea, adaptada a los páramos andinos (Eguiguren, 2010). En cada cima se instaló cuadrantes de 5×5 m, que se usaron para el muestreo de la vegetación. Cada cuadrante se subdividió en parcelas de 1 x 1 m., las observaciones de vegetación se llevó a cabo únicamente en las cuatro parcelas de las esquinas o extremos, ya que los otros pueden quedar alterados por el pisoteo de los investigadores a lo largo del muestreo (Caranqui et al., 2013). De las 4 parcelas se escogieron totalmente al azar 1 unidad muestreal, es decir se muestreo 1 m² por zona de muestreo. En cada uno de las parcelas de 1x1 m., se subdividieron en cuadriculas de0.1 x 0.1m., para ello se utilizó un armazón de tubería PVC con un enrejado formado por hilos finos que delimitan 100 celdillas de 0,1 x 0,1 m., de acuerdo a la metodología del manual GLORIA adaptado por Rodríguez, (2011), con el fin de obtener la mayor cantidad de información para su posterior análisis (Caranqui et al., 2013). Dentro de las parcelas se levantó información referente al número de especies y la cobertura de cada una de ellas, esto sirvió para determinar la diversidad por familia, densidad, diversidad alfa y beta. Se colectaron especímenes botánicos de la mayoría de los individuos (incluyendo todas las especies no identificados en el campo) un duplicado para muestras infértiles y tres para muestras fértiles (Caranqui et al., 2013). Para la identificación de las especies vegetales se la realizó mediante una comparación en seco con: especímenes herborizados del Herbario de la Escuela Superior Politécnica del Chimborazo (CHEP), claves dicotómicas, bases de datos en línea del New York Botanical Garden (NY: http://www.nybg.org/), el Catálogo de Plantas Vasculares (Jørgensen & León-Yánez, 1999) y la actualización se consultó en la base de datos Trópicos (www.tropicos.org) del Missouri Botanical Garden. El trabajo de identificación taxonómica fue realizado en su totalidad por el equipo de trabajo del Herbario de la Escuela Superior Politécnica del Chimborazo (CHEP). Para la caracterización de las zonas investigadas, se utilizó como base el Catálogo de Objetos Nacional, documento donde se describe las diferentes coberturas vegetales y su diversidad florística; se utilizaron métodos comparativos para la clasificación de ecosistemas, principalmente con las muestras obtenidas en cada sitio, se ordenaron según las especies registradas en cada uno de los ecosistemas que presenta el Sistema de Clasificación de los Ecosistemas del Ecuador Continental, MAE (2013). En la ordenación no se suponen discontinuidades entre los hábitats y éstos se agrupan en series ecológicas o sistemas de coordenadas para reducir patrones complejos a formas simples e interpretables (Moreno, 2001); estos análisis se realizaron con datos de especies cualitativos (de presencia – ausencia) determinando a que ecosistema pertenece cada zona muestreada. Para la zonificación económica y ecológica, se optó por utilizar el mapa de ecosistemas de la provincia como insumo principal junto con las herramientas antes mencionadas en los criterios de trabajo, dando importancia a tres áreas particulares biofísica-ambiental, socio económica y cultural, de allí se aplicó el cruce de diversas variables entre las cuales se destacan: ecosistemas y uso de suelo, NBI, capital social, riego, pendientes, altitud y ombrotérmico, cada una 100 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 de ellas respondiendo a la necesidad de obtener áreas lo más uniformemente posibles; de allí se priorizó cada una de las áreas para encontrar un equilibrio entre lo productivo y lo conservacional; en las áreas prioritarias se podrán planear los diferentes proyectos tanto agropecuarios como ambientales (dependiendo de su valoración y priorización); estas áreas se han trabajado hasta llegar a un nivel comunitario, siendo una herramienta muy específica de planificación rural. Resultados La diversidad encontrada en los bosques estudiados son de índice bajo a medio según el número de especies inventariadas, principalmente se debe a las actividades antropogénicas realizadas a través del tiempo, el bosque como ecosistema se encuentra fragmentado en parches de 20 has en promedio, manifestando una marcada heterogeneidad, determinándolos referentemente como únicos en un sentido particular de dominancia de las especies y su diversidad, de allí la trascendencia de proteger con fines de conservación el máximo número de fragmentos. Con la finalidad de utilizar esta información como una herramienta oficial y homogenizar resultados, en relación a descripciones y nomenclatura de los ecosistemas de Chimborazo, se utilizó lo establecido en el Sistema de Clasificación de los Ecosistemas del Ecuador Continental, MAE (2013), pues esta clasificación ha unido las experiencias de clasificaciones anteriores, generando mayor detalle de la información florística El sistema de clasificación generó 17 ecosistemas dentro del territorio provincial. (Tabla N° 1) El territorio de la Provincia de Chimborazo tiene, un área aproximada de 650.062,12 Has, mantiene una cobertura vegetal natural del 37,9% (246.421,67 has) entre arbustales, bosques y herbazales. Dentro de esta cobertura el Ecosistema de Páramo (herbazales) resulta además de ser el más sobresaliente, el que ocupa la mayor superficie con 169.030,01 ha., representa el 26%. Los restantes ecosistemas representan menos del 4% cada uno; sumados todos, ocupan un porcentaje menor a la cobertura de páramo, esto es el 10.57%, contrastada con la zona antrópica 60,95% que corresponde a las áreas intervenidas (396.195,73 has). Gráfico 1. Territorio de la Provincia Fuente: GADPCH (2013) Estos datos nos han ayudado a trabajar en una propuesta de zonificación económica y ecológica, dentro de la cual se han detectado principalmente tres macrozonas: uso de conservación, uso de producción y uso especial; estas tres responden al enfoque que cada área debe tener para su mejor desarrollo y uso. La macrozona de conservación, objeto de nuestro estudio, se la ha subdividido en cuatro subzonas, respondiendo a la priorización realizada en talleres participativos con la gente y desde un criterio técnico del equipo de trabajo del Gobierno Provincial, las cuales son: Zona de importancia ecológica para protección, recuperación y manejo, Zona bajo acuerdo de conservación, Zona de Áreas Protegidas, y Zonas de importancia hídrica. 101 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Tabla 1. Sistema de Clasificación de Sitios Visitados. Sitios Cantón Parroquia Clasificación Ecológica del MAE Código Igualata Guano San Isidro Herbazal del Páramo HsSn02 Ganquis Cuiquiloma Riobamba San Juan Herbazal del Páramo HsSn02 Cóndor Samana Guano San Andrés Herbazal ultrahúmedo subnival del Páramo HsNn02 Ruta del Hielero Guano San Andrés Herbazal y arbustal siempre verde subnival del Páramo Hs Nn03 El lirio Colta Columbe Herbazal del Páramo HsSn02 El Puyal Colta Sicalpa Herbazal del Páramo HsSn02 Maguazo Riobamba Pungala Herbazal del Páramo HsSn02 Tuilcha Penipe Candelaria Arbustal siempre verde y Herbazal del Páramo As Sn 01 Cubillin Chambo Chambo Herbazal del Páramo HsSn02 Pulucorral Guargualla Guamote Cebadas Herbazal del Páramo HsSn02 Pomacocho Alausí Achupallas Herbazal del Páramo HsSn02 Juval Alausí Achupallas Herbazal del Páramo HsSn02 Tambo Palitahua 1 Penipe El Altar Bosque siempreverde Montano alto del Norte de la Cordillera Oriental de los Andes. BsAn01 Tambo Palitahua 2 Penipe El Altar Bosque siempreverde Montano alto del Norte de la Cordillera Oriental de los Andes. BsAn01 San Francisco Chambo Matriz Bosque siempreverde Montano alto del Norte de la Cordillera Oriental de los Andes. BsAn01 Llucud Chambo Matriz Bosque siempreverde Montano alto del Norte de la Cordillera Oriental de los Andes. BsAn01 Bacun Chunchi Matriz Bosque siempreverde Montano alto del Norte de la Cordillera Occidental de los Andes. BsAn03 Pollongo Colta J. de Velasco Bosque siempreverde Montano alto del Norte de la Cordillera Occidental de los Andes. BsAn03 Yunguilla Guangras Alausí Achupallas Bosque siempreverde Montano alto del Sur de la Cordillera Oriental de los Andes. BsAn02 102 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Chilincay Manuelita Cumandá Matriz Bosque siempreverde piemontano de la Cordillera Occidental de los Andes. BsPn01 El corazón - Jaluví Pallatanga Matriz Bosque siempreverde montano de la Cordillera Occidental de los Andes. BsAn03 Palitahua Penipe Altar Arbustal siempreverde montano del Norte de los Andes. AsMn01 Guagal Cumandá Matriz Bosque siempreverde montano bajo de la Cordillera Occidental de los Andes. BsBn04 El Carmen Alausí Multitud Bosque siempreverde montano de la Cordillera Occidental de los Andes. BsAn03 Bilbao Penipe Bilbao Bosque siempre verde montano del Norte de la Cordillera Oriental de los Andes BsMn01 Cuenca alta Mazar Alausí Achupallas Bosque siempre verde montano del sur de la Cordillera Oriental de los Andes BsMn 02 Puyal- H Colta Sicalpa Herbazal húmedo montano alto superior de páramo HsSn03 Las Abras boca toma Guano San Andrés Herbazal húmedo montano alto superior de páramo HsSn03 Pasguazo Riobamba San Juan Herbazal húmedo montano alto superior de páramo HsSn03 El arenal RFCH Riobamba San Juan Herbazal húmedo Subnival de páramo HsMn01 Laguna de Colta Colta Santiago de Quito Herbazal inundable de páramo HsSn04 Asaco Guano La Matriz Herbazal y arbustal seco* HsAs00* * Se ha considerado este ecosistema, aunque no se encuentra dentro del Sistema de Clasificación propuesto por el MAE. 103 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Estas zonas nos han ayudado a definir, propuestas y proyectos que fueron encaminados con las categorías del uso de cada zona, estan categorías estan englobadas en: de uso recomendado, no recomendado y de uso compatible; de este modo se han definido las actividades encaminadas a recuperar, proteger y manejar las áreas ambientales con las siguientes propuestas: Plan de reforestación esta orientado en dos líneas de acción diferentes: la primera que es estimular a los productores agrícolas en el uso combinado de sistemas productivos, específicamente la silvopastura, agroforestería, agrosilvopastoriles, barreras vivas, entre otras; de tal manera que se mantenga la biodiversidad, y exista manejo de tierras en áreas que tienen problemas de erosión o degradación. La segunda alternativa será la reforestación a gran escala en áreas que pueden ser tranformadas en plantaciones forestales con fines de conservación, estas áreas deberán tener un plan de manejo sustentable por cada población beneficiaria en este proyecto. Como alternativas se han identificado dos propuestas: promoción de proyectos a través de los denominados mecanismos de desarrollo limpio (MDL), de Implementación Conjunta (IC) y de Actividades Implementadas Conjuntamente (AIC), como venta de servicios ambientales, estos mecanismos podrían considerarse como de incentivos económicos, considerando la imperiosa necesidad de un entorno limpio y equilibrado para asegurar que la provincia tenga un desarrollo sustentable. Además de identificar zonas para la creación de corredores biológicos que permitan un flujo o movimiento de los individuos, de fauna silvestre y de semillas de un fragmento a otro, garantizando así la regeneracion natural, la repoblación de fauna silvestre, el cause normal de cursos hídricos, entre otros. Estas alternativas a corto, mediano y largo plazo, deben ser consideradas como un proyecto de interés provincial después de su aprobación dentro del presupuesto participativo. BIBLIOGRAFÍA Austin, M.P., 2002. Spatial prediction of species distribution: An interface between ecological theory and statistical modeling. Ecol Model 157, 101-118 Caranqui, Jorge et al. 2013. Diversidad Y Similitud Arbórea De Los Bosques Montanos Altos De La Provincia De Chimborazo. En Herbario Escuela Superior Politécnica del Chimborazo - CHEP. Riobamba. Caranqui, Jorge. 2013. Diversidad Y Similitud De Los Páramos De La Provincia De Chimborazo En Ecuador. En Herbario Escuela Superior Politécnica del Chimborazo - CHEP. Riobamba. Ecuador. Instituto Nacional de Estadística y Censos “INEC” – Estadísticas Agropecuarias “ESPAC”, Uso del suelo por categorías de Chimborazo 2011. Ecuador. Ministerio del Ambiente del Ecuador. 2013. Metodología para la Representación Cartográfica de los Ecosistemas del Ecuador Continental. Subsecretaría de Patrimonio Natural. Quito. Ecuador. Ministerio del Ambiente del Ecuador. 2013. Sistema de Clasificación de los Ecosistemas del Ecuador Continental. Subsecretaría de Patrimonio Natural. Quito. Eguiguren, P., Ojeda T., y Aguirre, N. A. (2010). Diversidad florística del ecosistema paramo del Parque Nacional Podocarpus para el monitoreo del cambio climático. Disponible en: Freire-Fierro, A., 2004. Botánica Sistemática Ecuatoriana. Missouri Botanical Garden, St. Louis, Missouri. 104 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Gentry, A.H. 1995. Patterns of diversity and floristic composition in neotropicalmontane forest. Pp. 103-126. In: Churchill, S.P., H. Balslev, Forero,E., &LuteynJ.L. (eds), Biodiversity and conservation of neotropicalsmontane forests. The New York Botanical Garden, Bronx http://www.unl.edu.ec/miccambio/wp-content/uploads/2010/07/Eguiguren-Ojeda-2010_DiversidadFlor%C3%ADstica-del-PNP_docx.pdf IDEAM. 2010. Leyenda Nacional de Coberturas de la Tierra. Metodología CORINE Land Cover adaptada para Colombia Escala 1:100.000. Instituto de Hidrología. Meteorología y Estudios Ambientales: Bogotá, D.C. p. 72. León-Yánez, S. 2000. La flora de los páramos ecuatorianos. En: La biodiversidad de los páramos. Serie Páramo 7: 5-21. GTP/Abya Yala. Quito. Moreno, Claudia, 2001. Métodos para medir la biodiversidad. M&T-Manuales y Tesis SEA, Zaragoza. España vol. 1. Zaragoza, 84 pp Pauli, Harald. et al. 2003. Manual para el trabajo de campo del proyecto GLORIA. Instituto de ecología y conservación biológica. Universidad de Viena. Disponible en: http://www.gloria.ac.at/downloads/GLORIA_MS4_Web_espanol.pdf (Consultado julio 10 del 2013) Sayre, R., et al., A New Map of Standardized Terrestrial Ecosystems of the Conterminous United States, in Professional Paper 1768. 2009, U.S. Geological Survey p. 17. Sierra, Rodrigo. 1999. Propuesta preliminar de un sistema de clasificación de vegetación para el Ecuador continental. Quito – Ecuador. Trópicos base de datos. Missouri Botanical Garden. [Consulta de internet 01 jul. 2013] http://www.tropicos.org 105 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 CONSERVACIÓN Y USO SOSTENIBLE DE RECURSOS GENÉTICOS FORESTALES Andrea Pinos PATRONES ESPACIALES DE RECLUTAMIENTO Y VARIABILIDAD GENÉTICA DE LA PALMA Oenocarpus bataua Mart., EN EL PARQUE NACIONAL YASUNÍ (PNY) Andrés Recalde Redrován DIVERSIDAD GENÉTICA DEL MORETE (Mauritia flexuosa L. f.) EN PANTANOS DE LA REGIÓN SEPTENTRIONAL DEL PARQUE NACIONAL YASUNÍ. Rommel Montúfar PRIORIDADES DE INVESTIGACIÓN GENÉTICA Y ECOLÓGICA PARA LA CONSERVACIÓN DE ESPECIES Y BOSQUES Y EL DESARROLLO FORESTAL EN ECUADOR Sebastián Escobar ESTRUCTURA GENÉTICA DE LAS POBLACIONES DE Oenocarpus bataua (ungurahua): UNA HERRAMIENTA PARA LA CONSERVACIÓN DE UN RECURSO FORESTAL EN EL NETRÓPICO María Mercedes Pavón EVALUACIÓN DEL ESTADO DE CONSERVACIÓN Y CARACTERIZACIÓN POBLACIONAL DE DOS ESPECIES FORESTALES PRIORIZADAS EN LA MICRO-CUENCA DEL RIO CHIMBORAZO Juan Chucuri COLECTA Y CARACTERIZACIÓN MORFOLÓGICA IN SITU DE CAPULÍ (Prunus serotina Ehrh) DEL BANCO NACIONAL DE GERMOPLASMA DEL INIAP-ECUADOR 106 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 PATRONES ESPACIALES DE RECLUTAMIENTO Y VARIABILIDAD GENÉTICA DE LA PALMA Oenocarpus bataua Mart., EN EL PARQUE NACIONAL YASUNÍ (PNY) Andrea Pinos, Rommel Montúfar Pontificia Universidad Católica del Ecuador (PUCE), Lab. Ecología y Genética (113), Av. 12 de Octubre y Av. Patria, Quito, Ecuador [email protected] Introducción El funcionamiento de los sistemas forestales está regulado por fuerzas evolutivas como la selección natural, endogamia y deriva génica; y por procesos ecológicos e históricos. Estos procesos influyen directamente en la estructura genética de las poblaciones de plantas silvestres como en (i) la habilidad reproductiva de la especie, la cual está determinada por las tasas de dispersión de semillas (alelos maternales y paternales) y polen (alelos paternales; Karubian et al. 2010) y (ii) las tasas de flujo génico entre poblaciones, las cuales permiten inferir las consecuencias ecológicas de la dispersión y polinización sobre la estructura genética de la especie (Loveless y Hamrick 1984). El flujo génico entre poblaciones puede realizarse a cortas o largas distancias dependiendo de la presencia de barreras que lo limiten. La dispersión a corta distancia induce el apareamiento de individuos geográficamente próximos lo que provoca una estructuración de la diversidad genética a nivel local, y que favorece la divergencia de las poblaciones. Por otro lado, eventos de dispersión a media y larga distancia, permiten niveles altos de flujo génico entre poblaciones lo que incrementa su base genética, reduce la endogamia y mantiene poblaciones biológicamente viables y genéticamente homogéneas (Lowe et al. 2005). Por lo tanto, el caracterizar la cantidad y diversidad genética asociada al movimiento de polen y semillas (propágulos) es fundamental para comprender la dinámica de las poblaciones actuales y como estas intercambian su material genético. Particularmente en palmas (Arecaceae), la dispersión es un proceso fundamental para el reclutamiento (colonización y establecimiento de nuevos individuos) y su regeneración dentro de un bosque. Los propágulos de palmas no dispersados sufren altas tasas de mortalidad cerca de los adultos (mortalidad dependiente de la distancia), mientras que los propágulos dispersados a largas distancias presentan mayores tasas de supervivencia (Swamy y Terborgh 2010). Los efectos de la dispersión de semillas han sido analizados con modelos de diversidad, los cuales explican cómo una alta diversidad de especies es mantenida dentro de los bosques tropicales (Loveless y Hamrick, 1984). El modelo más estudiado de estos es el de Janzen y Connell (Janzen 1970; Connell, 1971). Esta hipótesis sugiere que el reclutamiento de plántulas cerca de los padres puede reducirse como resultado de la densidad dependiente de la mortalidad, debido a depredadores (herbívoros) y patógenos (Clark y Clark 1984). Estos usualmente concentran sus actividades cerca de individuos adultos, que es donde las semillas caen y germinan con mayor frecuencia. Al existir una baja tasa de reclutamiento de las plántulas, la probabilidad de establecimiento de otras especies en estos sitios aumenta reduciendo el potencial de dominio de una sola especie en una comunidad. En consecuencia, las bajas tasas de reclutamiento cerca de parentales permiten mantener la alta diversidad de especies en bosques tropicales (Clark y Clark 1984). 107 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 El conocer los efectos de la dispersión y reclutamiento nos permite entender los mecanismos de flujo génico en las poblaciones. Estos mecanismos y en particular la dispersión, dan pautas para desarrollar estrategias dirigidas a preservar los procesos que favorecen el flujo de genes y la resilencia de las poblaciones a la fragmentación del paisaje u otros cambios abióticos (Dick et al., 2003). Esto ayuda al manejo y conservación de poblaciones vegetales y determina la capacidad que tienen las plantas para dispersarse en un hábitat heterogéneo y dinámico. Mediante estudios de paternidad, con el uso de marcadores moleculares, es posible evaluar tasas de flujo génico entre poblaciones (Gaiotto et al. 2003; Karubian et al. 2010). Estos estudios permiten determinar el parentesco de las plántulas establecidas con sus potenciales padres; por lo tanto, nos permiten estimar las distancias promedio a las cuales las plántulas se dispersan de sus padres. Oenocarpus bataua es una palmera arborescente, monoica, alógama que se distribuye ampliamente en las regiones tropicales y subtropicales de Sudamérica, incluyendo la isla de Trinidad en el Caribe (Montúfar y Pintaud 2008). O. bataua es la séptima especie vegetal más abundante dentro de los bosques amazónicos, registrándose hasta 108 individuos/ha (Steege et al. 2013). Esta especie se caracteriza por sus múltiples usos por parte de los pueblos amerindios (Balick ,1979). Debido a su abundancia y alta productividad de frutos ricos en aceites comestibles, O. bataua es un recurso alimenticio importante tanto para comunidades humanas como para animales frugívoros. A pesar de su importancia ecológica y su potencial económico como nueva fuente oleaginosa la dinámica de la diversidad genética de O. bataua no ha sido explorada. El presente estudio aporta sobre los patrones de dispersión, estructura y diversidad genética, mediante estudios de paternidad los cuales nos permite esclarecer aspectos claves sobre la ecología de la dispersión de este recurso forestal. Objetivo general Estudiar la dinámica, reclutamiento y variabilidad genética entre cohortes (adultos y juveniles) de la especie de palma Oenocarpus bataua en un bosque tropical. Metodología Área de estudio y diseño de muestreo El estudio se realizó en el Parque Nacional Yasuní (PNY) en la parcela de 50 ha establecida por el del Proyecto Dinámica de Bosque de Yasuní (PDBY) de la Pontificia Universidad Católica del Ecuador. Se seleccionó 25 ha contiguas de las 50 ha que comprende la parcela. La selección de las 25 ha se realizó tomando en cuenta las condiciones topográficas del terreno, incluyendo las zonas de colinas en donde la abundancia de la especie es mayor. Las 25 ha elegidas se encuentran divididas en 625 cuadrantes de 20 x 20m. Para el muestreo de plántulas se seleccionó al azar 200 cuadrantes de 20 x 20m de los 625. En cada uno de los 200 cuadrantes se colectó tejido vegetal (1cm x 2cm) de una plántula. En total se colectaron 199 plántulas (una plántula fue descartada). La base de datos de PDBY proporcionó la ubicación exacta de 44 palmeras adultas (tronco visible y presencia de estructuras reproductivas) registradas dentro de las 25 ha estudiadas. Se colectaron muestras de tejido foliar o radicular de todos los individuos adultos. Se incluyó en el diseño de colecta adultos reproductivos cercanos a la parcela de 25 ha. Para esto se trazó tres transectos perpendiculares de 200m x 50m para cada 108 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 uno de los lados de la parcela de 25 ha (norte, sur, este y oeste). La distancia entre los transectos fue de 100 m y se obtuvo un total de 16 individuos adultos adicionales muestreados en los alrededores del área de estudio (Figura 1), los cuales se utilizaron para realizar el análisis de parentesco junto con los individuos adultos colectados dentro de las 25 ha. Se realizó extracciones manuales para aislar el ADN siguiendo el protocolo modificado de Doyle y Doyle (1987). El protocolo que se utilizó para la reacción en cadena de la polimerasa fue el propuesto por Montúfar et al. (2006) para cada uno de los 6 loci de microsatélites utilizados en este estudio. Los productos de PCR fueron analizados mediante geles denaturantes de poliacrilamida para obtener los patrones alélicos de los microsatélites. Análisis de datos Una vez generadas las matrices para plántulas (n=199) y adultos (n=60) se realizó los siguientes análisis estadísticos: estructuración poblacional (FST), heterocigocidad observada y esperada (Ho, He), equilibrio Hardy-Weinberg (H-W) e índice de endogamia (FIS) con los programas estadísticos GenePop 4.0.10 (Raymond y Rousset 1995) y GenAlEx 6.0 (Peakall y Smouse 2005). El porcentaje de loci polimórficos, número promedio de alelos por locus, diferenciación genética (Dest; Jost, 2008) y análisis de autocorrelación espacial (test de Mantel) con los programas estadísticos GenAlEx 6.0, Arlequin 3.11 (Excoffier et al. 2005) y SMOGD 1.2.5 (Crawford, 2010). Para el análisis de paternidad se siguió la guía descrita en Jones et al. (2010) con la ayuda del programa Cervus 3.5 (Marshal et al. 1998). Resultados y discusión En la población de adultos de la parcela (n=44) cuatro loci presentaron desviaciones estadísticamente significativas a lo esperado bajo el equilibrio H-W; mientras que, en plántulas (n=199) los seis loci se desviaron significativamente del equilibrio H-W. La población de adultos como la de plántulas presentaron altos valores de diversidad génica (Hepromedio= 0,87 para plántulas, Hepromedio= 0,86 para adultos). Los valores de diversidad obtenidos son superiores a estudios similares en poblaciones de O. bataua provenientes de la amazonía peruana en donde la He varió entre 0,66 a 0,77 (He= 0,66, 0,73, 0,77; Montúfar, 2007). La escasa diferencia en diversidad (He) obtenida a favor de las plántulas es más evidente en base al número de alelos. El Napromedio en plántulas fue de 18, mientras que en adultos llegó a 13,6; si bien el Na es altamente dependiente del área de estudio, los resultados obtenidos provienen de áreas muestreales relativamente similares y son comparables. Adicionalmente, las poblaciones de plántulas presentaron un valor promedio de 4,3 alelos únicos, mientras que las poblaciones de adultos no presentaron alelos únicos. Los valores del índice de endogamia indican que, las poblaciones de plántulas (FIS promedio = 0,280) a pesar de ser más diversas son más endogámicas que las poblaciones de adultos (FIS(promedio)= 0,175; Tabla 1, Tabla 2). El aparente antagonismo entre más diversas y endogámicas podría estar explicado por la naturaleza de las poblaciones de plántulas las cuales al no estar sometidas a un proceso de selección natural tienden a presentar un mayor número de alelos durante sus primeros estadíos que las poblaciones de adultos, las cuales son producto de procesos de selección (Eguiarte et al., 1992; (Primack y Kang, 1989; Lenormand, 2002). Adicionalmente, la alta diversidad en plántulas también está relacionada a procesos de dispersión a larga distancia los cuales favorecen el ingreso de semillas provenientes de poblaciones diferentes. En particular, se ha evidenciado la existencia de mecanismos de dispersión a larga distancia en O. bataua a 109 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 través de aves como los guácharos (Steatornis caripensis), quienes tienen la capacidad de movilizarse a distancias mayores a 70 km (Montúfar et al. 2011). Una mayor endogamia en plántulas que en adultos, propone dos escenarios en la dinámica del bosque: (i) Los mecanismos de flujo génico actuales que estructuran a las poblaciones de plántulas son menos eficaces que los mecanismos de flujo génico que formaron las poblaciones de adultos hace más de 50 años en el pasado. Teóricamente una palmera adulta y reproductiva de O. bataua dentro del bosque requiere de al menos 50-80 años para desarrollarse (Miller, 2002). Por lo tanto, un índice menor de endogamia en adultos puede estar relacionado a una mayor abundancia de dispersores en el bosque y consecuentemente a mejores condiciones de conservación del bosque en el pasado. (ii) Al igual que la diversidad genética, las poblaciones de plántulas al no estar totalmente reguladas por la selección natural pueden presentar una mayor tasa de individuos homocigóticos, los cuales posteriormente serán eliminados via selcción natural. Los análisis de asignamiento de paternidad proveen dos valores que representan la distancia hacia ambos padres potenciales, generalmente uno de ellos encontrándose cerca de la plántula y el otro más distante. Debido a, que la distancia de dispersión de los propágulos hacia la madre va a ser menor que la distancia hacia el padre, nosotros asumimos que la distancia más corta representa la distancia hacia la madre, y la distancia más larga hacia el padre. En base a este diseño, el rango de la distancia de dispersión hacia las potenciales madres fue de 8 – 570 m, con una media de 277 m y σ= 155m, mientras que el rango de distancia hacia el potencial padre fue de 31 a 850m, con una media de 345 m y σ= 116m. Dentro del contexto de la hipótesis de Janzen y Connel (año), se asume una correlación significativa entre distancia de dispersión y número de plántulas; sin embargo estos valores no fueron obtenidos en este estudio. La dispersión de plántulas en función de las madres no presentó una correlación significativa entre distancia y número de individuos; pero sí se obtuvo una correlación positiva y significativa entre distancia hacia el padre y número de individuos. Por consiguiente en nuestro diseño experimental no se pudo detectar el modelo propuesto por Janzen y Connel. Dos aspectos adicionales inferidos de los resultados de este estudio son: (i) Un alto porcentaje de plántulas fueron asignadas a pocos potenciales parentales. En particular, dos individuos fuera de la parcela fueron asignados como potenciales padres de un alto número de plántulas (qué número o porcentaje). (ii) La estructura genética observada en las 25 ha (con que valores se hizo a matriz genética, Fst???) no fue correlacionada (valores r= y probabilidades) a la matriz espacial (X, Y). Esto sugiere que la estructura genética de los individuos a escala de micro hábitat no está controlada por factores abioticos. Sin embargo, esta variación topográfica no tuvo ninguna influencia en los patrones de diversidad dentro de la parcela estudiada (r = 0.019, p = 0.085). Los resultados obtenidos del presente estudio proveen información sobre el funcionamiento de las poblaciones de O. bataua. Programas de conservación y manejo de poblaciones silvestres deben tomar en cuenta los mecanismos genéticos que controlan las poblaciones a escalas locales. Agradecimientos Al. Dr. Renato Valencia por facilitarnos información y el acceso a la parcela del proyecto Damica del Bosques de Yasuni. 110 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Literatura citada Balick, M. 1979. Amazonian Oil Palms of Promise: a Survey. Economic Botany. 33(1):11–28. Clark, DA; Clark, DB; Sandoval, MR; Castro, CM. 1995. Edaphic and human effects on landscape-scala distributions of tropical rainforest palms. Ecology. 76 (8): 2581–2594. Clark, DA; Clark, DB. 1984. Spacing dynamics of a tropical rain forest tree: evaluation of the Janzen-Connell model. American Naturalist. 124(6): 769–788. Connell, JH. 1971. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. In: Boer PJ den, Gradwell GR (eds) Dynamics of Populations. Centre for Agricultural Publishing and Documentation, Wageningen, pp 298-312 Dick, CW; Etchelecu, G; Austerlitz, F. 2003. Pollen dispersal of tropical trees (Dinizia excelsa: Fabaceae) by native insects and African honeybees in pristine and fragmented Amazonian rainforest. Molecular Ecology. 12: 753–764. Eguiarte, LE; Perez-Nasser, N; Pijero, D. 1992. Genetic structure, outcrossing rate and heterosis in Astrocaryum mexicanum (tropical palm): implications for evolution and conservation. Heredity. 69: 217–228. Excoffier L; Laval G; Schneider S. 2005. ARLEQUIN version 3.0: an integrated software package for population genetics data analysis. Evolutionary Bioinformatics Online. 47–50. Gaiotto, F; Grattapaglia, D; Vencovsky, R. 2003. Genetic Structure, Mating System, and Long-Distance Gene Flow in Heart of Palm (Euterpe edulis) Mart. Journal of Heredity. 94: 399–406. Ghazoul, J; McLeish, M. 2001. Reproductive ecology of tropical forest trees in logged and fragmented habitats in Thailand and Costa Rica. Plant Ecology. 153: 335–345. Janzen, DH. 1970. Herbivores and the number of tree species in tropical forests. American Naturalist. 104:501–528. Jones, A; Clayton, M; Small, K; Paczolt, A; Ratterman, N. 2010. A practical guide to methods of parentage analysis. Molecular Ecology Resources. 10: 6–30. Karubian, J; Sork, VL; Roorda, TR; Duraes y Smith, TB. 2010. Destination-based seed dispersal homogenizes genetic structure of a tropical palm. Molecular Ecology. 19: 1745–1753. Laurance, WF; Gascon, C; Rankin-de Merona, J. 1999. Predicting effects of habitat destruction on plant communities: a test of a model using Amazonian trees. Ecology Applied. 9: 548–554. Lenormand, T. 2002. Gene flow and the limits on the natural selection. Trends in Ecology and Evolution. 17: 183–189. 111 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Loveless, MD; Hamrick. JL. 1984. Ecological Determinants of Genetic Structure in Plant Populations. Annual Review of Ecology and Systematics. 15: 65–95. Lowe, AJ; Boshier, D; Ward, D; Bales, CFE; Navarro, C. 2005. Genetic resource impacts of habitat loss and degradation; reconciling empirical evidence and predicted theory for neotropical trees. Heredity. 95:255–273. Marshall, TC; Slate, J; Kruuk, L; Pemberton, JM. 1998. Statistical confidence for likelihood-based paternity inference in natural populations. Molecular Ecology. 7: 639–655. Miller, C. 2002. Fruit production of the ungurahua palm (Oenocarpus bataua subsp. bataua, Arecaceae) in an indigenous managed reserve. Economic Botany. 56: 165–176. Montúfar, R; Maria, C; Pham, JL; Pintaud, JC. 2006. Isolation of 23 polymorphic microsatellite loci in the Neotropical palm Oenocarpus bataua Martius (Arecaceae). Moleculary Ecology Notes. 7 : 75–78. Montúfar, R; Pintaud, JC. 2008. Estatus taxonómico de Oenocarpus bataua (Euterpeae, Arecaceae) inferido por secuencias del ADN cloroplástico. Revista Peruana de Biología. 15: 73–78. Montúfar, R. 2007, Structure génétique, biochimique, morphologique et écologique de Oenocarpus bataua Mart. (Arecaceae): perspectives pour la valorisation durable d' une ressource foretiére néotropicale. Tesis de doctorado. École Nationale Supérieure Agronomique de Montpellier. Montúfar, R; Anthelme, F; Pintaud, J-C; Balslev H. 2011. Disturbance and Resilience in Tropical American Palm Populations and Communities. Botanical Review. 77:426–461. Peakall, R; Smouse, PE. 2012. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research – an update. Bioinformatics. 28: 2537–2539. Primack, R. y Kang, H. 1989. Measuring fitness and natural selection in wild plant populations. Annual Review of Ecology. Systematics. 20: 367–96. Raymond, M; Rousset, F. 1995. GENEPOP (version 1.2): population genetics software for exact tests and ecumenicism. Journal of Heredity. 86: 248–249. Svenning, JC. 2001. On the role of micro environmental heterogeneity in the ecology and diversification of Neotropical rainforest palms (Arecaceae). Botanical Review. 67: 1–53. Swamy, V; Terborgh, JW. 2010. Distance-responsive natural enemies strongly influence seedling establishment patterns of multiple species in an Amazonian rain forest. Journal of Ecology. 98: 1096–1107. Steege, H ter; Pitman, NC; ... ; Silman, N. 2013. Hyperdominance in the Amazonian Tree Flora. Science. 342(6156):1243092. 112 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figuras y tablas Fig 1. Distribución de plántulas (n=) y adultos (n=) de Oenocarpus bataua en el área de studio (parcela Dinámica del Bosque tropical de Yasuni). Tabla 1. Valores del índice de endogamia (FIS) presentes en la población de plántulas (n = 199) analizada con 6 loci de microsatélites. Abreviatura: PP = plántulas Pop PP Locus Ob22 Ob11 Ob01 Ob04 Ob21 Ob23 FIS 0,408*** 0,384*** 0,354*** 0,184** 0,074 0,276*** Media de 0,280** plántulas Tabla 2. Valores del índice de endogamia (FIS) presentes en la población de adultos (n= 44) analizada con 6 loci de microsatélites. Símbolos= ** p<0,01, *** p<0,001, NS= p>0,05 Adultos Adultos dentro de la parcela Locus Ob22 Ob11 Ob01 Ob04 Ob21 Ob23 Media adultos 113 FIS 0,291*** 0,130** 0,09NS 0,115** 0,268*** 0,160** 0,175** PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 DIVERSIDAD GENÉTICA DEL MORETE (Mauritia flexuosa L. f.) EN PANTANOS DE LA REGIÓN SEPTENTRIONAL DEL PARQUE NACIONAL YASUNÍ Andrés Recalde Redrován, Rommel Montúfar G. Lab. 113 Ecología y Genética, Pontificia Universidad Católica del Ecuador. Av. 12 de Octubre 1076 y Roca. [email protected] Introducción Los pantanos son hábitats que albergan una diversa y especializada flora y cumplen un rol importante en la dinámica forestal del bosque amazónico (Horn et al. 2012; Endress et al. 2013). Estos hábitats se ubican sobre áreas con suelos pobremente drenados, topográficamente planos y sometidos a inundaciones temporales y permanentes (Junk y Piedade 2010). En particular, la palmera Mauritia flexuosa forma densas poblaciones en pantanos permanentemente inundados de la región Amazónica (Junk y Piedade 2010). Estudios realizados por ter Steege et al. (2013) estiman que poblaciones de M. flexuosa cubren c.a. 3 millones hectáreas en la Amazonía, y se calcula una población total de 1.500 millones de troncos de esta palmera. En este contexto, M. flexuosa es un elemento forestal hiperdominante en la Amazonía. Mauritia flexuosa, localmente denominada morete, es una palmera dioica (sexo separados), silvestre, solitaria con tallos de > 25 m de altura y diámetro c.a. 40 cm. Cada individuo tiene en promedio ocho inflorescencias que pueden alcanzar hasta 2 m de largo (Borgtoft-Pedersen y Balslev 1993; Henderson et al. 1995). Sus frutos son drupas esféricas, cubiertas por escamas rojas con un mesocarpio amarillo carnoso rico en ácidos grasos insaturados (Borgtoft-Pedersen y Balslev 1993). M. flexuosa juega un rol clave en la ecología del bosque debido a que: (i) es altamente abundante y es una de las pocas palmeras que tienen adaptaciones fisiológicas (neumatóforos) para vivir en medios inundados, (ii) aporta en la captura y fijación del carbono a través de sus enormes estructuras vegetativas que son incorporadas al sistema de pantano como materia orgánica, (iii) mantiene múltiples interacciones con polinizadores, dispersores y descomponedores (Gragson 1995; Horn et al. 2012; Endress et al. 2013). Esta palmera produce una gran cantidad de flores que atraen a insectos polinizadores (Henderson et al. 1995); además, puede generar hasta dos mil frutos por infructescencia (Gragson 1995) que son consumidos por gran diversidad de fauna (tapires, saínos, monos, aves y roedores) y por el hombre (Endress et al. 2013). M. flexuosa es un recurso importante por la multiplicidad de usos que brinda a las comunidades indígenas de la Amazonía (Endress et al. 2013). En particular sus frutos son cosechados para la obtención de aceites derivados del mesocarpio y para preparar una variedad de alimentos y bebidas (Rojas et al. 2001). Adicionalmente, las fibras de las hojas se usan para hacer cestos, esteras y techos de las chozas (Martins et al. 2012). En la Amazonía peruana se ha reportado prácticas extractivas destructivas a gran escala para la comercialización de frutos procesados y sin procesar (Hiraoka 1999; Rojas et al. 2001). Esta tala destructiva de palmas femeninas para obtener sus frutos tiene un impacto negativo en la conservación de este recurso forestal. En la región septentrional del Parque Nacional Yasuní (PNY) se calcula que los pantanos cubren c.a. 3% del área del parque (Romero-Saltos et al. 2001). En esta zona, los pantanos con M. flexuosa son áreas pequeñas 114 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 (1-20 ha), distribuidos en parches y aislados por sistemas de colinas (< 20 m de alto) dentro del bosque de terra firme (Romero-Saltos et al. 2001). Debido a esta distribución espacial se sugiere que las poblaciones de M. flexuosa se comportan como una metapoblación. Una metapoblación se describe como un sistema de subpoblaciones (demes) espacialmente discretas conectadas por mecanismos que favorecen el intercambio de semillas y de polen (Hanski y Gaggiotti 2004). El flujo génico (dispersión y polinización) es esencial para mantener la conectividad biológica entre las subpoblaciones (Bossuyt 2007). El conocimiento sobre las relaciones ecológicas y genéticas dentro de estos hábitats pantanosos es desconocido, a pesar de la importancia ecológica de esta especie, su potencial económico y la amenaza a su conservación. Los objetivos de este estudio son: (a) analizar la estructura genética de las poblaciones de M. flexuosa en cinco pantanos ubicados en la región norte del PNY, (b) evaluar la variación genética entre cohortes (adultos y plántulas) de esta palmera, (c) analizar si el modelo genético de M. flexuosa se ajusta a un sistema de metapoblación. Metodología Se seleccionaron cinco pantanos en la región norte del PNY, tres pantanos (P1, P2, P3) en el km 32, distanciados entre 1-2 km de la carretera Maxus, cerca de la comunidad Huaorani de Guillero; y dos pantanos (P4, P5) en el km 40, distanciados entre 0,5-1 km de la misma carretera. En ambas áreas, los pantanos se encontraban dispersos dentro del bosque de terra firme no disturbado, pero relativamente cerca de comunidades Huaorani (Fig. 1). En cada pantano se analizó la demografía de la palmera a través de transectos (250 x 5 m); se tomaron muestras de suelo (~ 100 gr) en cada pantano para su posterior análisis. Los individuos censados fueron clasificados como: plántulas (individuos < 0,5 m, con hojas palmeadas y con la semilla unida al individuo), juveniles (individuos de 0,5-8 m de altura, sin tallo y hojas en forma de roseta), subadultos (individuos con 8-16 m de altura, con tallo pero sin estructuras reproductivas) y adultos (individuos con tronco > 16 m de altura con estructuras reproductivas). Para los análisis genéticos se seleccionó el tejido vegetal de 30 adultos (n = 150) y de 30 plántulas (n = 150) en cada pantano. El genotipaje fue realizado con ocho marcadores microsatélites (SSRs) específicos para M. flexuosa (Federman et al. 2011). Con el fin de evaluar los niveles de diversidad genética entre los pantanos se calcularon índices de diversidad (diversidad alélica A, heterocigocidad esperada He, heterocigocidad observada Ho y coeficiente de endogamia FIS). Los análisis se realizaron para individuos adultos y plántulas por separado. Con el fin de evaluar la divergencia genética entre pantanos se realizó un análisis de agrupamiento Bayesiano en el programa STRUCTURE v.2.3 (Pritchard et al. 2010). El número de agrupamientos (K) fue determinado mediante el estadístico ΔK desarrollado por Evanno et al. (2005). Adicionalmente, se realizó análisis de la varianza molecular (AMOVA) y se calcularon valores diferenciación (FST) y el número de migrantes (Nm). Resultados La demografía de M. flexuosa mostró un mayo - -202) y -39) y -39). El número de hembras (207 individuos) fue mayor al de rango: 4,55-4,77) y que él porcentajes reportados en suelos de terra firme -17,5%) comparado con 115 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 P2) a limo-arcillosos (P3, P4, P5) y la suma de cationes (Ca++, Mg++, Na+, K+ cmol/kg. Los resultados de los análisis genéticos mostraron que en promedio la diversidad alélica (A), el número de alelos privados (PA), y la heterocigosidad esperada (He) fueron mayores en plántulas (A PA He A PA He el coeficiente de endogamia (FIS) fue mayor en plántulas (FIS FIS Los valores del índice de fijación (FST) fueron altamente significativos para todas las comparaciones poblacionales (FST : 0,12; p < 0.001; Tabla 2). Los resultados del AMOVA revelaron que, tanto para plántulas (90,71%) como para adultos (82,49%), la mayoría de la variación genética se encuentra dentro de las poblaciones, y esta partición fue altamente significativa en ambas cohortes. Sin embargo, el porcentaje de variación entre poblaciones fue relativamente alto considerando la reducida escala espacial entre los pantanos (0,3 a 8 km). Estos valores concuerdan con el bajo número de migrantes entre pantanos calculado (Nm = 0.29) y con la fuerte discriminación de los individuos a su respectiva población según los análisis de asignación Bayesiana con STRUCTURE (Fig. 3). Discusión Equilibrio ecológico de las poblaciones de Mauritia flexuosa en el PNY La estructura demográfica en cada pantano mostró un patrón de “J” invertida con un mayor número de plántulas y adultos que de juveniles y subadultos. Holm et al. (2008) reportaron patrones similares en poblaciones de M. flexuosa en Cuyabeno (Amazonía ecuatoriana) con un mayor número de plántulas y adultos. Contrario a este patrón de “J” invertida, en la región de Maijuna (Amazonía peruana) se describieron poblaciones de M. flexuosa con alta densidad de plántulas pero con una baja densidad de adultos (Horn et al. 2012). La baja densidad de adultos podría ser explicado por el efecto de la cosecha destructiva de frutos que involucra la tala de individuos hembra (Horn et al. 2012). En las poblaciones estudiadas de M. flexuosa en el PNY las hembras fueron más abundantes que los machos, a pesar que estos valores variaron ligeramente entre pantanos (♀/♂ = 1,09:1-1,86:1). Sin embargo, otros estudios en la región reportaron un patrón diferente al observado en nuestro estudio. Por ejemplo, en la región del Cuyabeno se describió un mayor número de machos (90 individuos/ha) que de hembras (70 individuos/ha; Holm et al. 2008); al igual que en la localidad de Maijuna (73 machos/ha sobre 21 hembras/ha; Horn et al. 2012). La tala de individuos hembra por la cosecha de frutos ocasiona desequilibrios demográficos en las poblaciones de palmeras (Freckleton et al. 2003; Endress et al. 2013). Habitantes de la zona norte del PNY relatan que la tala de las hembras por sus frutos no es una práctica común en la zona; ésta evidencia indirecta explicaría la mayor proporción de hembras que de machos en el área de estudio. Las características edáficas de los pantanos estudiados mostraron valores similares a otros pantanos de la Amazonía (Khan 1991; Lips y Duivenvoorden 2001; Poulsen et al. 2006; San José et al. 2010). Evaluación de la estructura genética poblacional 116 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Los resultados sugieren que la deriva genética y la endogamia son factores importantes en la estructura genética de M. flexuosa. A pesar que, las poblaciones estudiadas son pequeñas y aisladas físicamente por distancias cortas (0,3 a 8 km), estas reportaron una alta diversidad genética (He 0,71). Otros estudios en palmeras realizados a escalas espaciales mayores describieron una menor diversidad, por ejemplo en Euterpe edulis (He = 0,26; Conte et al. 2003) y en Pseudophoenix ekmanii (He 0,57; Namoff et al. 2011). Curiosamente, a pesar que las poblaciones de M. flexuosa están espacialmente cerca y poseen una alta diversidad genética, el valor de endogamia (FIS) en cada pantano fue alto (FIS plántulas = 0.28; adultos = 0.23). En poblaciones de Euterpe edulis en Brasil se describieron valores de FIS bajos (FIS plántulas = 0.01; adultos = 0,11) a pesar de que las poblaciones se encontraban a 22 km de distancia (Gaiotto et al. 2003). En poblaciones de Pseudophoenix ekmanii, separadas por distancias de 4-30 km, presentaron valores de endogamia < 0,25 (Namoff et al. 2011). La fuerte endogamia observada en este estudio sugiere que el flujo génico entre pantanos es muy limitado. Fuerte estructuración genética en cada población de Mauritia flexuosa La diferenciación genética (FST) entre las cinco poblaciones de M. flexuosa fue sorprendentemente alta a pesar de las cortas distancias geográficas entre los pantanos. Estudios similares en poblaciones de Euterpe edulis (FST = 0,01-0,09; Conte et al. 2003) y Pseudophoenix ekmanii (FST = 0,07-0,22; Namoff et al. 2011) reportaron valores FST más bajos que los obtenidos en este estudio, a pesar de encontrarse a escalas espaciales mayores (4 a 30 km). Los altos valores de diferenciación obtenidos (FST) indican que el flujo génico entre los pantanos es limitado y que cada población mantiene su propia huella genética construida por limitaciones en el flujo génico y por la deriva. La asignación Bayesiana indicó también que la mayoría de los individuos (96%) tuvieron una asignación superior al 95% de pertenecer a un respectivo pantano; y solo pocos individuos (4%) fueron asignados con un porcentaje entre 85%-95%. Este limitado flujo génico entre pantanos reportado por los valores FST y los análisis Bayesianos es coherente con el número bajo de migrantes (Nm = 0,29); esto sugiere que a pesar de la fuerte divergencia genética se mantienen mecanismos que permiten un flujo génico reducido pero constante entre pantanos. Adicionalmente, la presencia de alelos privados en la cohorte de plántulas refleja el ingreso de nuevos alelos vía reclutamiento de plántulas (Chaves et al. 2010). Ellstrand y Elam (1993) sugirieron que, en caso de que la deriva génica esté actuando sobre las poblaciones, un Nm de 0,5 será suficiente para impedir una fuerte diferenciación. En nuestro caso, el reducido número de migrantes sugiere que las poblaciones están en proceso de divergencia genética. Sin embargo, en poblaciones pequeñas un migrante por generación es suficiente para asegurar que los mismos alelos serán compartidos entre los individuos durante largos períodos de tiempo evolutivo (Lowe y Allendorf 2010). En el caso de las poblaciones de M. flexuosa unos pocos migrantes pueden ser lo suficientemente eficaces para mantener a estas poblaciones con una alta diversidad genética a pesar de los significantes valores de endogamia y de diferenciación biológica. ¿Podrían los pantanos de Mauritia flexuosa en Yasuní considerarse un sistema de metapoblación? En las plantas, existen tres parámetros que se deben cumplir para que las poblaciones formen un modelo de metapoblación: (i) las subpoblaciones deben estar en parches espacialmente separados, (ii) las subpoblaciones deben exhibir un cierto nivel de migración entre ellas, y (iii) poder recolonizar una subpoblación después de un proceso de extinción local (Hanski y Gaggiotti 2004). Las poblaciones de M. 117 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 flexuosa en este estudio son pequeñas y aisladas en parches; y presentan cierto grado de migrantes entre ellas (Nm = 0,29). Los resultados de este estudio sugieren que bajo estos parámetros propuestos por Hanski y Gaggiotti (2004), las poblaciones de M. flexuosa en la región norte del Yasuní forman un sistema de metapoblación que se mantienen con bajo flujo de genes y con tasas relativamente altas de endogamia. Implicaciones para la conservación de Mauritia flexuosa y su hábitat La presión humana por la cosecha de frutos de M. flexuosa es cada vez mayor en la región Amazónica (Rojas et al. 2001; Endress et al. 2013). En las poblaciones de M. flexuosa de la Amazonía peruana se han reportado la tala de hasta 1000 individuos hembras adultas por mes (Rojas et al. 2001). Este extractivismo destructivo genera un fuerte impacto negativo en la proporción de sexos y en la dinámica genética local de las poblaciones en pantanos. Estos cambios poblacionales limitaran la capacidad de resiliencia de las poblaciones a futuro (Horn et al. 2012). A diferencia de la Amazonía peruana, en la región Amazónica del Ecuador, la cosecha de frutos se realiza a escala familiar o comunitaria. Sin embargo, la escala de comercialización puede incrementarse a futuro producto de la demanda creciente y, si esta fuese acompaña con prácticas destructivas de cosecha, generaría impactos perjudiciales en la estructura ecológica y genética de las poblaciones. En particular, las poblaciones de M. flexuosa en el PNY son pequeñas (1-20 ha) consecuentemente una cosecha destructiva producirá cambios irreversibles en la resiliencia ecológica y genética de las poblaciones de esta palmera. Un factor importante que afecta a la conservación de los moretales es la defaunación. Debido a la disponibilidad de recursos, los pantanos con M. flexuosa son sitios de alimentación de una gran diversidad de animales frugívoros (Hiraoka 1999). Por este motivo, estas áreas constituyen sitios de interés de los habitantes locales para la cacería (Endress et al. 2013). En Yasuní se reportó que las actividades de caza redujeron en un 45% la fauna que se encuentra cerca de los asentamientos de las comunidades (Suárez et al. 2012). Estos datos sugieren una fuerte defaunación en la Amazonía Ecuatoriana con implicaciones negativas en la dinámica del bosque. El flujo génico en poblaciones de M. flexuosa es muy limitado y, si estas tasas de defaunación se incrementan, afectarían negativamente el flujo génico hasta romper la débil conectividad genética entre estas poblaciones. El presente estudio recomienda investigaciones de este importante y estratégico recurso forestal, el cual presenta un frágil equilibrio en su dinámica genética y ecológica. Adicionalmente, mecanismos de intercambio de material genético entre pantanos podría ser una herramienta importante para enriquecer la base genética de las poblaciones de morete en el país. Agradecimientos El presente proyecto fue financiado por la Pontificia Universidad Católica del Ecuador y el proyecto Europeo PALMS FP-7- Impactos de la cosecha de productos de palmas en bosques tropicales Bibliografía Borgtoft-Pedersen, H; Balslev, H. 1993. Palmas útiles: especies ecuatorianas para agroforestería y extractivsmo, Editorial Abya Yala, Quito, Ecuador. 118 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Bossuyt, B. 2007. Genetic Rescue in an Isolated Metapopulation of a Naturally Fragmented Plant Species, Parnassia palustris. Conservation Biology. 21: 832−841. Chaves, L; Vencovsky, R; Mendoça, R; Campos-Telles, M; Zucchi, M; Guedes, A. 2010. Estimating inbreeding depression in natural plant populations using quantitative and molecular data. Conservation Genetics. 12:569576. Conte, R; Nodari, O; Vencovsky, R; dos Reis, S. 2003. Genetic diversity and recruitment of the tropical palm, Euterpe edulis Mart., in a natural population from the Brazilian Atlantic Forest. Heredity. 91:401-406. Ellstrand, C; Elam, D. 1993. Population Genetic Consequences of Small Population Size: Implications for Plant Conservation. Annual Review of Ecology and Systematics. 24:217-242. Endress, B; Horn, C; Gilmore, M. 2013. Mauritia flexuosa palm swamps: Composition, structure and implications for conservation and management. Forest Ecology and Management. 302:346-353. Evanno G, Regnaut S, Goudet J (2005) Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Molecular Ecology. 14:2611-2620. Federman, S; Hyseni, C; Clement, W; Caccone, A. 2011. Isolation of 13 novel highly polymorphic microsatellite loci for the Amazonian Palm Mauritia flexuosa L. f. (Arecaceae). Conservation Genetics Resources. 4:355-357. Freckleton, R; Silva-Matos, D; Bovi, M; Watkinson, A. 2003. Predicting the impacts of harvesting using structured population models: the importance of density-dependence and timing of harvest for a tropical palm tree. Journal of Applied Ecology. 40:846-858. Gaiotto, A; Grattapaglia, D; Vencovsky, R. 2003. Genetic Structure, Mating System, and Long Distance Gene Flow in Heart of Palm (Euterpe edulis Mart.). J Hered 94:399-406. Gragson, T. 1995. Pumé explotation of Mauritia flexuosa (Palmae) in the Llanos of Venezuela. Journal of Ethnobiology. 15:177-188. Hanski, I; Gaggiotti, O. 2004. Ecology, Genetics, and Evolution of Metapopulations, Elsevier Academic Press, London, United Kingdom. Henderson, A; Galeano, G; Bernal, R. 1995. Field Guide to the Palms of the Americas, Princeton University Press, United States, New Jersey. Hiraoka, M. 1999. Miriti (Mauritia flexuosa) Palms and their uses and management among the Ribeirinhos of the Amazon Estuary. In: Padoch, C; Ayres, J; Pinedo-Vasquez, M; Henderson, A. (eds) Várzea. Diversity, development, and conservation of Amazonia’s Whitewater floodplains. The New York Botanical Garden Press, New York. Holm, J; Miller, C; Cropper, W. 2008. Population Dynamics of the Dioecious Amazonian Palm Mauritia flexuosa: Simulation Analysis of Sustainable Harvesting. Biotropica. 40:550-558. Horn, C; Gilmore, M; Endress, B. 2012. Ecological and socio-economic factors influencing aguaje (Mauritia flexuosa) resource management in two indigenous communities in the Peruvian Amazon. Forest Ecology and Management. 267:93-103. Junk, W; Piedade, M. 2010. An Introduction to South America Wetland Forest: Distribution, Definitions and General Characterization. In: Junk, W; Wittmann, F; Parolin, P; Piedade, M; Schöngart, J. (eds) Amazon Floodplain Forest. Springer Science+Business Media, New York, USA. Kahn, F. 1991. Palms as key swamp forest resources in Amazonia. Forest Ecology and Management. 38:133142. Namoff, S; Veloz, A; …; Francisco-Ortega, J. 2011. Sweet drinks are made of this: conservation genetics of an endemic palm species from the Dominican Republic. Journal of Heredety. 102:1-10. 119 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Lips, J; Duivenvoorden, J. 2001. Caracterización ambiental. In: Duivenvoorden, J; Balslev, H; Cavelier, J; Grandez, C; Tuomisto, H; Valencia, R. (eds) Evaluación de recursos vegetales no maderables en la Amazonía noroccidental, IBED, Universiteit van Amsterdam, Amsterdam. Ellstrand, C; Elam, D. 1993. Population Genetic Consequences of Small Population Size: Implications for Plant Conservation. Annual Review of Ecology and Systematics 24:217-242. Martins, R; Filgueiras, T; Albuquerque, U. 2012. Ethnobotany of Mauritia flexuosa (Arecaceae) in a Maroon Community in Central Brazil. Economic Botanic 66:91-98. Poulsen, A; Tuomisto, H; Balslev, H. 2006. Edaphic and floristic variation within a 1-ha plot of lowland Amazonian rain forest. Biotropica. 38:468-478. Pritchard J, Stephens M, Donnelly P (2000) Inference of population structure using multilocus genotype data. Genetics. 155:945-959. Rojas, R; Salazar, F; …; Panduro, F. 2001. Industrialización primaria del aguaje (Mauritia flexuosa L.f) en Iquitos (Perú). Folia Amazónica. 12:107-122. Romero-Saltos, H; Valencia, R; Macía, M. 2001. Patrones de diversidad, distribución y rareza de plantas leñosas en el Parque Nacional Yasuní y la Reserva Étnica Huaorani, Amazonía ecuatoriana. In: Duivenvoorden J, Balslev H, Cavelier J, Grandez C, Tuomisto H, Valencia R (eds) Evaluación de recursos vegetales no maderables en la Amazonía noroccidental, IBED, Universiteit van Amsterdam, Amsterdam. San José, J; Montes, R; Mazorra, M; Matute, N. 2010. Heterogeneity of the inland water-land palm ecotones (morichals) in the Orinoco lowlands, South America. Plant Ecol 208:259-269. Suárez, E; Zapata-Ríos, G; Utreras, V; Strindberg, S; Vargas, J. 2012. Controlling access to oil roads protects forest cover, but not wildlife communities: a case study from the rainforest of Yasuní Biosphere Reserve (Ecuador). Animal Conservation. doi:10.1111/j.1469-1795.2012.00592.x. ter Steege, H; Pitman, N; …; Silma, M. 2013. Hyperdominance in the Amazonian Tree Flora. Science. doi:10.1126/science.1243092. Figuras Fig. 1. Mapa de la ubicación de los cinco pantanos (en negro) muestreados de M. flexuosa en la región norte del PNY. 120 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Fig. 2. Demografía de las cinco poblaciones de M. flexuosa. A. Gráfico de barras de la estructura demográfica de cada pantano estudiado (P1 a P5). B. Gráfico de pastel del ratio sexual total de las cinco poblaciones estudiadas. Fig. 3 Resultados del análisis de asignación Bayesiana realizado con el programa STRUCTURE A. Determinación del número de agrupamientos genéticos (K = 5) de los individuos de M. flexuosa. B. Asignación de los individuos a un determinado agrupamiento genético (pantanos). Los valores q son la proporción de un genotipo en un grupo determinado. 121 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Tablas Tabla 1. Diversidad genética de plántulas y adultos en las cinco poblaciones de M. flexuosa en el PNY. A Ho He FIS Pantano Plántulas (n= 30) Adultos Plántulas (n= 30) Adultos Plántulas (n= 30) Adultos P1 6,75 7,13 0,54 0,62 0,68 P2 7,5 6,63 0,51 0,49 P3 7 6,75 0,48 P4 6,63 6,5 P5 6,75 Media 6,93 Plántulas (n= 30) Adultos (n= 30) 0,77 0,31 0,22 0,76 0,71 0,32 0,25 0,50 0,77 0,76 0,24 0,18 0,55 0,52 0,68 0,70 0,20 0,19 6 0,53 0,49 0,70 0,63 0,35 0,31 6,60 0,52 0,52 0,72 0,71 0,28 0,23 (n= 30) (n= 30) (n= 30) A, la diversidad alélica; Ho, heterocigosidad observada; He, heterocigosidad esperada; FIS, coeficiente de endogamia. Los valores FIS para ambos cohortes fueron estadísticamente significativos p <0,001 Tabla 2. Valores del índice de diferenciación (FST) de las cinco poblaciones de M. flexuosa en la parte inferior, con su respectiva significación estadística en la parte superior (** p < 0,001). P1 P2 P3 P4 P5 0 0,118 0,107 0,128 0,131 ** 0 0,099 0,133 0,124 ** ** 0 0,117 0,110 ** ** ** 0 0,139 ** ** ** ** 0 122 P1 P2 P3 P4 P5 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 PRIORIDADES DE INVESTIGACIÓN GENÉTICA Y ECOLÓGICA PARA LA CONSERVACIÓN DE ESPECIES Y BOSQUES Y EL DESARROLLO FORESTAL EN ECUADOR Javier Montalvo1,2, Rommel Montúfar3, Jimmy Cevallos4 1Escuela Superior Politécnica del Litoral, Guayaquil; Universidad de Vigo, Vigo (España); 3Pontificia Universidad Católica del Ecuador, Quito; 4Universidad Laica Eloy Alfaro de Manabí, Manta 2 [email protected] Bosques y desarrollo forestal en Ecuador: realidad e investigación pública Los bosques del Ecuador continental ocupan 12,8 millones de hectáreas, el 60,7% de la superficie total potencial de bosques (Sierra 2013). No toda esta superficie corresponde a bosques intactos: resulta del balance entre procesos históricos antropogénicos (deforestación y degradación) y ecológicos (regeneración natural por sucesión). La deforestación ha sido más relevante en la Costa y la Sierra (valles interandinos y vertiente andina occidental), mientras los bosques de la Amazonía se han conservado en mayor proporción (MAE 2012a). La tasa de deforestación anual neta promedio en Ecuador continental es aproximadamente 75.000 hectáreas (periodo 2000-2008) y, aunque ha disminuido, resulta de una deforestación bruta anual de más de 140.000 hectáreas contrarrestada por una regeneración anual aproximada de 67.000 hectáreas (Sierra 2013). Este autor indica que el 40% de la deforestación bruta actual del Ecuador continental está en la Costa (en especial en las provincias de Esmeraldas y Manabí), siendo también relevante en la Amazonía (Morona Santiago y Zamora Chinchipe). Ecuador es uno de los países clave para proteger la biodiversidad a nivel mundial, y alcanzar las metas del Convenio sobre Diversidad Biológica (Joppa et al. 2013). Cuenta con 17.748 especies nativas de plantas (León-Yánez et al. 2011) –muchas propias de ecosistemas forestales y una alta proporción son endémicas, en su mayoría amenazadas–, y una notable diversidad de ecosistemas terrestres naturales, con 66 tipos diferentes de bosque (MAE 2013). En Ecuador se explotan al menos 750 especies arbóreas, la mayoría nativas: el 45% es maderable y el 48% genera productos forestales no maderables (Añazco et al. 2010). Deforestación y degradación forestal son determinantes de erosión genética; fragmentación, alteración y desaparición de hábitats; extinción de especies, y reducción de diversidad de especies en los bosques a escala local, así como un deterioro en la provisión de servicios ecosistémicos. La forestación y reforestación es prioritaria en la agenda de desarrollo (MAE 2012b). En el periodo 2013-2017 la meta anual es 44.000 hectáreas de plantaciones forestales con fines de producción, protección y restauración, superando el bajo promedio de los últimos 28 años (6.600 hectáreas al año). La necesidad de conservación y restauración de la biodiversidad y servicios de los bosques es apremiante. Las políticas públicas afrontan los desafíos en el ámbito forestal mediante incentivos a la protección de los bosques, y a la forestación y reforestación, opciones relevantes para mitigar el cambio climático (secuestro y disminución de emisiones de CO 2). Sin embargo, en general la investigación forestal con financiación pública no está conectada con la realidad forestal ni los desafíos mencionados. La investigación forestal en Ecuador es escasa, muy limitada en ecología y genética, y restringida a contadas universidades y muy recientemente al Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP; Añazco et al. 2010; Grijalva et al. 2012). En el periodo 2006-2013, la inversión pública 123 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 en investigación vía SENESCYT refleja el interés por iniciativas en el ámbito forestal sensu lato (29 proyectos financiados). En este periodo se invirtió un monto de 8.156.941 USD, del cual fue destinado alrededor del 50% al INIAP. El 30% fue invertido en siete proyectos sobre sistemas agroforestales y silvopastoriles, el 20% en proyectos de cuencas hidrográficas y el 14% en cinco iniciativas de producción forestal y adaptación agroforestal. El 42% del monto de inversión fue a proyectos desarrollados en los Andes, 33% en la Amazonía, y apenas un 9% en la Costa; el resto corresponde a investigaciones desarrolladas en más de una región. Según el concepto de áreas prioritarias para la investigación de la SENESCYT, el 50% de inversión fue destinado a la generación de conocimiento e innovación en sistemas, y apenas un 3% a la conservación y uso de sistemas forestales. No existen iniciativas que exploren a largo plazo los bosques como ecosistemas o la ecología de las especies forestales individuales, que son prioritarias para su conservación y manejo integral. Solo tres proyectos abordan la investigación en el área de genética; de éstos, apenas un proyecto enfoca de forma explícita la importancia del estudio de la diversidad genética, la caracterización molecular y la resiliencia de poblaciones; éste representó el 8,5% del monto total invertido por SENESCYT en este periodo. Este análisis revela que la inversión pública en investigación forestal está desequilibrada en su distribución geográfica y presenta una conexión muy débil con la realidad forestal y sus principales desafíos agroproductivos, ambientales y de conservación. Prioridades de investigación genética La protección del patrimonio genético de las especies forestales es una prioridad y debe ser estudiada en dos dimensiones: diversidad genética domesticada y diversidad genética silvestre. a) Protección, manejo y uso sostenible de la diversidad genética domesticada. La diversidad domesticada está compuesta por especies vegetales que han sido sujetas a selección artificial por los pueblos amerindios para generar productos (frutos, tubérculos, fibras, semillas, etc.) con características útiles para el ser humano. Esta diversidad domesticada es producto de la interacción hombre-recurso durante cientos a miles de años, en los que se han seleccionado variedades con particulares características morfológicas, bioquímicas y fisiológicas. La domesticación es un proceso de selección genética que ha producido una rica variabilidad cultivada con potencial real de explotación comercial. En particular, las leguminosas y tubérculos andinos han sido el foco de investigación de la diversidad domesticada, debido a su importancia para la seguridad alimentaria; en cambio poco se ha explorado de las especies domesticadas de las zonas tropicales bajo los 1.000 msnm. Cacao (Theobroma cacao) y yuca (Manihot esculenta) son ejemplos de recursos amazónicos con estrategias de conservación, manejo y explotación de esta variabilidad domesticada. Pero también hay ejemplos de especies domesticadas subvaloradas como los frutos de chontaduro (Bactris gasipaes), uva de monte (Pourouma cecropiifolia), chirimoya (Anona spp.), guaba (Inga edulis, I. spectabilis), y palma de fibra (Aphandra natalia), entre otras. Por ejemplo, la domesticación del chontaduro ha sido dirigida a producir un fruto rico en carbohidratos y ácidos grasos. A pesar de ser una importante fuente de carbohidratos y aportar a la seguridad alimentaria en la Costa y la Amazonía, conocemos y protegemos muy poco de la variabilidad genética de esta especie (Montúfar y Rosas 2013). La diversidad genética del chontaduro está amenazada por la presencia de plantaciones comerciales de palmito (Bactris gasipaes de cultivo intensivo), las cuales a través de plantaciones de clones o genotipos seleccionados crean escenarios que favorecen la erosión genética de los cultivares locales (Couvreur et al. 2005). La investigación de la diversidad domesticada debe dirigirse a comprender: (i) la diversidad genética y fenotípica y determinar los centros de origen de la domesticación; (ii) la dinámica de las poblaciones domesticadas y su resiliencia genética (capacidad de recuperación); (iii) los mecanismos de erosión y contaminación genética, y (iv) los sistemas de conservación 124 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 in situ y ex situ. Las técnicas moleculares actuales permiten el uso de una gran variedad de herramientas genéticas para explorar la domesticación y su dinámica. b) Conservación, manejo y uso sostenible de la diversidad genética silvestre. La biodiversidad forestal, entendida como la variedad de especies y diferencias naturales de sus poblaciones en la flora nativa de los bosques, es un recurso estratégico, y consecuentemente su uso, manejo, y explotación deben basarse en estudios científicos apropiados. Es fuente de numerosos recursos, incluyendo productos forestales no maderables, y representa una invaluable diversidad genética con potenciales aplicaciones en biotecnología y medicina. Está amenazada por las actividades antropogénicas mencionadas que afectan a los bosques, y otras como cambio climático, contaminación y especies invasoras... Los análisis de la dinámica de la diversidad genética de una especie forestal constituyen la base fundamental para la planificación del uso y conservación del recurso, sin embargo estos estudios son muy limitados en el país. El informe ‘Situación de los recursos genéticos forestales’ en Ecuador (Grijalva et al. 2012) reporta estudios de diversidad genética para menos de una docena de especies silvestres, y se desconoce si la información generada ha sido sometida a evaluación y publicación científica. Por otro lado, para especies con gran valor maderable como laurel (Cordia alliodora), pigüe (Pollalesta discolor), sande (Brosimum utile), chalviande (Virola sebifera), cedro (Cedrela odorata), canelos (Nectandra spp.), chuncho (Cedrelinga cateniformis) y otras especies maderables amenazadas –como caoba (Swietenia macrophylla) o chanul (Humiriastrum procerum)– no hay estudios que permitan planificar su manejo y conservación. Adicionalmente, hay recursos forestales no maderables de importancia económica altamente amenazados por la erosión genética, como paja toquilla (Cardulovica palmata), guadúa (Guadua angustifolia) o tagua (Phytelephas aequatorialis), que requieren estudios de diversidad genética para diseñar sus planes de manejo. Un ejemplo de la importancia del estudio y conservación de los recursos genéticos forestales nativos es Elaeis oleifera, que es la palma aceitera americana. Constituye la especie ‘hermana’ de la palma africana (Elaeis guineensis), y un foco de interés para la ingeniería genética de esta especie de alta importancia económica. En la actualidad no existen estudios ecológicos ni genéticos de las poblaciones silvestres de este recurso forestal estratégico. La planificación de la conservación, manejo y explotación de los recursos genéticos forestales requiere desarrollar diferentes líneas de investigación: (i) estudios citológicos y la estructura cromosómica (desconocemos el número cromosómico de casi la totalidad de las especies de importancia forestal); (ii) estudios de diversidad con marcadores genéticos, bioquímicos y morfológicos informativos, diseños experimentales con escala y análisis de datos adecuados; (iii) estudios en genética de la conservación que permitan evaluar el impacto de las actividades antropogénicas en la viabilidad y resiliencia genética de las poblaciones; (iv) estudios filogenéticos que proporcionen información del estatus taxonómico y relaciones evolutivas de las especies de mayor importancia forestal, y (v) ecogenómica, área de conocimiento emergente que explora los mecanismos de la adaptación de las especies, su variabilidad fenotípica e interacciones, de especial importancia en las poblaciones de plantas silvestres (Ouborg y Vriezen 2007). Prioridades de investigación ecológica y para el desarrollo forestal La investigación ecológica forestal debe hacerse a varios niveles: especies, ecosistemas, y paisaje o territorio. Es recomendable que exista cooperación transdisciplinar y estratégica para fundamentar diferentes políticas sectoriales desde la visión holística e integradora propia de la ciencia ecológica para la conservación y uso sostenible de ecosistemas y especies forestales. A continuación se describen cuatro temas de investigación prioritarios, justificando su importancia o idoneidad con ejemplos y criterios relevantes. 125 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 a) Identificación de áreas y ecosistemas prioritarios de conservación, en especial con alta riqueza de especies endémicas de ámbito de distribución restringido. Las mayores concentraciones de plantas endémicas coinciden con áreas con alta riqueza de especies en general y de especies endémicas de aves, mamíferos y anfibios, destacando Ecuador a nivel mundial (Joppa et al. 2013). Las preguntas de investigación ecológica deben atender por una parte, a la viabilidad de las poblaciones y a las amenazas de su hábitat, considerando el tamaño de fragmentos de bosques remanentes y su estado de conservación, porque es imposible proteger todo el territorio. Por otra parte, a la variabilidad espacial en superficie, fragmentación, degradación estructural, vulnerabilidad, y relevancia para la conservación de los diferentes tipos de bosques remanentes. El área de distribución de la estrellita esmeraldeña (Chaetocercus berlepschi), un colibrí endémico amenazado de extinción por la pérdida y fragmentación de su hábitat, se limita al bosque siempreverde de tierras bajas de Esmeraldas. De las 4.500 especies de plantas endémicas de Ecuador, el 8% está en peligro crítico de extinción, como la liana Passiflora roseorum, cuya única población conocida habita en un bosque siempreverde montano alto del sur del país. Un tercio de las especies de plantas endémicas son orquídeas, el 85% presenta algún grado de amenaza y una alta proporción son epífitas propias de los bosques siempreverdes, en especial los montanos (Endara 2011). El ‘bosque seco’, que incluye varios tipos de bosque deciduo y semideciduo, requiere esfuerzos de conservación por su alta deforestación, fragmentación y degradación (Portillo-Quintero y Sánchez-Azofeifa 2010). Queda el 25% de la superficie potencial de este bosque, presenta un alto riesgo de perturbación, y sólo el 2,3% está protegido. De 239 especies de árboles y arbustos de los bosques secos de Ecuador identificadas por Aguirre et al. (2006), 47 son endémicas compartidas sólo con Perú (20% de endemismo) y 17 son exclusivas de Ecuador (7%). La identificación con alta resolución espacial de áreas susceptibles de restauración dentro de un área protegida de la Costa con bosques siempreverde estacional piemontano y deciduos promueve la conectividad y conservación de su biodiversidad (Esteban et al. 2013). b) Evaluación de la variabilidad espacial en la degradación y resiliencia de bosques, y su influencia sobre interacciones ecológicas clave, la distribución y conservación de especies, y atributos ecosistémicos. Existe un notable vacío de conocimiento sobre los cambios estructurales y funcionales en los bosques de Ecuador tras diferentes tipos de perturbación, y sobre su resiliencia o capacidad ecosistémica de recuperación. Por ejemplo, en general el efecto de la tala selectiva de especies arbóreas y otras perturbaciones sobre la diversidad de especies (riqueza y abundancia relativa de especies o ecodiversidad) es negativo (Clark y Covey 2012), sin embargo, la estructura y apertura del dosel o las condiciones climáticas de regeneración difieren mucho entre bosques deciduos o siempreverdes. Interesa conocer la capacidad y velocidad de recuperación de la composición florística, ecodiversidad y biomasa, el potencial de secuestro de CO 2 y de almacenamiento de carbono, y los principales factores condicionantes en diferentes ecosistemas forestales degradados que ocupan espacios sin presiones de uso en la Costa y en la Amazonía. La teledetección en áreas protegidas ayuda a comprender los mecanismos de regeneración natural (Nagendra et al. 2013), y a evaluar la eficacia y efectos de medidas de restauración de bosques o de reforestación comercial en zonas de amortiguamiento, de interés para el manejo a escala local y regional del Sistema Nacional de Áreas Protegidas. Así se fortalecerá el conocimiento ecosistémico del estado del patrimonio natural del Ecuador en general, y la contribución de los bosques ecuatorianos a la mitigación del cambio climático –manteniendo y recuperando stocks de carbono– y los cobeneficios para la conservación de la biodiversidad, de interés para implementar el Programa Nacional REDD+. 126 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 La alteración de la red de interacciones ecológicas entre las especies produce efectos que pueden depender del tipo de bosque y la densidad de poblaciones implicadas. Es preciso generar conocimientos básicos de la ecología de poblaciones de especies valiosas, indicadoras o amenazadas tras la degradación de su hábitat para su conservación, manejo y uso sostenible en su caso. La deforestación y degradación de bosques deciduos y semideciduos es una amenaza de extinción local para el pijío (Cavanillesia platanifolia), un árbol caducifolio que puede superar los 25 m de altura; en las cavidades de su tronco anida una subespecie endémica y en peligro crítico de extinción, el papagayo de Guayaquil (Ara ambiguus guayaquilensis). Otro ejemplo es la caza intensa, que en los bosques tropicales produce múltiples efectos indirectos y en cascada – comprobados o hipotéticos–, entre ellos cambios en los patrones de producción y dispersión de semillas, y de reclutamiento poblacional, con efectos potenciales sobre la densidad de algunas especies de plantas y diversidad del ecosistema a nivel local (Wright 2003). c) Influencia de la diversidad de especies arbóreas nativas, identidad y tipo de especie, y diseño en la producción de plantaciones forestales mixtas y en la restauración ecológica de diferentes bosques. Las plantaciones mixtas producen más que los monocultivos (Piotto 2008). Debe promoverse la investigación sobre tipos innovadores de plantaciones mixtas para restauración de bosques y producción forestal (Kelty 2006). Sin embargo, la Subsecretaría de Producción Forestal promueve en Ecuador las plantaciones forestales monoespecíficas –para la industria de la madera–, incentivando sólo 17 especies arbóreas, nueve nativas. Se debe procurar la innovación en la selección de especies nativas de árboles, investigando sus aptitudes, crecimiento y comportamiento en ensayos en vivero y plantaciones experimentales para optimizar el diseño y asegurar los fines de protección o comerciales en la forestación y reforestación en diferentes condiciones ambientales. Para ciertas especies objetivo y maderables, no basta con reconocer su predecible variabilidad genética, se requieren ensayos de procedencia apropiados, de progenie e incluso programas de mejora genética si existe heredabilidad (mayor crecimiento, calidad de fuste y madera, resistencia a condiciones adversas, etc.). Así se puede identificar y caracterizar germoplasma idóneo para diferentes tipos de manejo silvicultural, productos forestales o condiciones climáticas, tan contrastadas dentro de algunas regiones, como la Costa. Es necesario generar conocimientos ecológicos para conservar la biodiversidad y un manejo forestal sostenible, dentro y fuera de las áreas protegidas. En áreas de bosque seco, la restauración mediante enriquecimiento o plantaciones de especies nativas es la principal opción de manejo. Expande la superficie de hábitat necesaria para preservar muchas especies de fauna, en particular de vertebrados con poblaciones con baja densidad y viabilidad comprometida. Repoblar con higuerones (por ejemplo, Ficus insipida y F. obtusifolia), es oportuno porque constituye una fuente de alimento clave para el mono aullador (Alouatta palliata), también en la estación seca. El diseño de plantaciones forestales productivas y la restauración requieren la tipificación ecológica de las especies arbóreas, basada en el conocimiento de sus características, los factores limitantes de su crecimiento, incluyendo interacciones interespecíficas integrando en una comunidad. Según el tipo de bosque a restaurar o condiciones ambientales, varía la necesidad de combinar especies que difieren en su tolerancia a la sombra, velocidad de crecimiento en altura, estructura de copa o fenología foliar (en particular deciduo frente a siempreverde). En una plantación de balsa (Ochroma pyramidale) y cedro en la misma proporción en tierras bajas de la Costa, el crecimiento diametral de la balsa a los 34 meses fue el triple que el del cedro, que resultó dominado. Sin competencia de balsa, el cedro puede crecer al menos un 50% más, reflejando el diseño inadecuado de la plantación con estas dos especies nativas heliófilas. Es necesario investigar los diseños que combinen diferentes proporciones y densidad de 127 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 especies y promuevan tipos de interacciones que favorezcan la coexistencia de las especies y los fines del ecosistema forestal. Entre estas interacciones destaca la reducción de competencia interespecífica por complementariedad en el uso de recursos, y la facilitación, en la que una especie beneficia a otra, como una fijadora de nitrógeno a otra no fijadora. Es necesaria la evaluación experimental de interacciones entre especies, y ensayos de plantaciones mixtas a escala operativa, con propietarios y empresas del sector forestal. Por ejemplo, para acelerar la recuperación de la estructura, ecodiversidad y biomasa del bosques siempreverdes, pueden ensayarse la introducción de especies esciófilas de baja densidad, semillas grandes, dispersión por gravedad y tardías en la sucesión en plantaciones mixtas de especies heliófilas, pioneras y de crecimiento rápido como balsa. En áreas degradadas de la Costa, se puede ensayar la influencia de especies facilitadoras como algarrobo (Prosopis pallida), u otras especies arbóreas fijadoras de nitrógeno del subdosel, para promover la estabilidad y producción de una plantación mixta con otras especies maderables como guayacán (Tabebuia chrysantha), fernán sánchez (Triplaris cumingiana) y laurel. d) Identificación, valoración y restauración de servicios ecosistémicos forestales prioritarios. Se conoce poco sobre la variabilidad, procesos de generación y restauración de servicios ecosistémicos forestales (Mass et al. 2005, Hall et al. 2011). La provisión de agua, regulación del clima y el control de inundaciones son tres servicios principales del bosque seco (Mass et al. 2005). Es preciso conocer, por ejemplo, cómo los bosques y cambios de uso del suelo en cuencas hidrográficas (deforestación, regeneración natural, reforestación, establecimiento de sistemas agroforestales, etc.) afectan a su protección y valorar su influencia sobre la captación, provisión y ciclo del agua (Stallard et al. 2010) en diferentes regiones de Ecuador. Dicha información, derivada de estudios a largo plazo, a mesoescala y modelización, es crítica para un ordenamiento territorial con el objetivo de la gestión y restauración de servicios de los ecosistemas forestales. Conclusión Ecuador precisa conocimientos, genéticos y ecológicos, de especies arbóreas nativas, y de aspectos ecosistémicos estructurales y funcionales que sean estratégicos para la conservación y restauración de bosques y áreas prioritarias, su biodiversidad y servicios ecosistémicos. Y también para el diseño y manejo sostenible de plantaciones forestales con fines productivos o comerciales. En particular se debe priorizar el conocimiento de las aptitudes e interacciones ecológicas clave entre especies arbóreas nativas de interés para la conservación, restauración y uso sostenible de bosques, y la innovación en el diseño de sistemas agroforestales protectores y productores de servicios ecosistémicos. La investigación genética y ecológica en el ámbito forestal en Ecuador debe fortalecerse para atender a los desafíos ambientales y socioeconómicos. Puede generar conocimientos cruciales para el progreso y desarrollo sostenible del Ecuador, y en particular para el cambio de la matriz productiva. Se debe articular dentro de un Programa Nacional de Investigación Forestal elaborado de forma participativa (Añazco et al. 2010), con incentivos a la cooperación transdisciplinar y evaluación del impacto de sus resultados. La investigación forestal debe fundamentar las políticas públicas de conservación de la biodiversidad, forestación y reforestación, ordenamiento territorial y mitigación del cambio climático. Agradecimientos Este trabajo ha sido financiado por el Proyecto Prometeo de la Secretaría de Educación Superior, Ciencia y Tecnología (SENESCYT) a uno de los autores. Agradecemos a la Dirección de Orientación, Diseño y Coordinación de la Investigación de la SENESCYT los datos de proyectos de investigación con financiación 128 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 pública; a la empresa Tadel, SA las facilidades de acceso a su plantación forestal, y a Álvaro Pérez sus comentarios al manuscrito. Bibliografía Aguirre Mendoza, Z; Linares-Palomino, R; Kvist, LP. 2006. Especies leñosas y formaciones vegetales en los bosques estacionalmente secos de Ecuador y Perú. Arnaldoa 13:324-350. Añazco M; Morales, M; Palacios, W; Vega, E; Cuesta, A. 2010. Sector Forestal Ecuatoriano: propuestas para una gestión forestal sostenible. Quito, Ecuador. (Serie Investigación y Sistematización nº 8. Programa Regional ECOBONA-INTERCOOPERATION). Clark, JA; Covey, KR. 2012. Tree species richness and the logging of natural forests: A meta-analysis. Forest Ecology and Management 276:146-153. Couvreur, T; Billote, N; Risterucci, A; Lara, C; Vigouroux, Y; Ludeña, B; Pham, JL; Pintaud, JC. 2005. Close genetic proximity between cultivated and wild Bactris gasipaes Kunth revealed by microsatellite markers in western Ecuador. Genetic Resources and crop Evolution 53:1361-1373. Endara, L. 2011. Orquídeas endémicas de Ecuador. Consultado el 30 oct. 2011. http://www.flmnh.ufl.edu/ecuadororchids/Patrones_endemismo.htm Esteban, S; Baus, B; Segovia, G; López, Á; Jensen, R; Montalvo, J. 2013. Cartografía ecológica de la vegetación y sistemas de información geográfica: aplicación preliminar para la conservación y manejo en el Refugio de Vida Silvestre Marina y Costera Pacoche (Manabí). Comunicación presentada. 9 p. II Congreso Nacional de Áreas Protegidas (Quito, Ecuador, 29 al 31 jul. 2013). Grijalva, J; Checa, X; Ramos, R; Barrera, P; Limongi, R. 2012. Situación de los recursos genéticos forestales. Informe País Ecuador. Documento sometido a la Comisión Forestal de la FAO. 95 p. Hall, JS; Ashton, MS; Garen, EJ; Jose, S. 2011. The ecology and ecosystem services of native trees: Implications for reforestation and land restoration in Mesoamerica. Forest Ecology and Management 261:1553-1557. Maass, J; Balvanera, P; Castillo, A; Daily, GC; Mooney, HA; Ehrlich, P; Quesada, M; Miranda, A; Jaramillo, VJ; García-Oliva, F; Martínez-Yrizar, A; Cotler, H; López-Blanco, J; Pérez-Jiménez, A; Búrquez, A; Tinoco, C; Ceballos, G; Barraza, L; Ayala, R; Sarukhán, J. 2005. Ecosystem services of tropical dry forests: insights from long-term ecological and social research on the Pacific Coast of Mexico. Ecology and Society 10:17. [online] http://www.ecologyandsociety.org/vol10/iss1/art17/ Ouburg, NJ; Vriezen, WH. 2007. An ecologist’s guide to ecogenomics. Journal of Ecology 95:8-16. Stallard, RF; Ogden, FL; Elsenbeer, H; Hall, J. 2010. Panama Canal Watershed Experiment: Agua Salud Project. Water Resources Impact 12:17-20. Joppa, LN; Visconti, P; Jenkins, CN; Pimm, SL. 2013. Achieving the Convention on Biological Diversity’s Goals for Plant Conservation. Science 341:1100-1103. Kelty, MJ. 2006. The role of species mixtures in plantation forestry. Forest Ecology and Management 233:195204. 129 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 León-Yánez, S; Valencia, R; Pitman, N; Endara, L; Ulloa Ulloa, C; Navarrete, H (eds.). 2011. Libro rojo de las plantas endémicas del Ecuador, 2 ed. Quito, Ecuador, Publicaciones del Herbario QCA, Pontificia Universidad Católica del Ecuador. Ministerio del Ambiente (MAE). 2012a. Línea Base de Deforestación del Ecuador Continental. Quito, Ecuador. 31 p. MAE 2012b. Plan Nacional de Forestación y Reforestación. Quito, Ecuador. 56 p. MAE 2013. Sistema de Clasificación de los Ecosistemas del Ecuador Continental. Quito, Ecuador, Subsecretaría de Patrimonio Natural. 177 p. Montúfar, R; Rosas, J. 2013. Chontaduro, Chontilla. Bactris gasipaes. Palmeras ecuatorianas: biología y uso sostenible (Eds. R. Valencia, R. Montúfar, H. Navarrete, H. Balslev). Quito, Publicaciones del Herbario QCA, Pontificia Universidad Católica del Ecuador. Nagendra, H; Lucas, R; Pradinho Honrado, J; Jongmand, RHG; Tarantinoe, C; Adamoe, M; Paola Mairotaf, P. 2013. Remote sensing for conservation monitoring: Assessing protected areas, habitat extent, habitat condition, species diversity, and threats. Ecological Indicators 33:45-59. Ouborg, NJ.; Vriezen, WH. 2007. An ecologist’s guide to ecogenomics. Journal of Ecology 95:8-16. Piotto, D. 2008. A meta-analysis comparing tree growth in monocultures and mixed plantations. Forest Ecology and Management 255:781-786. Portillo-Quintero, CA; Sánchez-Azofeifa, GA. 2010. Extent and conservation of tropical dry forests in the Americas. Biological Conservation 143:144-155. Sierra, R. 2013. Patrones y factores de deforestación en el Ecuador continental, 1990-2010. Y un acercamiento a los próximos 10 años. Quito, Ecuador, Conservación Internacional Ecuador y Forest Trends. 51 p. Wright, SJ. 2003. The myriad consequences of hunting for vertebrates and plants in tropical forests. Perspectives in Plant Ecology, Evolution and Systematics 6:73-86. 130 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 ESTRUCTURA Y DIVERSIDAD GENÉTICA DE LA PALMERA Oenocarpus bataua Mart. (ARECACEAE): UNA HERRAMIENTA PARA LA CONSERVACIÓN DE UN RECURSO OLEAGINOSO FORESTAL Sebastián Escobar1, Rommel Montúfar1, Jean-Christophe Pintaud2 1 Pontificia Universidad Católica del Ecuador (PUCE), Lab. de Ecología y Genética (113), Av. 12 de Octubre y Av. Patria, Quito, Ecuador 2 Institut de Recherche pour le Développement (IRD), UMR DIAPE/DYNADIV, 911 Avenue Agropolis, B.P. 64501, 34394 Montpellier Cedex 5, France [email protected] Introducción Los bosques del Neotrópico albergan una riqueza considerable en términos de biodiversidad. En particular, los recursos forestales oleaginosos son de gran interés debido a la constante búsqueda de nuevas fuentes de aceites comestibles beneficiosos para la salud humana. En este sentido, el conocer la variabilidad intraespecífica de especies oleaginosas provenientes de bosques tropicales y subtropicales, así como las fuerzas que estructuran su diversidad genética, juega un rol estratégico en el manejo y conservación de estos importantes recursos forestales. La familia de las palmeras (Arecaceae) tiene aproximadamente 550 especies en el Neotrópico (Henderson et al. 1995). Varias palmeras son hiperdominantes en los bosques tropicales, como el pambil (Iriartea deltoidea), el palmiche (Euterpe precatoria) o la ungurahua (Oenocarpus bataua; Steege et al. 2013). Sus frutos, especialmente el mesocarpo, sintetizan una gran diversidad de ácidos grasos, propiedad metabólica aprovechada por el ser humano para el aislamiento y consumo de aceites (Montúfar et al. 2010). Especies como Oenocarpus bataua, Mauritia flexuosa, Euterpe oleracea, Bactris gasipaes, Elaeis oleifera, Attalea butyracea o Attalea colenda han sido tradicionalmente utilizadas como fuentes oleginosas en Ecuador (Montúfar & Brokamp 2011). El estudio y conservación de estos recusos provenientes del bosque tropical constituyen una estratégica línea de investigación a ser desarrollda en el país. La palmera Oenocarpus bataua, conocida localmente como ungurahua, es un valioso recurso oleaginoso proveniente de los bosques tropicales y subtropicales de Sudamérica y la isla de Trinidad (Henderson 1995; Montúfar et al. 2010). Ésta es una palmera silvestre, arborescente (>20 m de alto y ~20 cm de diámetro), monoica y principalmente alógama (Henderson 1995). O. bataua forma poblaciones moderadamente densas en bosques tropicales bajo los 800 m de altitud, e incluso alcanza elevaciones hasta los 1400 msnm en las estribaciones andinas (Henderson 1995). La polinización en O. bataua es realizada por insectos de los órdenes Coleoptera e Himenoptera (Núñez-Avellaneda & Rojas-Robles 2008), mientras que la dispersión de sus semillas es realizada por aves y mamíferos (Karubian et al. 2010). El mesocarpo de sus frutos es rico en ácido graso oleico (>70%, mono-insaturado) y en varios ácidos grasos poli-insaturados; y también presenta una alta concentración de precursores de la vitamina A (β-carotenos; Montúfar et al. 2010). Adicionalmente, la composición de ácidos grasos del aceite de O. bataua es similar al aceite de oliva (Olea europeae) en términos nutricionales (Montúfar & Brokamp 2011). 131 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 El presente estudio caracteriza la diversidad genética de O. bataua variedad bataua en el noroccidente de América del Sur con el objetivo de identificar patrones intraespecíficos de diversidad (agrupamientos genéticos o clusters). Los resultados son discutidos en un contexto biogeográfico, y están enfocados a la conservación de la variabilidad intraespecífica de este importante recurso forestal. Materiales y métodos Un total de 591 individuos de O. bataua var. bataua procedentes de 26 localidades (Fig. 1) fueron colectados en Colombia, Ecuador, Perú y Bolivia. La extracción de ADN se realizó con un Qiagen Dneasy plant mini kit siguiendo las instrucciones del fabricante; su concentración fue determinada con un espectrofotómetro Thermo Scientific NanoDrop™ ND-100 y finalmente diluida con agua ultra pura a 5 ng/µl. Siete marcadores SSR (Ob02, Ob06, Ob08, Ob16, AC5-3#4, AG5-5#1, AG1; Lepsch-Cunha et al. 2003; Montúfar et al. 2006; Ludeña et al. 2011) fueron utilizados en el genotipaje de las muestras utilizando la técnica Multiplex PCR. La amplificación PCR fue realizada en un volumen final de 10 µl como se indica: 4 µl del templado de ADN (5 ng/µl), 5 µl de Multiplex Mastermix 2X, y 1 µl de primer mix 10X (primers a 0.2 µM cada uno). Las condiciones PCR consistieron de una denaturación inicial de 15 min a 96 °C, seguido de 30 ciclos of denaturación de 30 s a 94 °C, hibridación de 90 s a 56 °C, y elongación de 90 s a 72 °C. Un paso de extensión final de 10 min a 72 °C fue añadido al final. El producto final de cada PCR fue diluido con agua y 0.15 μl del marcador de talla 500 LIZ (GeneScan™), resultando una dilución final de ~1/300. Los productos fueron analizados en un secuenciador 3500 Genetic Analyzer (Applied Biosystems™), y los cromatogramas resultantes fueron examinados con el software GeneMapper v3.7 (Applied Biosystems™). La identificación de patrones genéticos se realizó bajo dos metodologías: (i) Los patrones genéticos regionales (agrupamientos) dentro de O. bataua fueron determinados mediante un análisis de agrupamiento bayesiano en el programa Structure v.2.3 (Pritchard et al. 2010). El número de agrupamientos (K) fue determinado mediante el estadístico ΔK desarrollado por Evanno et al. (2005). Posteriormente, la información genética y geográfica de cada individuo fue integrada en el programa Geneland (Guillot et al. 2005) para determinar discontinuidades genéticas a nivel espacial. (ii) Los patrones genéticos a nivel poblacional fueron determinados mediante varios índices de diversidad genética (número promedio de alelos por locus - A, heterocigocidad esperada - He, heterocigocidad observada –Ho, coeficiente de endogamia - FIS) y estadísticos F, los cuales fueron obtenidos con el programa Arlequin 3.1 (Excoffier et al. 2005). A esta escala de análisis se utilizaron las localidades con n > 15 individuos, las cuales fueron tratadas como poblaciones. Resultados y discusión Cinco agrupamientos genéticos fueron identificados dentro del rango de distribución de O. bataua, los cuales coinciden con grandes ecoregiones del noroccidente de Sudamérica. Los agrupamientos identificados corresponden a: (i) el valle del río Magdalena (VRM), (ii) el Chocó, (iii) la zona Amotape-Huancabamba (ZAH), (iv) la región del Napo, y (v) la región de Inambari (Fig. 2). El agrupamiento del Napo está formado por dos regiones disyuntas: la cuenca del río Napo y el noroccidente de Bolivia. La mayor diversidad genética fue encontrada en Napo (He = 0.83, A = 16.57) e Inambari (He = 0.79, A = 16.42), mientras que la menor diversidad fue reportada para VRM (He = 0.54, A = 3.43; Tabla 1). Todos los agrupamientos reportaron valores moderados de endogamia cercanos a cero (FIS = -0.002 – 0.09). La mayor diferenciación genética fue determinada entre los agrupamientos VRM y Chocó (FST = 0.32), mientras que la menor se reportó entre Napo e Inambari (FST = 0.04). 132 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Los agrupamientos genéticos encontrados en O. bataua son explicados por: (i) el levantamiento de los Andes, y (ii) las condiciones climáticas actuales. La orogénesis andina separó físicamente la región del Chocó de la Amazonía, permitiendo que poblaciones de las regiones trans/cis andinas generen una huella genética propia (Trénel et al. 2008). Debido a la baja diferenciación genética entre el Chocó y los agrupamientos amazónicos (Napo e Inambari; FST = 0.09, 0.12, respectivamente), se infiere que a pesar de la presencia de los Andes como barrera biogeográfica, el flujo génico en O. bataua se ha mantenido entre las dos regiones a través de corredores andinos de menor altitud (Montúfar 2007) o por procesos de dispersión a larga distancia. Este proceso de orogénesis también dio paso a la formación de regiones andinas con características fitogeográficas particulares como el valle del río Magdalena (entre las cordilleras central y oriental en Colombia), donde se determinó la presencia del agrupamiento VRM; y la zona Amotape-Huancabamba (suroriente del Ecuador/noroccidente de Perú), donde se ubicó un peculiar agrupamiento genético (ZAH). Por otro lado, la estructura genética de O. bataua en la amazonía occidental está influenciada por variables climáticas modernas, principalmente precipitación anual y estacionalidad (número de meses secos al año; Kreft & Jetz 2007). La variación en precipitación y estacionalidad se correlaciona con la separación de los agrupamientos Napo e Inambari. Localidades como Yasuní, en la Amazonía ecuatoriana (Napo), presentan una precipitación anual de ~3200 mm, y no presentan estacionalidad (ningún mes bajo 100 mm). Por otro lado, localidades como Puerto Maldonado, ubicado en el sur de Perú (Inambari), presentan una precipitación anual de ~2300 mm, con >80% de ésta restringida entre abril y octubre (Pitman 2000). Adicionalmente, la ausencia de una barrera física actual al flujo génico entre las regiones de Napo e Inambari ha facilitado que estos agrupamientos mantengan un continuo flujo génico reflejado en los bajos valores de diferenciación obtenidos (FST = 0.04). A nivel poblacional, la mayor diversidad genética fue reportada en las poblaciones Intuto (He = 0.86, A = 11.71) y Jenaro Herrera (He = 0.81, A = 11.86; Tabla 2), ubicadas alrededor de la ciudad de Iquitos (norte de la Amazonía peruana, Departamento de Loreto). La alta diversidad genética de O. bataua reportada en esta región coincide con la localización de comunidades con alta diversidad de palmeras como Iquitos con 54 especies/2.75 ha, o Yasuní con 33 especies/2.5 ha (Vormisto et al. 2004). Adicionalmente, la localidad de La Pedrera (suroriente de Colombia, ~350 km de Iquitos) presenta la mayor diversidad de especies del género Oenocarpus (seis especies; Galeano & Bernal 2010). En base a: (i) la alta diversidad genética reportada para O. bataua, (ii) la alta diversidad de especies de palmeras descritas, y (iii) la alta diversidad del género Oenocarpus, se infiere que la región del Napo y sus alrededores podrían ser un centro de diversificación del género Oenocarpus. Estos altos niveles de diversidad reportados en el norte de la Amazonía peruana se correlacionan con la baja estacionalidad de la zona (Kristiansen et al. 2011), el cual es un factor determinante en la generación de patrones de diversidad en palmeras (Bjorholm et al. 2005). Por otro lado, la menor diversidad genética fue reportada en San Francisco al norte Colombia (He = 0.54, A = 3.43) y en varias poblaciones del Chocó ecuatoriano, las cuales son periféricas dentro del rango de distribución de la especie. Poblaciones periféricas han sido propuestas como zonas de menor diversidad en relación a poblaciones centrales debido a bajas tasas de flujo génico, su menor tamaño poblacional y mayor aislamiento físico (Hipótesis central-marginal; Eckert et al. 2008). Los valores de endogamia no reflejaron un patrón espacial definido. El mayor nivel de endogamia fue reportado en Villaseca (FIS = 0.37), en el suroccidente de Ecuador, un área de bosque semi-deciduo afectada por la deforestación. Los resultados del presente estudio permiten identificar poblaciones ecuatorianas de O. bataua consideradas prioridades de conservación debido a sus patrones genéticos. Ecuador alberga tres agrupamientos genéticos 133 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 de esta especie (Chocó, ZAH y Napo) los cuales deben ser incluidos en programas de conservación y en el Sistema Nacional de Áreas Protegidas (SNAP). O. bataua presenta una gran variedad de ecotipos en la región del Chocó, creciendo en varios tipos de hábitat. Por ejemplo, poblaciones de O. bataua han sido localizadas en bosques húmedos de Esmeraldas, bosques montanos de Imbabura, y bosques semideciduos en El Oro. A pesar de su diversidad de ecotipos, gran parte de la diversidad genética de esta especie en el Chocó no está protegida por parques nacionales. El PN Cotacahi Cayapas y el PN Mache Chindul ubicados al norte del Chocó, albergan poblaciones silvestres de O. bataua. Sin embargo, las poblaciones ubicadas en las estribaciones andinas occidentales y en el suroccidente del Ecuador se encuentran seriamente amenazadas por la deforestación y degradación del hábitat. Las poblaciones andinas en la zona Amotape-Huancabamba representan un agrupamiento genético particular, potencialmente asociado a un ecotipo que favorece su crecimiento en regiones sobre los 1000 m de altitud. Por lo tanto, estas poblaciones andinas deben ser prioridades de conservación. En particular, las poblaciones cercanas a la ciudad de Zamora están presentes en pequeños remanentes de vegetación, por lo que se encuentran altamente amenazadas por la deforestación. Adicionalmente, en esta zona existen poblaciones que crecen en pastizales sin regeneración natural. El patrón genético de estas poblaciones podría estar protegida dentro del PN Podocarpus (suroriente de Ecuador), sin embargo esto requiere ser evaluado. Las poblaciones pertenecientes al agrupamiento del Napo están mejor representadas dentro del SNAP, en particular en el PN Yasuní, el PN Sumaco NapoGaleras y la RPF Cuyabeno. La caracterización de la diversidad genética y la forma en que ésta se distribuye espacialmente es una herramienta importante para la conservación de los recursos forestales (Eckert et al. 2008). Por lo tanto, la identificación de patrones genéticos de O. bataua en este estudio está enfocada a la conservación de la variabilidad intraespecífica de la especie, la cual debe ser conservada como materia prima para futuros procesos de domesticación y mejoramiento genético. Agradecimientos El presente estudio fue financiado por la Pontificia Universidad Católica del Ecuador y el Proyecto PALMSFP7 (Palms Harvest Impacts in Tropical Forests). Agradecemos al equipo DYNADYV del IRD en Montpellier (Francia) particularmente a Yves Vigouroux, Cédric Mariac, Marie Couderc, Julissa Roncal y Thomas Couvreur; a Betty Millán (Universidad Mayor San Marcos, Perú); a Mónica Moraes (Universidad Mayor San Andrés, Bolivia); y a Gloria Galeano, María José Sanín y Rodrigo Bernal (Universidad Nacional de Colombia) por su ayuda y soporte durante esta investigación. Un agradecimiento especial a Gabriel Rivadeneira y Luis Eduardo López por su colaboración durante la fase de campo en Ecuador. Literatura citada Balick, MJ. 1986. Systematics and economic botany of the Oenocarpus-Jessenia (Palmae) complex. In Advances in Economic Botany. New York, US, New York Botanical Garden. Vol. 3. p. 1-140. Bjorholm, S; Svenning, J-C; Skov, F; Balslev, H. 2005. Environmental and spatial controls of palm Arecaceae) species richness across the Americas. Global Ecology and Biogeography. 14:423–429. Eckert, CG; Samis, KE; Lougheed, SC. 2008. Genetic variation across species’ geographical ranges: the central–marginal hypothesis and beyond. Molecular Ecology. 17:1170–1188. 134 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Evanno, G; Regnaut, S; Goudet, J. 2005. Detecting the number of clusters of individuals using the software structure: a simulation study. Molecular Ecology. 14:2611–2630. Excoffier, L; Laval, G; Schneider, S. 2005. ARLEQUIN version 3.0: an integrated software package for population genetics data analysis. Evolutionary Bioinformatics Online. 1:47–50. Galeano, G; Bernal, RG. 2010. Palmas de Colombia. Guía de campo. Bogotá, CO, Editorial Universidad Nacional de Colombia. 688 p. Guillot, G; Mortier, F; Estoup, A. 2005. Geneland: a computer package for landscape genetics. Molecular Ecology Notes. 5:712–715. Henderson, A. 1995. The palms of the Amazon., Oxford, GB, Oxford University Press. 353 p. Henderson, A; Galeano, G; Bernal, R. 1995. Field Guide to the Palms of America. New Jersey, US, Princeton University –Press. 352 p. Karubian, J; Sork, VL; Roorda, T; Duraes, R; Smith, T. 2010. Destination-based seed dispersal homogenizes genetic structure of a tropical palm. Molecular Ecology. 19:1745–1753. Kreft, H; Jetz, W. 2007. Global patterns and determinants of vascular plant diversity. Proceedings of the National Academy of Sciences. 104(14):5925–5930. Kristiansen, T; Svenning, J-C; Pedersen, D; Eiserhardt, WL; Grández, C; Balslev, H. 2011. Local and regional palm (Arecaceae) species richness patterns and their cross-scale determinants in the western Amazon. Journal of Ecology. 99:1001–1015. Lepsch-Cunha, N; Lund, CA; Hamilton, MB. 2003. Isolation and characterization of nuclear microsatellite loci in the tropical arboreal palm Oenocarpus bacaba (Arecaceae). Molecular Ecology Notes. 3:435–437. Ludeña, B; Chabrillange, N; Aberlenc-Bertossi, F; Adam, H; Tregear, J; Pintaud, J-C. 2011. Phylogenetic utility of the nuclear genes AGAMOUS 1 and PHYTOCHROME B in palms (Arecaceae): an example within Bactridinae. Annals of Botany. 108:1433–1444. Miller, C. 2002. Fruit production of the Ungurahua palm (Oenocarpus bataua subsp. bataua, Arecaceae) in an indigenous managed reserve. Economic Botany. 56(2):165–176. Montúfar, R. 2007. Structure génétique, biochimique, morphologique et écologique de Oenocarpus bataua Mart. (Arecaceae): perspectives pour la valorisation durable d’une ressource forestière néotropicale. Tesis Ph.D., Montpellier, Fr. SupAgro (Centre international d'études supérieures en sciences agronomiques). 138 p. Montúfar, R; Mariac, C; Pham, J; Pintaud, J-C. 2006. Isolation of 23 polymorphic microsatellite loci in the Neotropical palm Oenocarpus bataua Martius (Arecaceae). Molecular Ecology Notes. 7(1):75–78. Montúfar, R; Laffargue, A; Pintaud, J-C; Hamon, S; Avallone, S; Dussert, S. 2010. Oenocarpus bataua Mart. (Arecaceae): Rediscovering a Source of High Oleic Vegetable Oil from Amazonia. Journal of the American Oil Chemists' Society. 87(2):167–172. Montúfar, R; Brokamp, G. 2011. Palmeras aceiteras del Ecuador: estado del arte en la investigación de nuevos recursos oleaginosos provenientes del bosque tropical. Revista ecuatoriana de Medicina y Ciencias Biológicas. 32(1,2):93–118. 135 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Núñez-Avellaneda, LA; Rojas-Robles, R. 2008. Biología reproductiva y ecología de la polinización de la palma milpesos Oenocarpus bataua en los andes colombianos. Caldasia. 30(1):101–125. Pintaud, J-C; Galeano, G; Balslev, H; Bernal, R; Borchsenius, F; Ferreira, E; Granville, J-J de; Mejía, K; Millán, B; Moraes, M; Noblick, L; Stauffer, FW; Kahn, F. 2008. Las palmeras de América del Sur: diversidad, distribución e historia evolutiva. Revista Peruana de Biología. 15(supl. 1):7–29. Pitman, NCA. 2000. A large-scale inventory of two Amazonian tree communities. Tesis Ph.D., Durham, GB. Duke University. 220 p. Pritchard, JK; Stephens, M; Donnelly, P. 2000. Inference of population structure using multilocus genotype data. Genetics. 155:945–959. Steege, H ter; Pitman, NC; ... ; Silman, N. 2013. Hyperdominance in the Amazonian Tree Flora. Science. 342(6156):1243092. Trénel, P; Hansen, M; Normand, S; Borchsenius, F. 2008. Landscape genetics, historical isolation and crossAndean gene flow in the wax palm Ceroxylon equinulatum (Arecaceae). Molecular Ecology. 17:3528–3540. Vormisto, J; Svenning, J-C; Hall, P; Balslev, H. 2004. Diversity and dominance in palm (Arecaceae) communities in terra firme forests in the western Amazon basin. Journal of Ecology. 92:577–588. Figuras y tablas Fig. 1 Mapa de Sudamérica mostrando la localización de las 591 muestras de Oenocarpus bataua var. bataua utilizadas en este estudio. Las circunferencias grandes representan localidades con n>15. Elevaciones sobre los 1000 m de altitud se muestran en gris. 136 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Fig. 2 Cinco mapas representando los agrupamiento genéticos encontrados dentro de Oenocarpus bataua var. bataua usando 591 muestras. Estos fueron identificados con un análisis bayesiano-espacial realizado en Geneland (Guillot et al. 2005) usando probabilidades de pertenencia a cada agrupamiento (K=5) como fue identificado en Structure (Pritchard et al. 2000). Cada punto representa una localidad muestreada, mientras que las líneas representan probabilidades de pertenencia a cada agrupamiento. Tabla 1 Valores de diversidad e índices pareados de diferenciación genética (FST) obtenidos en Arlequin (Excoffier et al. 2005) para cada uno de los cinco agrupamiento genéticos de Oenocarpus bataua var. bataua identificados en Structure (Pritchard et al. 2000). Abreviaturas: VRM=valle del río Magdalena, CH=Chocó, ZAH=zona Amotape-Huancabamba, N=Napo, I=Inambari. n=sampling, A=número promedio de alelos por locus, He=heterocigosidad esperada, Ho=heterocigosidad observada, FIS=coeficiente de endogamia. ** = p<0.01 FST Agrupamiento VRM CH ZAH N I n A He Ho FIS VRM CH ZAH N I 25 118 107 186 155 3.43 12 12.57 16.57 16.43 0.54 0.75 0.77 0.83 0.79 0.54 0.69 0.71 0.73 0.71 -0.002** 0.071** 0.065** 0.086** 0.095** 0 0.32** 0.30** 0.26** 0.27** 0 0.12** 0.09** 0.12** 0 0.05** 0.09** 0 0.04** 0 Tabla 2 Valores de diversidad genética para cada una de las 19 poblaciones de Oenocarpus bataua var. bataua obtenidos en Arlequin (Excoffier et al. 2005). Abreviaturas: VRM=valle del río Magdalena, CH=Chocó, ZAH=zona Amotape-Huancabamba, N=Napo, I=Inambari. n=sampling, A=número promedio de alelos por locus, He=heterocigosidad esperada, Ho=heterocigosidad observada, FIS=coeficiente de endogamia. ** = p<0.01 Población San Francisco Esmeraldas Agrupamiento n VRM 25 CH 26 137 A 3.43 6.43 He 0.54 0.70 Ho 0.54 0.81 FIS -0.002 -0.158 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 El Chontal Bilsa Villaseca Pachicusa Zamora Rioja Yasuní Pantoja Chiriap Intuto Jenaro Herrera Pucallpa Ahuaytía Shuaro Iñapari Tambopata San Buenaventura CH CH CH ZAH ZAH ZAH N N N N N I I I I I N 32 31 19 19 57 18 30 26 30 32 30 43 32 15 30 35 17 138 6.86 6.86 6.6 7 8.29 8.14 8.43 8.43 8.71 11.71 11.86 10.71 8.14 7.14 10.14 9.43 6 0.70 0.73 0.77 0.73 0.71 0.76 0.75 0.74 0.72 0.86 0.81 0.76 0.67 0.71 0.77 0.72 0.75 0.75 0.84 0.47 0.70 0.74 0.61 0.60 0.65 0.88 0.82 0.86 0.75 0.66 0.53 0.82 0.68 0.60 -0.074 -0.172 0.377** 0.031 -0.046 0.157** 0.17** 0.038 -0.314 0.034 -0.066 0.007 0.01 0.242** -0.064 0.045 0.23** PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 EVALUACIÓN DEL ESTADO DE CONSERVACIÓN Y CARACTERIZACIÓN POBLACIONAL DE DOS ESPECIES FORESTALES PRIORIZADAS EN LA MICROCUENCA DEL RÍO CHIMBORAZO. María Mercedes Pavón Naveda, Jorge Grijalva Olmedo, Raúl Ramos, Paulo Barrera Instituto Nacional Autónomo de Investigaciones Agropecuarias. Estación Experimental Santa Catalina. Programa Nacional de Forestería. Panamericana Sur Km1, sector Cutuglahua, Quito. [email protected] INTRODUCCIÓN El análisis del estado de conservación de los Recursos Genéticos Forestales en el Mundo fue acogido por la 9ª reunión de la Conferencia de las Partes (COP, 2008) del Convenio sobre Diversidad Biológica. Paralelamente, la Estrategia Regional de Biodiversidad para los Países del Trópico Andino, adoptada por la Decisión 523 de la Comunidad Andina (julio de 2002), destaca también la importancia de adelantar programas y proyectos de investigación y desarrollo relacionados con iniciativas de conservación y manejo adecuado de la biodiversidad. Ese marco global alienta a las políticas sectoriales y los proyectos de desarrollo con impacto subregional, a que integren en todo su ciclo elementos específicos de conservación de biodiversidad y desarrollo sustentable. En respuesta a esas prioridades de orden global, el Ecuador ha respondido mediante la Política Forestal del Ecuador la cual promovió la elaboración de la Estrategia de Desarrollo Forestal sustentable para el Ecuador actualmente en discusión (MAE, 2011). Para una adecuada implementación, es valioso la Estrategia Nacional para la Conservación de la Biodiversidad 2001-2010, el Plan Nacional de Forestación y Reforestación (MAGAP, 2006) y la normativa asociada a bosques, promovida por el MAE. El fortalecimiento de las administraciones, la expedición de leyes y su aplicación y la armonización sectorial son los desafíos de las autoridades ambientales. Los bosques andinos son importantes sistemas sustentadores de vida. De ellos dependen numerosas comunidades humanas, pues les proveen de agua (para riego y consumo humano), madera, plantas medicinales y animales silvestres. Son espacios de educación, recreación y albergues de sitios sagrados. La provincia de Chimborazo no es ajena a este complejo panorama. Las cifras a nivel provincial muestran que en las microcuencas durante los últimos 30 años, la frontera agrícola aumentó en 160 mil hectáreas en detrimento de áreas frágiles de páramo (FAO 1995). De hecho, en la microcuenca del río Chimborazo predomina la actividad agropecuaria; el 85% del agua se usa para agricultura, y más de las dos terceras partes se desperdician y contaminan (Municipio de Riobamba, citado por Grijalva, 2006). Por otro lado, como efecto del calentamiento global, se evidencia la desglaciación del nevado Chimborazo que pierde 0,5 m de hielo/año, lo que genera un cambio en el régimen hidrológico de las cuencas y micro-cuencas (Martínez, 2004). 139 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 JUSTIFICACIÓN A pesar de la importancia señalada sobre recursos forestales, el conocimiento de RGFs aún es precario e insuficiente; son escasos los estudios y las instituciones que realizan actividades para su protección, y la disponibilidad actual de información específica sobre la situación y tendencias de los recursos genéticos forestales. El proyecto “Conservación y Uso Sostenible de Recursos Genéticos Forestales en áreas críticas de bosques húmedos y secos de los Andes y Amazonía”, que conduce el Programa Nacional de Forestería busca proteger la variabilidad genética remanente de especies forestales prioritarias vulnerables para contribuir a la conservación de los sitios altoandinos, la protección y recuperación de cuencas hidrográficas andinas. Esta investigación es una iniciativa pionera en el país, cuyo propósito es generar datos y nuevo conocimiento sobre el estado de conservación y la vulnerabilidad de RGF’s de especies forestales priorizadas en la microcuenca del río Chimborazo. OBJETIVOS Objetivo General a. Determinar el estado de conservación de dos especies a ser priorizadas en la microcuenca del Río Chimborazo. Objetivos específicos a. Priorizar con enfoque participativo dos especies forestales en la microcuenca del río Chimborazo. b. Evaluar el estado de conservación de dos especies forestales priorizadas en el área de estudio. c. Caracterizar y recolectar la base poblacional in situ de las dos especies forestales priorizadas en el estudio. HIPOTESIS Ho = Las especies forestales priorizadas se encuentran amenazadas. Ha = Las especies forestales priorizadas no se encuentran amenazadas. MATERIALES Y MÉTODOS Localización La investigación se realizará en la micro-cuenca del río Chimborazo, fuente de la sub-cuenca del río Chibunga, afluente del río Chambo. Se localiza al noroccidente de la provincia de Chimborazo, cantón Riobamba, parroquia San Juan, geográficamente localizada entre las coordenadas UTM de los puntos extremos 9836 000 y 9820 000 Norte y, 732 000 y 747 000 Este. El rango altitudinal va desde los 3240 hasta 140 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 los 6280 metros (Programa de Forestería, 2008). El 50 % de las comunidades pertenecen a la UOCIC y UCASAJ. La micro-cuenca presenta precipitaciones de hasta 1100 mm año, que se extiende desde noviembre hasta mayo, el rango de humedad relativa es de 75 a 80 %. La temperatura media es de 10 ºC con variaciones diarias principalmente en los meses de noviembre y diciembre, lo que determina la presencia de heladas, principal factor determinante de la pérdida de producción de cultivos (PDA-UOCIC 2006; Programa Nacional de Forestería, 2008). Priorizar con enfoque participativo dos especies forestales en la micro-cuenca del río Chimborazo. Diagnóstico rural participativo para seleccionar las especies Se revisará la información generada por el INIAP, donde se presenta una lista de especies forestales preseleccionadas por las propias comunidades dos años atrás. La priorización de las especies se realizará a través de votación individual utilizando cinco stikers de diferente color. El color será ponderado como se describe a continuación: Sticker de color amarillo = especie de primera importancia con una ponderación de 0,45 Sticker de color azul = especie de segunda importancia con una ponderación de 0,25 Sticker de color rojo = especie de tercera importancia con una ponderación de 0,15 Sticker de color verde= especie de cuarta importancia con una ponderación de 0,10 Sticker de color rosado = especie de quinta importancia con una ponderación de 0,05 Se presentará una tercera matriz a fin de que cada participante coloque en orden de importancia las especies seleccionadas e identifiquen los usos de las mismas. Finalmente se analizarán las frecuencias de los usos de las especies priorizadas, eligiendo aquellas dos especies que obtengan las mayores frecuencias. Evaluar el estado de conservación de dos especies forestales priorizadas en el área de estudio.Mediante el uso de la información geográfica de la microcuenca del río Chimborazo disponible, se identificarán los tres tipos de uso del suelo predominante: Área de Remante de bosque natural6 Área de Bosque plantado7 Área de sistemas integrados8 Categorías de conservación a utilizarse 6 Puede definirse como la transformación de un bosque continuo en muchas unidades más pequeñas y aisladas entre sí, cuya extensión resultante es mucho menor que la del bosque original.(www.buenastarreas/fragmentaciondebosques) 7 Son poblaciones arbóreas sembradas o plantadas bajo la supervisión e intervención del hombre en el proceso de forestación y reforestación, sea con una o varias especies; por lo general tienen una misma edad, altura y similar densidad entre individuos. 8Son sistemas agroforestales con remanentes de vegetación arbustiva. 141 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Para el presente estudio se utilizarán las categorías definidas por la UICN9 (2001 versión 3.1), que se detallan a continuación Criterios a utilizarse para definir la categoría de conservación En la presente investigación se ha definido utilizar solamente los criterios B y D, por considerarlos suficientes para el cumplimiento de objetivos del estudio, dado que son los más adecuados debido a su menor complejidad al momento del análisis, las características del terreno y la información potencial disponible. Se aplicará el criterio D únicamente en aquellos casos donde no sea posible la aplicación del criterio B. El Criterio B Y D se refiere a: Criterio B. Distribución geográfica en la forma B1 (extensión de la presencia) O B2 (área de ocupación) o ambas. Criterio D. Se estima que el tamaño de la población que es menor de 250 individuos maduros. (UICN, 2001). Recopilación y análisis de información secundaria Se compilará la mayor información disponible sobre las dos especies priorizadas en la base de datos de los Herbarios del país. Cartografía base, en este grupo se clasifica a elementos como son las curvas de nivel, ríos, poblados, vías de acceso, cotas, principalmente. Unidades de Muestreo A través de la cartografía base y las fotos satelitales, se determinarán los remanentes ubicados en las zonas ya definidas anteriormente. Se codificaran los remanentes (R1) con su respectivo hectareaje, así consecutivamente según el número de remanentes que se encuentren en las zonas. En cada remanente se establecerán unidades de muestreo dependiendo del área separadas 50 m. En cada unidad se establecerá el uso de transectos, la forma del transecto será en zigzag, de esa forma se homogenizará el lugar muestreado (Cerón, 1993). Las variables a ser analizadas en este estudio son las siguientes: La evaluación del estado de conservación en remanentes utiliza como indicador El Índice de Vegetación Remanente La frecuencia. Estimación de la disminución inferida del hábitat. Estimación del tamaño de la población: Se delimitará el área de extensión potencial, descrito en el cálculo del área de extensión (Criterio B), conteo de individuos maduros en parcelas de muestreo (40x60 m), se establecerán 3 parcelas por estrato de vegetación, se contarán los individuos encontrados por hectárea. (Mostacedo, 2000). 9 Categoría o unidad formal taxonómica a cualquier nivel en una clasificación (familia, género, especie, etc.) 142 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Caracterización y recolección de la base poblacional in situ de las especies forestales priorizadas. Se considerarán los mismos estratos utilizados anteriormente, al igual que el análisis de información secundaria. (Fernández 2009). Elaborado: María Mercedes Pavón Naveda RESULTADOS Priorizar con enfoque participativo dos especies forestales en la microcuenca del río Chimborazo. Desarrollo del taller. Se realizó el taller de Priorización con enfoque participativo de dos especies forestales con representantes de las comunidades que conforman la microcuenca del río Chimborazo se obtuvo un registro de 41 asistentes (incluyendo al equipo técnico del PNF10) , en la parroquia San Juan – Cantón Riobamba, Provincia de Chimborazo, en el que se seleccionaron las dos especies forestales para su conservación estas fueron Polylepis sp, con una frecuencia de 29 y su puntaje de ponderación de 12,25 y para Buddleja incana 21 de frecuencia y ponderación de 4,75, con la colaboración de los participantes de identificaron los usos las especies priorizadas. 10 Programa Nacional de Forestería 143 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Elaborado: María Mercedes Pavón Naveda BIBLIOGRAFIA Cerón, C 1993. Manual de Botánica Ecuatoriana. U. Central del Ecuador. Quito. Fernandez Jaramillo, AC. 2009. Elaboración de un Plan de Manejo y Conservación de Pinus tropicalis Morelet, en fase a Criterios de Ecofisiología de la especie, en alturas de pizarra, Viñales, en la provincia de Pinar del Río, Cuba. Tesis Lic Ing. For. 109-114p. FAO (Organización de las Naciones Unidad para la Agricultura y la Alimentación, IT). 1995. Pequeñas industrias forestales: carpintería comunal en San Juan de Chimborazo. Quito, EC. Proyecto FAO-Holanda. Desarrollo Forestal Participativo en los Andes. p 131, 217 p. MAE, 2011. Estrategia Nacional para la Conservación de la Biodiversidad. Disponible en: www.ambiente.gob.ec/.../TDR-Asesoría-en-Productividad-Sostenible.pdf Martínez, E. 2004. Se precipita el retroceso de los glaciares andinos. Revista: tendencias científicas. Consultado 15 ene. 2008. Disponible en: http://dave.madteam.net/blog001002/art_23/ Mostacedo, Bonifacio; Fredericksen, Todd S. 2000. Manual de Métodos Básicos de Muestreo y Análisis en Ecología Vegetal. Santa Cruz, Bolivia. Editorial El País. 35p, 43p. PDA-UOCIC (Programa de Desarrollo de Área - Unión de Organizaciones Campesinas Indígenas intercomunales Chimborazo Rey de los Andes, EC). 2006. Diagnóstico participativo comunitario. San Juan, Chimborazo, EC. Sinchiguano y equipo de facilitación. Visión Mundial. 193 p. Programa Nacional de Forestería. 2008. Caracterización de la microcuenca del río Chimborazo. Proyecto Iniciativa interinstitucional de Investigación/Desarrollo Agroforestal Participativo para la protección y manejo sostenible de la microcuenca del río Chimborazo. Riobamba, EC. INIAP. 44 p. UICN (Unión Mundial para la Naturaleza). 2001 Categorías y Criterios de la Lista Roja de la UICN Versión 3.1 Preparado por la Comisión de Supervivencia de Especies UICN Aprobado en la 51° Reunión del Consejo de la UICN Gland, Suiza 9 Febrero 2000 144 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 COLECTA Y CARACTERIZACIÓN MORFOLÓGICA IN SITU DE CAPULÍ (Prunus serotina Ehrh) DEL BANCO NACIONAL DE GERMOPLASMA DEL INIAPECUADOR Juan Chucuri M., Álvaro Monteros A., Edwin Borja B., César Tapia B. Instituto Nacional Autónomo de Investigaciones Agropecuarias INIAP, Apartado Postal N° 17-01-340. Quito – Ecuador [email protected] Introducción El capulí (Prunus serotina Ehrh.), es un árbol nativo de México; sin embargo, ha sido utilizado en Ecuador desde tiempos precolombinos por agricultores ubicados en la región sierra (León, 2000). En el Ecuador, no existen plantaciones extensivas, pero se encuentran plantas dispersas en todas las provincias de la sierra Ecuatoriana. Cabe resaltar que la provincia de Cotopaxi y Tungurahua poseen el mayor número de árboles de capulí (Flores, 2008). La madera es aprovechable en la construcción rural, decoración de interiores, carpintería en general; se caracteriza por tener un color rojizo brillante. La facilidad de labrado es lo que permite hacer esculturas y decoraciones de alto valor estético, aparte tiene uso como leña y carbón en los andes Ecuatorianos (Flores, 2008). El INIAP promueve la conservación, manejo y uso de la agrobiodiversidad del país. El capulí ha sido identificado como una especie poco estudiada con potencial para uso y conservación. El capulí no sólo tiene importancia por su riqueza genética, sino también debido a sus propiedades nutricionales y tolerancia a condiciones adversas. Problema En la serranía ecuatoriana se observa variabilidad genética de capulí, sin embargo no se ha realizado ningún tipo de estudios enfocados a la descripción de esta diversidad. Actualmente los agricultores tumban los árboles antiguos porque causan sombra a los cultivos. Esta tumba de árboles puede provocar una erosión genética de esta especie forestal. Objetivos 1. General Colectar y caracterizar morfológicamente in situ la variabilidad genética de capulí en la sierra ecuatoriana 2. Específicos 2.1 Colectar la variabilidad genética de capulí existente en la región interandina del Ecuador para su conservación en el banco de germoplasma de INIAP. 145 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 2.2 Caracterizar in situ las accesiones identificadas como representativas de las provincias mediante descriptores morfológicos estandarizados. Método de estudio Colecta de fruto (semilla) Se identificaron accesiones en base a datos pasaporte registrados en años anteriores por el banco de germoplasma de INIAP. Se colectaron frutos de capulí en fundas de papel, se colectaron al menos 2000 semillas por accesión, identificados de acuerdo al código de colecta. Estas semillas se trasladaron hasta el laboratorio de semillas del Banco de germoplasma del INIAP- Ecuador, para su procesamiento y conservación. Caracterización morfológica insitu Para la caracterización morfológica in situ se utilizaron 32 descriptores cuantitativos (15) y cualitativos (17), que determinan la morfología de la colección de capulí en base a descriptores propuestos por Internacional Board For Plant Genetic Resources (IBGRI) (2002), y la: Unión Internacional Para la Protección de las Obtenciones Vegetales, (UPOV) (2002) Análisis estadístico El presente trabajo investigativo se empleó el análisis multivariado utilizando Infostat (2008) método de Ward y distancia Euclediana. Además, se utilizó el DIVA GIS para identificar los puntos de colecta. Resultados En este estudio se identificaron 168 accesiones de las diez provincias de la región andina, y al realizar el análisis de agrupamiento utilizando el método de Ward y el distancia Euclidiana, se obtuvieron tres grupos (Figura 1). 146 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figura 1. el agrupamiento de 168 cesiones de capulí obtuvieron tres grupos Dendrograma generado mediante Ward y distancia Euclideana de (Prunus serotina Ehrh.), y se representativos de la colección. 147 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 El análisis de 15 descriptores cuantitativos se obtuvieron los siguientes resultados (Cuadro 1): El (G1), presentó mayor número de ramas secundarias por árbol (11,00), longitud de inflorescencia (95,35 cm) y número de inflorescencias por rama terciaria (13,00). El (G2), se observó el mayor promedio (0,43 m) entre grupos para diámetro de tallo (DAP) y altura total del árbol 26,50 m. El (G3), presentó el mayor promedio de los grupos para diámetro de la flor (11,00 mm), longitud de sépalo. (5,97 mm) y ancho de sépalo (4,93 mm). El grupo (G1) incluye 71 accesiones, el grupo 2, (G2) 57 y el grupo (G3) 40 accesiones. Los datos de las variables cualitativas son resumidos en el Cuadro 2. 148 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Cuadro 1. Resumen de las variables cuantitativas para cada uno de los tres grupos identificados GRUPO 1 DESCRIPTOR Diámetro del tallo (cm). Altura total (m). Altura de inserción de ramas secundarias (m). Número de ramas secundarias. Longitud del pecíolo de la hoja (mm). Diámetro del pecíolo de la hoja (mm). Longitud de la hoja (mm). Ancho de la hoja (mm). Longitud de inflorescencia (excluida el Pedúnculo) (mm). Número de inflorescencia por rama terciaria. Longitud del pedúnculo floral (mm). Diámetro de la flor (mm). Longitud de sépalo (mm). Ancho de sépalo, (ubicado en la parte más ancha) (mm). Longitud de la flor (mm). Valor Valor mínimo máximo 0,12 16,3 0,28 GRUPO 2 Valor Valor mínimo máximo PROMEDIO D.E. 0,95 0,39 0,21 0,12 26,3 20,57 2,59 18,45 3,15 1,43 0,57 0,19 4 GRUPO 3 Valor Valor mínimo máximo 0,25 0,11 3,72 15,52 1,77 0,75 0,1 PROMEDIO D.E. 1 0,43 32,23 26,47 PROMEDIO D.E. 0,64 0,3 0,14 30,3 21,43 3,5 2,25 1,2 0,58 3 41 11,52 6,35 1 25 9,3 4,63 1 17 7,98 3,48 8,52 19,89 14,59 2,49 9,38 24,36 16,96 3,14 9,38 21,98 16,99 2,82 0,64 2,11 1,09 0,27 0,35 14,54 1,42 2,33 0,36 1,9 0,99 0,26 59,14 212,21 101,58 20,6 18,3 133,1 100,34 22,08 46,23 126,9 101,57 18,02 21,03 52,64 33,63 5,24 20,43 63,21 35,53 7,39 29,51 96,95 40,95 14,64 42,75 155,33 95,35 24,35 11,29 113,35 78,95 19,01 43,67 120,51 89,05 17,5 2 50 12,8 7,27 2 23 9,33 4,03 3 27 9,75 4,61 1,87 6,22 3,83 0,96 2,55 8,14 4,5 1,32 3,27 17,7 5,73 2,42 2,29 11 8,11 2,07 4,41 14,2 9,86 1,68 3,27 34,89 11 6,29 2,04 6,08 3,78 0,67 3,02 7,95 4,44 0,85 3,26 11,17 5,97 2,25 2,27 7,54 3,69 0,86 2,67 6,39 4,12 0,73 2,86 11,02 4,93 1,79 3,36 10,12 6,39 1,29 5,25 21,39 7,46 2,34 2,34 34,5 7,91 4,58 149 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Cuadro 2. Resumen de los estados de descriptores cualitativos con mayor frecuencia par a 17 variables cualitativas en la caracterización de capulí (Prunus serotina) por grupo identificado con el análisis multivariado. G1 (%) G2 (%) G3 (%) 4 (0,38) 1 (0,70) 1 (0,73) 3 (0,45) 2 (0,54) 2 (0,53) Forma de las ramas secundarias 1. Erecto, 2. Semierecto, 3.Extendido, 4. Colgante: 4 (0,45) 1 (0,42) 3 (0,48) Presencia de yemas vegetativas en las ramas secundarias 1. No, 2. Si 1 (1,00) 1 (1,00) 1(1,00) Forma de la hoja 1. Elíptico, 2. Redondeado 1 (1,00) 1 (1,00) 1(1,00) Base de la hoja 1. Aguda, 2. Obtusa, 3. Redonda: 2 (0,94) 1(0,67) 1 (0,68) Ápice de la hoja 1. Acuminado, 2. Agudo 1 (1,00) 1 (1,00) 1 (1,00) Color de las hojas 1. Verde, 2. Verde Claro, 3. Verde Oscuro: 1 (0,56) 1 (0,51) 1 (0,40) Presencia de pubescencia en el envés de la hoja 0. No, 1. Si: 1 (1,00) 1 (1,00) 1 (1,00) 3 (0,66) 3 (0,98) 3 (0,98) Carácter Hábito de crecimiento 1.Erguido, 2. Semierguido 3. Extendido, 4. Colgante: Forma de la copa: 1. Aguda, 2. Obtusa, 3. Redondeada: Inserción del borde 1. Crenadas, 2. Crenadas aserrada, 3. Aserrada: Forma de la inflorescencia 1. Racimo, 2. Espiga, 3. Espiguilla Tipo de flor 1. Sencilla (5 pétalos), 2. Semidoble (2 o 3 circulo de pétalos: máximo de 15), 3. Doble (más de círculos de pétalos): Color del pedúnculo floral Verde claro Color de la flor Blanco Orientación de los sépalos 1. Replegados (ápice del sépalo con dirección al cáliz), 2. Horizontal (ápice del sépalo con dirección contraria al cáliz): Posición de las anteras 1. Ausente (anteras que no están cubiertas por la capucha), 2. Normal (anteras cubiertas por la capucha): Hábito de floración 1. Florece una sola vez, 2. Florece dos veces, 3. .Floración continua: 150 1 (0,82) 2 (0,63) 2 (0,63) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) 1 (1,00) PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Conclusiones La caracterización morfológica de las 168 accesiones, permitió iniciar con el desarrollo de conocimiento de la variabilidad de capulí en el campo y su estrecha relación genética con cada una de las entradas dando como resultado preliminar la conformación de 3 grupos representativos de la colección. Los grupos no tienen clara relación con la distribución geográfica por provincia. Sin embargo llama la atención que el grupo 1 tiene el 93% de las accesiones de Cotopaxi y el 100% de accesiones de Azuay están en el grupo 2. Todas las accesiones identificadas (168) presentan floración una sola vez al año entre septiembre y octubre y fructifican de enero hasta marzo. Este estudio ha permitido realizar una primera descripción de la diversidad de capulí existente en los andes de Ecuador y se prevé complementar el estudio de diversidad genéticautilizando 5 marcadores microsatélites en la colección. Estos resultados en conjunto permitirán determinar árboles claves que deben ser conservados in situ. Durante las colectas comparando la base de datos del banco con los materiales encontrados se determina que hay algunos materiales que han sido eliminados de las chacras, lo que daría indicios de una potencial erosión genética de materiales. Las colectas han permitido colectar, procesar y conservar semillas de capulí en el banco de germoplasma de INIAP para conservación ex situ. Bibliografía Di Rienzo J.A., Casanoves F., Balzarini M.G., Gonzalez L., Tablada M., Robledo C.W. (2008). InfoStat, versión 2008, Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Hijmans, Robert J., Luigi Guarino, Coen Bussink, Barrantes I., Rojas E, 2002. DIVA- GIS, versión 2. Sistema de información Geográfica para el análisis de Datos de Biodiversidad. Manual. International Potato Center, Lima, Perú. León J. 2000. Fundamentos Botánicos de los Cultivos Tropicales. Del Instituto Interamericano de Cooperación para la Agricultura (IICA). Edición tercera revisada y aumentada. San José. Costa Rica. p. 183. 151 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 GESTIÓN FORESTAL SOSTENIBLE Eduardo Colina COMPORTAMIENTO AGRONÓMICO DEL LAUREL (Cordia alliodora), BAJO DOS SISTEMAS DE PASTOREO EN LA ZONA DE FEBRES-CORDERO, PROVINCIA DE LOS RÍOS Franklin Sigcha EFECTOS DE LA EXCLUSIÓN DE GRANDES HERBÍVOROS SOBRE LA VEGETACIÓN Y EL SUELO, EN MINAS DE CARBÓN RESTAURADAS EN EL NORTE DE PALENCIA Janice Jácome ESTRUCTURA POBLACIONAL, PRODUCTIVIDAD Y FENOLOGÍA FLORAL DE LA TAGUA (Phytelephas aequatorialis) EN LA COSTA ECUATORIANA Jorge Luis Cué García PROPUESTA METODOLÓGICA DE CRITERIOS E INDICADORES DEL MANEJO FORESTAL SOSTENIBLE EN LAS CONDICIONES DE CUBA Roy Vera Vélez EFECTO DE VARIABLES CLIMÁTICAS Y LITOLÓGICAS SOBRE EL CRECIMIENTO EN ALTURA DOMINANTE EN MASAS DE Pinus nigra Arn Tomislav Sapic ADS80 AND MODIS BASED GIS AND REMOTE SENSING APPLICATIONS IN FOREST MANAGEMENT 152 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Gabriela Jami VIABILIDAD TÉCNICA, PRODUCTIVA, COMERCIAL Y AMBIENTAL DEL JABÓN ARTESANAL “Yakumama” ELABORADO CON PRODUCTOS FORESTALES NO MADERABLES DE LOS BOSQUES DE LA COMUNIDAD KICHWA RÍO BLANCO Angélica Cárdenas VALIDACIÓN DE UN PROTOCOLO PARA LA PROPAGACIÓN MASIVA IN VITRO DE Paulownia elongata, Paulownia fortunei Y UN HÍBRIDO (Paulownia fortunei x Paulownia elongata) BAJO SISTEMAS DE PROPAGACIÓN CONVENCIONAL E INMERSIÓN TEMPORAL Lucrecia Maldonado ZONIFICACIÓN AGROECOLÓGICA DEL CULTIVO DE CAUCHO Hevea brasiliensis EN EL ECUADOR EN BASE A LA INCIDENCIA DEL MAL SURAMERICANO DE LAS HOJAS José Riofrío ANÁLISIS DE LOS PATRONES DE CRECIMIENTO RADIAL DE MASAS DE Pinus pinaster Ait., EN RESPUESTA A LA VARIABILIDAD CLIMÁTICA EN EL CENTRO-SUR DE ESPAÑA 153 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 COMPORTAMIENTO AGRONÓMICO DEL LAUREL (Cordia alliodora), BAJO DOS SISTEMAS DE PASTOREO EN LA ZONA DE FEBRES-CORDERO, PROVINCIA DE LOS RÍOS Colina, Eduardo, Troya Germán, Sánchez Hugo, Castro Carlos. Universidad Técnica de Babahoyo, Agrícola Macondo. [email protected] Introducción El conocimiento de la vegetación del bosque premontano tropical, ha tenido un aumento en los últimos años en el país, esto ha hecho que se puedan encontrar especies adaptadas que sirvan con fines diferente a los conocidos durante mucho tiempo, esto ha hecho que los Andes tropicales sean conocidos como centros de diversidad mundial. Sin embargo, los estudios que documentan cuantitativamente la estructura y diversidad en los bosques subtropicales de Ecuador y específicamente la provincia de Los Ríos son todavía muy escasos (SNAP, 2010). Los bosques tropicales mantienen alrededor de 20 millones de especies de plantas y animales. Esta variedad de formas de vida se llama diversidad biológica o biodiversidad (Brack, 2004). Lastimosamente, la tala y la deforestación de los bosques naturales, originada por la expansión de las tierras agrícolas y ganaderas, han provocado la reducción de los bosques y la pérdida de muchas especies de la fauna y flora silvestre. Los bosques subtropicales húmedos de Ecuador cubren aproximadamente 80.000 km2 que corresponde al 25.7 % del territorio nacional y cerca de la mitad de las especies de flora y fauna - incluyendo la mayoría de las especies endémicas - se encuentran dentro áreas protegidas. En la zona de FebresCordero se estimaba la existencia entre 200 y 600 especies de plantas (Myers et al., 2000). El interés por los sistemas silvopastoriles ha generado la necesidad de la investigación del efecto de un componente vegetal en el otro y el beneficio de las asociaciones gramínea – leguminosa u otra especie arbórea en particular. Los sistemas de producción silvopastoriles se definen como una serie de sistemas y tecnologías para el uso de la tierra en las que se combinan árboles con pastos, en función del tiempo y el espacio para incrementar y optimizar la producción en forma sostenida. Estos sistemas pueden contribuir a solucionar problemas en el uso de los recursos naturales debido a las funciones biológicas y socioeconómicas que cumplen. La cobertura arbórea en pasturas contribuye a mejorar la productividad animal por medio de la reducción del estrés calórico a generar sombra y refracción de luz directa, además de sus beneficios hacia la conservación del suelo y la biodiversidad. Los árboles en las pasturas, contribuyen a mejorar la conservación de la fauna silvestre (Harvey y Haber, 1999).Varios estudios realizados en Latinoamérica con aves y mariposas, mostraron que las sistemas silvopastoriles mejorados, con alta densidad de árboles, presentaron una mayor riqueza de especies que las pasturas con baja densidad (menos de 30 árboles/ha) y las pasturas en mono-cultivo. 154 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Un estudio realizado en la zona seca de Cañas, Costa Rica muestran que el ganado pastoreado en potreros con muchos árboles (27 % de cobertura) ganó, en un período de 3 meses, en promedio 10,4 kilogramos más que los animales que estaban pastoreando en potreros con pocos árboles (7 % de cobertura) (Restrepo, 2002). Otro estudio, realizado en Matiguas, Nicaragua, determinó que las vacas pastoreando en potreros con muchos árboles (22 a 30 % de cobertura) produjeron, en promedio 0,9 litros/día más que las vacas pastoreando en potreros con pocos árboles (0 a 7 % de cobertura) (Betancourt et al, 2003). Muchas veces, los ganaderos sólo valoran el efecto negativo de la sombra sobre la reducción de pasto y no toman en cuenta el efecto positivo de la sombra sobre los incrementos en la producción animal. En la actualidad para el manejo tradicional de las fincas existen árboles dispersos o agrupados provenientes de la regeneración natural, inducida por el hombre y/o remanentes de la vegetación original. La mayoría de estudios se ha enfocado en la caracterización estructural, conocimiento local y producción de frutos (Esquivel et al., 2003, Souza, 2002). El objetivo del presente estudio fue conocer el efecto de la cobertura arbórea sobre la disponibilidad de pasto Brachiaria brizantha. El presente estudio tuvo como objetivo evaluar el comportamiento agronómico del laurel (Cordia alliodora) en dos sistemas de pastoreo. Los objetivos planteados para el efecto fueron 1) Realizar la caracterización morfológica de la especie en ambos sistemas de manejo, 2) Determinar el efecto de cobertura del pasto quinea (Panicum maximun) sobre la cobertura arbórea del laurel (Cordia alliodora) y 3) Realizar un análisis cuantitativo de la composición de especies encontradas en el área de estudio. Procedimiento Experimental El estudio se realizó en la parroquia Febres-Cordero del Cantón Babahoyo, provincia de Los Ríos, Ecuador (692891 sur y 9781978 oeste). Pertenece a la zona de vida de Bosque Sub Húmedo Tropical, con una altitud que varía entre 50 y 1000 m, temperaturas medias de 23 ºC y precipitaciones anuales entre 1.500 a 1900 mm. El presente estudio se llevó a cabo tanto en la época lluviosa, como en la seca, aunque el crecimiento del pasto se afectado (ausencia total de lluvias). Se dividieron los potreros en tres zonas una externa o cercas, una sección media o perimetral y una sección central cerca del rio que pasa en la zona media del lote. Se seleccionaron al azar 4 potreros (40 ha) con pasto Guinea (P. maximun) en buena condición y cobertura de suelo. Se realizó un inventario de los árboles con diámetro a la altura del pecho (DAP, tomado con metro flexible) de 10 cm en cada potrero y la altura efectiva del dosel del árbol (tomado con 155 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 un altímetro). Se identificó la especie de cada individuo y se estimó el área de copa mediante la medición perpendicular del diámetro mayor y el menor. Se determinó la disponibilidad de pasto (g MS/m2) bajo la copa en los potreros y se tuvieron 6 repeticiones en cada manejo del potrero y por árbol se cosechó la cobertura herbácea en los cuatro puntos cardinales bajo la copa y fuera de la copa (pastura abierta) a 10 m del tronco. El pasto fue cortado a una altura de 10 cm sobre el suelo utilizando un marco de 1.0 x 1.0 m. Los árboles fueron clasificados en copas livianas, moderadas y densas, según el nivel de sombra medida indirectamente por medio de la disponibilidad de pasto bajo las copas. Los límites de cada categoría se determinaron por medio de la opción tabla de frecuencias del programa estadístico Statgraf (2007). La disponibilidad de pasto se analizó por medio de una prueba de t para conocer si hubo significancia (P<0,05) entre las condiciones de sombra y pastura. Se realizó la prueba de significancia de Tukey al 95 % de probabilidad en lo referente a altura y diámetro de tronco. Los análisis fueron llevados a cabo por medio del programa Stafgraff. Resultados y Discusión. Inventario de especies La mayoría de las especies se determinó por lo menos hasta el género. En total se registraron 411 árboles que pertenecen a 13 géneros y 12 familias. La familia con mayor número de especies fue la Malvaceae (2 especies), el resto de familias tuvieron al menos una especie en cada predio evaluado (Cuadro 1). Cuadro 1. Especies identificadas en dos sistemas de pastoreo. Los Ríos. 2013 Nombre común Familia Género Especie Laurel Boraginacea Cordia alliodora Fernánsanchez Polygonacea Triplaris cumingiana Teca Lamiacea Tectona grandis Tambor Caesalpiniaceae Schizolobium parahybum Guadua Poaceae Guadua angustifolia Naranjo Rutácea Citrus sinensis Mata palo Morácea ficus sp. Cedro Meliácea Cedrela odorata Guarumo Urticácea Cecropia sp. Balsa Malváceae Ochroma pyramidale Chontilla Arecaceae bactris maraja Palo prieto Fabáceae Erythrina poeppigiana Guasmo Malváceae guazuma ulmifolia 156 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Diámetro al pecho Los diámetros de troncos encontrados presentaron crecimientos de entre 25-35%, presentándose diferencias significativas entre los sistemas de pastoreo utilizados. Se observó crecimientos relativamente bajos en el sistema de pastoreo intensivo, siendo el fuste afectado específicamente por el pisoteo generado por el ganado de manera constante y las excretas producidas que afectan el sistema radicular. No se encontró relación entre las zonas de muestreo en el potrero con el tipo de manejo, lo que se determinó un crecimiento parejo de la especie en los lugares colectados. La docima de comparación encontró mejores crecimientos en las zonas perimetrales (20.62 cm) y de cercas (20.60 cm) en el sistema de pastoreo rotativo, debido al poco recorrido hecho por los vacunos en estos sectores. El menor grosor se produjo en los mismo sectores (16.72 cm en zona perimetral y 16.45 en zona de cercas) pero en el sistema de pastoreo intensivo (Cuadro 2, fig. 1). Cuadro 2. Diámetro al pecho en evaluación de Cordia alliodora en dos sistemas de pastoreo. Los Ríos. 2013. Tratamientos Zonas de Influencia Cercas Centro Perímetro Promedio cm (**) Pastoreo Intensivo 16.72 c 17.55 bc 16.45 c 16.90 b Pastoreo Rotativo 20.62 a 20.40 ab 204.00 20.54 a 18.67 18.97 18.53 18.72 Promedio Subtratamientos ns Coeficiente de variación (%) 6.84 Figura 1. Comportamiento del diámetro de Cordia alliodora al inicio y final del ensayo. 2013. Altura del árbol La altura de árboles encontraron diferencias en las evaluaciones realizadas con promedios de 31 a 38 %. Se presentaron diferencias significativas entre los sistemas de pastoreo siendo la mayor altura la registrada con pastoreo rotativo (15.85 m). Entre las zonas evaluada se encontró mayor respuesta en la zona central de los lotes (15.41 m), a diferencia de la zona de cercas (11.55 m) que tuvo menor altura. 157 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Las interacciones de altura y zonas presentaron incrementos en el sistema de pastoreo rotativo en la zona central (19.10 m), siendo diferente estadísticamente a las demás zonas y sistemas. Se tuvo un coeficiente de variación de 10.20 % (Cuadro 3, fig 2). Figura 2. Comportamiento de la altura de Cordia alliodora al inicio y final del ensayo. 2013. Cuadro 3. Altura de árboles en evaluación de Cordia alliodora en dos sistemas de pastoreo. Los Ríos. 2013. Tratamientos Zonas de Influencia Cercas Centro Perímetro Promedio m (**) Pastoreo Intensivo 9.55 d 11.72 cd 11.55 cd 10.94 a Pastoreo Rotativo 13.55 bc 19.10 a 14.90 b 15.85 b Promedio Subtratamientos (**) 11.55 b 15.41 a 13.22 ab 13.39 Coeficiente de variación (%) 10.20 Área de la copa Se presentaron diferencias en el área cubierta por la copa de Cordia observándose incrementos de entre el 12 y el 21 %. Diferencias significativas entre los sistemas de pastoreo fueron reportadas con la prueba de T, siendo el área la encontrada con pastoreo rotativo (10.15 m) la de mayor cobertura. Las zonas de evaluación no presentaron respuesta, sin embargo mayor área se cubrió en la zona cercana a las cercas de los lotes (9.93 m). En las interacciones entre las factores se presentó incrementos en el sistema de pastoreo rotativo en la zona de cercas (11.00 m), este fue diferente estadísticamente a las demás interacciones. El coeficiente de variación fue 5.07 % (Cuadro 4, fig. 3). 158 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Cuadro 4. Área de la copa de árboles en evaluación de Cordia alliodora en dos sistemas de pastoreo. Los Ríos. 2013. Tratamientos Zonas de Influencia Cercas Centro Perímetro Promedio m (**) Pastoreo Intensivo 8.87 bc 8.70 c 8.92 bc 8.83 b Pastoreo Rotativo 11.0 a 9.62 bc 9.82 b 10.15 a 9.93 9.16 9.37 9.49 Promedio Subtratamientos (ns) Coeficiente de variación (%) 5.07 Figura 3. Comportamiento del área de la copa de Cordia alliodora al inicio y final del ensayo. 2013. Disponibilidad de pasto bajo la copa arbórea La disponibilidad de pasto en el área de influencia de la copa arbórea (Cuadro 5, fig. 4), encontró diferencias significativas en las evaluaciones en Cordia alliodora, incrementos del 11-38 % se observaron lo cual se podría asociar a que presentar una copa pequeña o reducida permite mayor entrada de luz solar (Esquivel et al., 2007). Los sistemas de pastoreo según la prueba inferencial de Tukey dieron mayor disponibilidad de pasto con pastoreo rotativo (285.93 g MS/m2) la de mayor cantidad. Tukey no presentó respuesta en la zonas de muestreo, encontrándose mayor disponibilidad en la zona perimetral de los lotes (253.93 g MS/m2). Las interacciones entre sistemas y zonas produjeron incrementos en el sistema de pastoreo rotativo en la zona de cercas (291.07 g MS/m2), zona central (279.02 g MS/m2) y zona perimetral (287.70 g MS/m2), estos fueron diferentes estadísticamente a las demás interacciones. El coeficiente de variación fue 7.13 %. 159 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Cuadro 4. Área de la copa de árboles en evaluación de Cordia alliodora en dos sistemas de pastoreo. Los Ríos. 2013. Tratamientos Zonas de Influencia Cercas Centro Perímetro Promedio g MS/m2 (**) Pastoreo Intensivo 185.17 b 204.12 b 220.17 b 203.15 b Pastoreo Rotativo 291.07 a 279.02 a 287.7 a 285.93 a 238.12 241.57 253.93 244.54 Promedio Subtratamientos (ns) Coeficiente de variación (%) 7.13 Figura 4. Disponibilidad de pasto debajo de copa en Cordia alliodora al inicio y final del ensayo. 2013. El excesivo pastoreo en los lotes del ensayo y el mal diseño del mismo, generaron efectos negativos por la falta de alimento y lento crecimiento de las especies estudiadas, afectando en parte la cobertura del suelo, aumentando el efecto de erosión disminuyendo la fertilidad y de la capacidad de infiltración del agua, generando el ganado compactación del suelo y por la eliminación de excretas la contaminación de las fuentes de agua (Sostenido por Betancourt et al. .2003) Casi todos los manejos pastoriles presentan deficiencias en el crecimiento de especies de acompañamiento, debido a que la mayoría de las especies nativas tienen un crecimiento comparativamente lento; sin embargo, para esto se necesita una acción rápida para instalar sistemas agroforestales y silvopastoriles. A largo plazo, los bosques secundarios en programas silvopastoriles en la región estudiada, son la fuente más importante de trabajo. El manejo racional de los mismos sólo depende de los agricultores y ganaderos, para la reforestación con especies nativas. Falta mucho aun conocer sobre ciclos de reproducción, requisitos ecológicos y condiciones para un adecuado manejo de las plantaciones en los potreros en la mayoría de especies nativas, más estudios aplicados al respecto deben de ser realizados para dar una base sólida a cualquier programa silvopastoril. 160 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Bibliografía Alvarado, A.; Fallas, JL. 2004. Efecto de la saturación de acidez sobre el crecimiento de la teca (Tectona grandis L.f.) en Ultisoles de Costa Rica. Agronomía Costarricense 28(1): 81-87. Betancourt, K.; Ibrahim, M.; Harvey, C.; Vargas, B. 2003. Efecto de la cobertura arbórea, sobre el comportamiento animal en fincas ganaderas de doble propósito en Matiguas, Matagalpa, Nicaragua. Agroforestería en las Américas Vol. 10 Nº39-40:47-51. Corporación de Desarrollo Forestal y Maderero del Ecuador (eds.). 2001. Guías técnicas para el establecimiento y manejo de plantaciones forestales productivas en el litoral ecuatoriano. CORMADERA/OIMT. Proyecto Piloto para la Reforestación y Rehabilitación de Tierras Forestales Degradadas en el Ecuador (PD 17/97 Rev.3F). Quito, Ecuador. 179 p. Esquivel, H.; Ibrahim, M.; Harvey, C.; Villanueva, C.; Benjamín, T.; Sinclair, F. 2003. Árboles dispersos en potreros de fincas ganaderas en un ecosistema seco de Costa Rica. Agrofor. Amer., 10 (39-40): 24-29. Harvey, C.A.; Haber, W. A. 1999. Remnant trees and the conservation of biodiversity in Costa Rican pastures. Agroforestry Systems, 44: 37-68. Loayza, M. del R.; León, V. 1996. Evaluación bajo pastoreo bovino del kikuyo (Pennisetum clandestinum Hochst) en asociación con tres especies forestales. Revista Rumipamba 10(1): 65. Myers, N.; Mittermeier, R.A.; Mittermaier, C.G.; DA Fonseca, G.A.B.; Kent, J. 2000. Biodiversity hotspots for conservation priorities. Nature, (403/25): 853-858. Raigosa, J. 1968. Efecto del abonamiento sobre el crecimiento inicial de plantaciones de Anthocephalus cadamba Miq. y Cordia alliodora (Ruiz y Pav.) Cham. en dos tipos de suelo. Tesis de Maestría. Instituto Interamericano de Ciencias Agrícolas, Turrialba, Costa Rica. 102 p. Restrepo, C.; Ibrahim, M.; Harvey, C.; Harmand, M. Morales, J. 2004. Relaciones entre la cobertura arbórea en potreros y la producción bovina en fincas ganaderas en trópico seco en Cañas, Costa Rica. Agrofor. Amer., 41-42: 29-36. Restrepo, C. 2002. Relaciones entre la cobertura arbórea en potreros y la producción bovina en fincas ganaderas en el trópico seco, Cañas, Costa Rica. M. Sc. Tesis,Turrialba, CR, CATIE. 102 p. Servicio Nacional de Áreas Protegidas-SNAP. 2010. Informe Anual. Ministerio del Ambiente. QuitoEcuador. P. 32. 161 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 EFECTOS DE LA EXCLUSIÓN DE GRANDES HERBÍVOROS SOBRE LA VEGETACIÓN Y EL SUELO, EN MINAS DE CARBÓN RESTAURADAS EN EL NORTE DE PALENCIA Franklin Sigcha Morales, Carolina Martínez-Ruiz Área de Ecología, E.T.S de Ingenierías Agrarias de Palencia, Universidad de Valladolid, Palencia-España [email protected] Introducción Aunque la superficie terrestre alterada por la minería es relativamente baja a nivel mundial, las cifras pueden ser considerables al realizar un análisis por países. En el norte de España la minería de carbón a cielo abierto ha afectado aproximadamente a 5.000 hectáreas de terreno, lo que genera fuertes impactos ambientales localizados (Moreno-de las Heras et al., 2008). Además, la minería va en aumento y sus impactos son, más severos que los provocados por otros tipos de actividades (Cooke y Johnson, 2002). La restauración de las áreas afectadas por la minería es una necesidad urgente, por lo que se deben buscar soluciones que respondan a las necesidades específicas de cada lugar, no sólo en términos del tipo de operación minera, sino también de clima, topografía, sensibilidad del entorno y exigencias sociales que permitan conseguir resultados consistentes con los principios y objetivos del desarrollo sostenible (Gardner, 2001). Por otro lado, el uso ganadero, tras el cese de la actividad minera, es frecuente en el norte de Palencia. Al igual que en otros lugares, este uso se considera adecuado debido a la baja productividad de la tierra requerida en comparación con los cultivos (Maczkowiack et al., 2012). Además, los herbívoros desempeñan un papel fundamental en la regulación de la estructura y función del ecosistema (Cebrián, 2004), lo que es esencial, no sólo para el análisis de las relaciones planta-herbívoro, sino también por sus implicaciones en la rehabilitación o conservación de áreas donde el pastoreo puede ser un factor clave que limita la revegetación. Aunque el efecto del pastoreo en los ecosistemas ha sido repetidamente estudiado, muchos de los resultados son contradictorios (Olff y Ritchie, 1998), ya que los estudios se han realizado en diferentes hábitats, a diferentes escalas espaciales y temporales, con diferentes abundancias, tipos y especies de herbívoros, y al hecho de haber utilizado diferentes protocolos experimentales para el análisis de la información (Bigger y Marvier, 1998). Pero pocos trabajos se han interesado por el efecto de la herbivoría en la vegetación de zonas restauradas tras la minería, lo que hace difícil discernir patrones claros de cuándo y dónde los herbívoros tienen efectos importantes (Gurevitch et al., 2000). Por ello, este trabajo plantea: Determinar los efectos de la herbivoría sobre la vegetación y el suelo, en una mina de carbón restaurada en el noroeste de Palencia. Objetivos El objetivo general de este trabajo es estudiar el efecto de la exclusión de grandes herbívoros en la composición, estructura y producción de las comunidades vegetales, así como en el suelo, en una mina de carbón restaurada en Muñeca (Palencia-España). Para alcanzar este objetivo general se plantearon los siguientes objetivos específicos: a) Caracterizar las comunidades vegetales de las zonas con y sin pastoreo, en términos de diversidad, cobertura y biomasa aérea. 162 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 b) Caracterizar los suelos de las comunidades vegetales con y sin pastoreo, a través del análisis de diferentes parámetros físicos y químicos. c) Analizar la influencia de los cambios en el suelo sobre la composición florística. d) Modelizar el patrón de respuesta de las principales especies de plantas en relación con los cambios en el suelo derivados de la exclusión de herbívoros. MATERIAL Y MÉTODOS Área de estudio El área de estudio se ubica en una antigua mina de carbón a cielo abierto en Muñeca, término Municipal de Guardo. Se trata de una zona montañosa con una altitud media de 1210 m, que pertenece a la cuenca carbonífera “Guardo Cervera”, una de las más importantes de Castilla y León. El clima es Mediterráneo Sub-húmedo, con menos de 1000 mm de precipitación media anual, concentrada en primavera y otoño, y una temperatura media anual de 9,3 ºC, existe riesgo de heladas seguras desde diciembre hasta marzo, y de heladas probables el resto del año excepto durante el periodo xérico (Torroba, 2011). La vegetación que rodea la zona experimental comprende bosques de Quercus pyrenaica, restos de matorrales naturales (Genista florida, Cytisus scoparius) y una compleja matriz de pastizales de anuales dominados por Bromus y Vulpia, entre otras especies características (Alday et al., 2011). Diseño experimental En la mina de Muñeca (restaurada en noviembre de 2006) se seleccionó una ladera con suave pendiente, orientada al norte, usado por ovejas y especies silvestres (ciervos y corzos). En febrero del 2008, se procedió a la exclusión de grandes herbívoros en una superficie de 50x50 m que fue vallada, justo al lado de la zona vallada se delimitó otra, de las mismas dimensiones, sin exclusión de grandes herbívoros. En junio de 2010, dentro de cada zona se establecieron, cinco parcelas de muestreo rectangulares, paralelas a la línea de pendiente. En cada una de las parcelas se muestreó la vegetación en 10 inventarios de 50x50 cm. Muestreo de la vegetación En cada inventario se tomó nota de los porcentajes de: suelo descubierto; suelo cubierto por piedras; cobertura de briofitos y cobertura en proyección vertical de cada una de las especies encontradas. Las especies se identificaron y se denominaron de acuerdo con Flora Europaea. Para la determinación de biomasa aérea se cortó manualmente, a ras del suelo, toda la vegetación presente en un cuadrado de 20x20 cm, seleccionado aleatoriamente dentro de cada cuadrado de 50x50cm. En el laboratorio, la biomasa se dividió en cuatro grupos taxonómicos: Asteraceae, Fabaceae, Poaceae y "otras" para el resto de las familias, ya que estudios previos en zona habían puesto de manifiesto que éstos eran los grupos taxonómicos más frecuentes y abundantes en las comunidades de plantas sobre escombreras de carbón (González-Alday y Martínez-Ruiz, 2007). Muestreo de suelos Simultáneamente al muestreo de vegetación se tomaron las submuestras de suelo, una junto a cada inventario de vegetación. Para ello, se uso una azada, y se cabo hasta aproximadamente unos 10 cm. Las 10 submuestras se mezclaron para ser analizadas como una única muestra. Por tanto, se analizaron un total de 10 muestras de suelo, una por parcela de muestreo. Tratamiento de los datos 163 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Objetivos a y b: las diferencias en los parámetros edáficos y de estructura de la vegetación, se analizaron mediante contraste de hipótesis de igualdad de medias. Se empleó estadística paramétrica y no paramétrica. Objetivo c: se empleó el DCA para decidir si una aproximación unimodal era adecuada al análisis de los datos, tanto de composición florística, como para describir la relación entre ésta y las variables edáficas (Ter Braak y Šmilauer, 2002). Luego se eligió el CA para ordenar las parcelas de muestreo en función de su composición florística. Para valorar la relación entre composición florística y variables edáficas se utilizó el proceso de “forward selection” aplicado al análisis CCA, luego el test de Montecarlo se usó para evaluar la significación de las variables seleccionadas y del modelo en su conjunto (Legendre y Legendre, 1998). Objetivo d. Se utilizaron los modelos de HOF para modelizar el patrón de respuesta de las especies individuales frente al gradiente asociado al pastoreo-no pastoreo, así como frente a los valores concretos de las variables edáficas significativas. RESULTADOS Y DISCUSIÓN Parámetros edáficos en comunidades con y sin pastoreo Los resultados muestran una clara influencia de la exclusión de grandes herbívoros en las propiedades edáficas, como se ha puesto de manifiesto en numerosos estudios. El pastoreo ha supuesto un incremento del porcentaje de arena y un descenso del porcentaje de arcilla. Probablemente, una mayor biomasa aérea incrementa la capacidad de la vegetación para atrapar las partículas finas arrastradas por el viento, aumentando así la proporción de partículas finas en la superficie del suelo y haciendo con ello disminuir la proporción relativa de partículas gruesas (Li et al., 2003). La exclusion ha supuesto un aumento significativo en algunas propiedades químicas del suelo, CE, K +, Ca2+, P, nitrógeno total y materia orgánica total y oxidable. El mayor contenido de materia orgánica en la comunidad sin pastoreo, se puede atribuir a la acumulación de biomasa aérea y hojarasca, que en la comunidad pastada son retiradas periódicamente por los herbívoros, mientras que los menores contenidos en nutrientes (N, K, P) que se registran en la zona pastada podrían derivarse principalmente de los aportes de la descomposición de la biomasa subterránea (Osem et al., 2004). Existen resultados contradictorios que nos deben hacer reflexionar sobre la importancia que puede tener el tiempo de exclusión, la intensidad de pastoreo y el tipo de ambiente en los diferentes estudios llevados a cabo. Parámetros estructurales de la vegetación con y sin pastoreo Al contrario de lo encontrado por otros autores, en este estudio no se encuentra influencia del pastoreo sobre la diversidad de especies o sus componentes. Esta ausencia de cambios significativos puede atribuirse al corto período de tiempo transcurrido desde la exclusión de los herbívoros hasta el muestreo; poco más de dos años (Fernández-Lugo et al., 2009). Además, el proceso de cambio de estos ecosistemas puede ser difícil de detectar cuando están sometidos a bajas presiones ganaderas, que pueden tener débiles o incluso efectos nulos sobre la riqueza de especies (Milchunas y Lauenroth, 1993), o bien estos efectos pueden ser poco evidentes en áreas con producciones inferiores a 200 g/m 2 (Osem et al., 2004). En ambientes pobres, como es el caso de las minas restauradas, la ausencia de pastoreo provoca un aumento de la cubierta vegetal y altura de la vegetación, que se traduce en un incremento de los valores de biomasa aérea (Rueda y Rebollo, 2006). La hojarasca en este sentido también contribuye al reducir la 164 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 erosión del suelo al reducir la escorrentía, y mejora la estructura del suelo, y la fertilidad mediante la adición de materia orgánica (Naeth, 1988). Composición florística y variables edáficas El análisis CA aplicado al conjunto de los datos muestra una ordenación, de las parcelas y especies a lo largo del primer eje, situándose las parcelas con herbivoría (F1-F5) en el extremo positivo del mismo y las parcelas con exclusión de herbívoros (D1-D5) en el negativo (Figura 1). Estos resultados muestran, por tanto, claras diferencias en composición florística entre la comunidad pastoreada y la no pastoreada. Ambas comunidades presentan un elevado número de especies exclusivas. No obstante, a pesar de haberse encontrado diferencias significativas en varios de los parámetros edáficos, entre las comunidades con y sin pastoreo, sólo tres variables edáficas (MOT, K y Mg) están relacionadas significativamente con esas diferencias en composición florística de acuerdo con el análisis de “forward selection” (Figura 2). En concreto, los valores más altos de materia orgánica total, potasio y magnesio están asociados a la comunidad sin pastoreo. 2,5 D3 Con arv Ama sp 2 Cre 2 Rum ace Ger mol Bro max 1,5 Dau car CA2 (λ2= 0,137) Cap bur 1 And int Vul myo Fil pyr Cer fon Cre 1 Fra ves Tri str Lac vir F4 F2 Con spp Cen sp Pet nan Aeg tri Aph mic Cyn ech Hie pil Api Leo his Bro hor Tee cor Ver arv Son ole Tri pra Vul bro Ger rob F3 Cyt esc Lol per Agr cas Fes spp Pap rho Pla lan Poa pra Ero cic Tri glo F1 F5 Tri rep Cer sp D4 Phl pra Hor mur Hyp rad D5 Dac glo Ant arv Tri cam Gal par Dip ful Lot cor Cer glo D1 Vic cra Lep het 0,5 0 -0,5 -1 D2 -1,5 -1,5 -1 -0,5 0 0,5 1 1,5 2 CA 1 (λ=0,293) Figura 1. Primer plano factorial del análisis CA en el que se ordenan las parcelas de muestreo de acuerdo con su composición florística. D1-D5: parcelas sin pastoreo; F1-F5: parcelas con pastoreo. Las especies marcadas en naranja son exclusivas de la comunidad pastoreada, las marcadas en verde exclusivas de la no pastoreada y el resto comunes a ambas, aunque las tres marcadas en rojo muestran abundancias similares en ambas comunidades. Las diferencias en composición florística, a corto plazo, pueden deberse más a diferencias en el aporte de semillas desde los alrededores (Alday et al., 2011) o a una combinación de otros factores abióticos y estocásticos (Alday et al., 2012) que a cambios en las propiedades edáficas. Es bien sabido que en sustratos de origen antrópico, como los estériles de mina, los cambios en el suelo son relativamente lentos (Marrs et al., 1980), y es posible que sea necesario más de dos años de exclusión para encontrar, mayor relación entre los cambios producidos en el suelo y en composición de especies de plantas. 165 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA 1.0 Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 D3 Bro max Con arv Rum ace Ama sp Cre 2 0.8 Ger mol Dau car 0.6 Na Mg Ca MOT 0.4 Cap bur Lac vir Cre 1 Vul myo CE Fil pyr Cer fon F2 0.2 And int Con spp Cen sp Are Fra ves Vul bro Aeg tri Cyn ech Cyt esc F1 Pap rho F5 Agro cas Leo his Tri pra pH 0.0 Ger rob Ero cic D4 D5 Hor mur Phl pra -0.2 F4 Tri str Tee cor Tri rep Cer sp Pla lan Hyp rad Dac glo Tri cam Dip ful Api Pet nan Poa pra Son ole Fes spp Ver arv Bro hor Lol per K Tri glo F3 Hie pil Aph mic Ant arv Lot cor Cer glo Gal par -0.4 Vic cra D1 D2 -0.6 Lep het -1.0 -0.5 0.0 0.5 1.0 Figura 5. Triplot especies-parcelas-variables edáficas del análisis CCA de la zona de estudio. Marcadas con vectores en rojo las tres variables químicas significativas, y en azul, la única variable física incluida en el modelo. D1-D5: parcelas sin pastoreo; F1-F5: parcelas con pastoreo. Respuesta de especies individuales a las diferencias edáficas En este estudio algunas de las especies siguen un patrón de respuesta distinto según el parámetro edáfico respecto al que se han modelizado. A pesar de ello, ha sido posible encontrar alguna tendencia general considerando los rasgos funcionales de las especies (Figura 3). Así Aphanes M, Bromus H y Trifolium R que son igualmente abundantes a lo largo del gradiente definido por el CCA1 son terófitos, mientras que el resto son herbáceas perennes, con forma de vida hemicriptófita e hidrosembradas, a excepción de Dactylis glomerata. Todas las especies con mayor cobertura en la comunidad sin pastoreo son Poáceas (Festuca spp., Dactylis glomerata y Poa pratensis), mientras que entre las especies que aumentan su cobertura hacia la comunidad con pastoreo predominan dos Fabáceas: Lotus corniculatus en la parte media del gradiente y Trifolium repens al final. 25 Fes spp Tri rep % Cobertura 20 15 Dac glo Lot cor 10 Poa pra 5 Lol per 0 -1,02 -1,00 -0,92 -0,42 -0,16 Sin pastoreo 0,95 CCA1 1,28 1,32 1,45 1,54 Con pastoreo Figura 3. Curvas respuesta de las especies derivadas del análisis de los modelos de HOF respecto al gradiente asociado al eje 1 del CCA. En resumen, se modelizaron nueve especies, aquellas que estaban presentes en el 100% de las parcelas de estudio. Aphanes microcarpa fue la única especie cuya cobertura permaneció constante a lo largo de 166 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 los gradientes considerados. La cobertura de T. repens disminuye al aumentar el porcentaje de MOT y K+ en el suelo en la comunidad sin pastoreo. La cobertura de Festuca spp., T. campestre, Dactylis glomerata y Lotus corniculatus aumenta al aumentar el porcentaje de MOT, K+, arcilla y CE en el suelo, y al disminuir el porcentaje de arena en la comunidad sin pastoreo. La cobertura de Poa pratensis aumenta al aumentar la CE en el suelo, y al disminuir el porcentaje de arena en la comunidad sin pastoreo. La cobertura de Lolium perenne disminuye al aumentar el K+ en el suelo en la comunidad sin pastoreo. La cobertura de Bromus horderaceus aumenta al aumentar el % de arena en el suelo en la comunidad con pastoreo. CONCLUSIONES 1.- El pastoreo ha supuesto una modificación de las propiedades físicas (incremento del % de arena y descenso del % de arcilla) y químicas (descenso de CE, K+, Ca2+, P, NT y MOT y MOX) del suelo. 2.- El pastoreo también ha supuesto cambios en la estructura de la vegetación. Un aumento en el % de suelo descubierto y, por tanto, un descenso en la cobertura vegetal y biomasa totales, debido al descenso de la contribución de los tres grupos taxonómicos dominantes (Poaceae, Fabaceae y Asteraceae), así como un descenso en la altura de la vegetación, cobertura de Perennes, cobertura y riqueza de Hemicriptófitos, cobertura de especies Anemócoras y Autócoras, y un aumento de la riqueza de especies endozoócoras. 3.- El aumento de especies con dispersión endozoócora con pastoreo pone de manifiesto la función potencial de las ovejas en la zona como herramienta de restauración, teniendo en cuenta los beneficios de la dispersión de semillas y el control de las especies dominantes introducidas. 4.- Al contrario de lo encontrado en otros trabajos en este estudio no se encuentra influencia del pastoreo sobre la diversidad de especies o sus componentes, posiblemente por la escasez de tiempo transcurrido entre la exclusión de los grandes herbívoros y el muestreo. 5.- Se ha encontrado escasa relación entre las diferencias en composición florística entre las comunidades con y sin pastoreo y los cambios en el suelo, pudiéndose relacionar únicamente con MOT, K y Mg, cuyos valores son más altos en la comunidad sin pastoreo. 6.- Parece existir un reemplazo en el grupo taxonómico que domina desde las Poáceas sin pastoreo hacia las Fabáceas con pastoreo. Sin embargo, no se ha encontrado ninguna tendencia en el mecanismo de dispersión dominante u otras características de las especies individuales. 8.- Teniendo en cuenta el tiempo de exclusión analizado, sería recomendable repetir el muestreo pasado más tiempo para observar si realmente afecta a los resultados obtenidos o hay que buscar explicación en otros factores. BIBLIOGRAFÍA Alday JG.; RH. Marrs and C.Martínez-Ruiz. 2012. Soil and vegetation development during early succession on restored coal wastes: a six-year permanent plot study. Plant Soil 353, 305–320. Alday JG.; Y. Pallavicini; RH. Marrs and C. Martínez-Ruiz. 2011. Functional groups and dispersal strategies as guides for predicting vegetation dynamics on reclaimed mines. Plant Ecol 212, 1759–1775. Bigger D and M. Marvier. 1998. How Different Would a World Without Herbivory Be?: A Search for Generality in Ecology. Integrative Biology 60-67. Cebrián, J. 2004. Role of first-order consumers in ecosystem carbon flow. Ecol Lett 7, 232–240. 167 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Cooke JA and MS. Johnson. 2002. Ecological restoration of land with particular reference to the mining of metals and industrial minerals: A review of theory and practice. Environ Rev 10, 41–71. Fernández-Lugo S.; L. Nascimento; I. Saro; L. Bermejo y JR. Arévalo. 2009. Efectos de la exclusión del pastoreo en la diversidad, riqueza, composición de especies y productividad de dos pastizales canarios. En: Sociedad Española para el Estudio de los Pastos. La multifuncionalidad de los pastos: producción ganadera sostenible y gestión de los ecosistemas. Eds. R. Reiné.; O. Barrantes, A. Broca y C. Ferrer. 155-162. Gardner J. 2001. Rehabilitación de minas para el mejor uso del terreno: la minería de bauxita en el bosque de jarrah de Australia Occidental. Unasylva 207, Vol. 52, 3-8. González-Alday J. y C. Martínez-Ruiz. 2007. Cambios en la comunidad vegetal sobre estériles de carbón tras las hidrosiembra. Ecología 21, 59-70. Gurevitch J.; JA. Morrison and LV. Hedges. 2000. The interaction between competition and predation: a meta-analysis of field experiments. Am Nat 155, 435–453. Legendre P. and L. Legendre. 1998. Numerical ecology. Elservier Science, Amsterdam, ND. Li FR.; H. Zhang; TH. Zhang and Y. Shirato. 2003. Variations of sand transportation rates in sandy grasslands along a desertification gradient in northern China. Catena 53, 255–272. Maczkowiack RI.; CS. Smith; GJ. Slaughter; DR. Mulligan and DC. Cameron. 2012. Grazing as a postmining land use: A conceptual model of the risk factors. Agr Syst 109, 76–89. Marrs RH.; RD. Roberts and AD. Bradshaw. 1980. Ecosystem development on reclaimed china clay wastes. I. Assessment of vegetation and capture of nutrients. J Appl Ecol 17, 709–717. Milchunas DG. and WK. Lauenroth. 1993. Quantitative effects of grazing on vegetation and soil over a global range of environments. Ecol Monogr 63, 327-366. Moreno-de las Heras M.; JM. Nicolau and T. Espigares. 2008. Vegetation succession in reclaimed coalmining slopes in a Mediterranean-dry environment. Ecol Eng 34, 168–178. Naeth MA. 1988. The impact of grazing on litter and hydrology in mixed prairie and fescue grassland ecosystem of Alberta. PhD University of Alberta, Canada. Olff H. and ME. Ritchie. 1998. Effects of herbivores on grassland plant diversity. Trends Ecol Evol 13, 261-265.) Osem Y.; A. Perevolotsky and J. Kigel. 2004. Site productivity and plant size explain the response of annual species to grazing exclusion in a Mediterranean semi-arid rangeland. J Ecol 92, 297-309. Ter Braak CJF.and P. Šmilauer. 2002. CANOCO Reference Manual and User’s Guide to Canoco for Windows: Software for Canocical Community Ordination (Version 4.5). Microcomputer Power: Ithaca, NY. Torroba P. 2011. Papel de los matorrales en la reforestación con Quercíneas de escombreras de carbón, en el municipio de Guardo (provincia de Palencia). Trabajo de fin de carrera ETSIIA de Palencia. Universidad de Valladolid. Ingeniero de Montes-Marzo. Walker LR. and MR. Willig. 1999. An introduction to terrestrial disturbances. In Ecosystems of disturbed ground. Edited by L. Walker. Ecosystems of the world, Elsevier, Amsterdam. pp. 1–16. 168 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 ESTRUCTURA POBLACIONAL, PRODUCTIVIDAD Y FENOLOGÍA FLORAL DE LA TAGUA (Phytelephas aequatorialis) EN LA COSTA ECUATORIANA Janice Jácome Andrade y Rommel Montúfar Lab. 113 Ecología y Genética, Pontificia Universidad Católica del Ecuador. Av. 12 de Octubre 1076 y Roca. [email protected] Introducción La extraccion de recursos forestales no maderables ha sido considerada como una alternativa de conservacion de los ecosistemas en America Latina (Freckleton et al. 2003). Sin embargo, los productos extraidos de bosques tropicales han sufrido fuertes presiones extractivistas, que conllevan un costo ecologico en la salud de las poblaciones (Uma et al. 2004). Existe un controvertido debate sobre las bondades y desventajas del extractivismo. Por un lado se han establecido costos ambientales como la declinacion de la abundancia del recurso, cambios en la demografia y dinamica poblacional (Olmsted y Alvarez 1995), reduccion del flujo genico y cuellos de botella (Uma et al. 2004). A nivel de comunidades humanas se pueden citar varios conflictos relacionados, como la perdida del conocimiento etnobotanico (Pinard y Putz 1992), la inequidad en la cadena de valor de los productos y la falta de incentivos y capacitacion para el manejo de los recursos. Existe ademas el riesgo inminente de que la demanda exceda la capacidad de produccion y, en consecuencia, que se merme progresivamente el recurso, antes de lograr la estabilizacion de los mercados y la regulacion del marco legal en torno al aprovechamiento de los recursos (Uma et al. 2004). Por otro lado el extractivismo aplicado adecuadamente, de manera responsable y no exhaustiva, puede ser una alternativa sustentable a través del tiempo, pues promueve la proteccion de áreas naturales y el cultivo de plantas nativas; de esta manera las especies son mantenidas en un estado seminatural, en lugar de recurrir a la tala de bosques (Borgtoft-Pedersen 1992). Las palmas constituyen uno de los grupos vegetales de mayor utilidad para las poblaciones locales (Borgtoft-Pedersen y Skov 2001), principalmente en regiones tropicales (Byg y Balslev 2006). Sin embargo, su aplicabilidad como modelos extractivistas ha sido pobremente explotada en el país (BorgtoftPedersen y Balslev 1992). En Ecuador el extractivismo de muchos productos forestales no maderables tiene importancia economica a escala local, y representa para las poblaciones rurales la mayor fuente de recursos económicos (Kvist et al. 2006), por ejemplo las hojas de cade, la fibra de mocora o la palma de ramos proveen de importantes ingresos a las comunidades rurales de Ecuador. Por otro lado, la tagua es uno de los pocos ejemplos de extractivismo con importancia comercial en el mercado exterior. Phytelephas aequatorialis Spruce, localmente conocida como tagua o cade, es una palmera dioica solitaria, con un tallo de hasta 10 m de altura y ~30 cm de diametro (Barfod 1991; Dransfield y Uhl 2008), infrutescencia esferica de ~30 cm de diametro con ~10–30 frutos conicos, cada uno de los cuales alberga 5–7 semillas. Es endemica del litoral y las regiones tropicales y subtropicales de las estribaciones occidentales de Ecuador, encontrandose tanto en bosques deciduos y semideciduos, como premontanos y montanos, hasta 1500 msnm (Borchsenius et al. 1998). La palma de tagua ha sido tradicionalmente usada de múltiples formas por las poblaciones humanas. Varias partes de la palma (mesocparpo, endospermo, palmito, cotiledón, inflorescencias) son consumidas por personas y animales (Koziol y Borgtoft-Pedersen 1993). El endospermo maduro de la semilla 169 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 constituye la tagua o marfil vegetal, a partir de la cual se fabrican artesanías, bisutería y precursores de botones (animelas), estos últimos principalmente para exportación. Las hojas (cade), son utilizadas para la elaboración de techos en áreas rurales del litoral. Tras 150 anos de extractivismo de la palma de tagua y su uso en agroforesteria, la cosecha e industrializacion de sus productos derivados ha pasado de ser una actividad de subsistencia, a una comercial e incluso destructiva. La palma de tagua ha sido empleada como componente arboreo en sistemas silvopastoriles y agroforestales, mismos que manejados adecuadamente podrian ser una alternativa a la recuperacion de los ecosistemas (Borgtoft-Pedersen y Balslev 1992). Sin embargo, a pesar de la importancia economica de esta especie como recurso forestal no maderable, poco se conoce sobre su ecologia, y no existen estudios que permitan tener una aproximación del estado de conservacion de las poblaciones, ni del valor económico que el extractivismo del recurso genera, sobre todo en la provincia de Manabi; esta información es clave para desarrollar programas de manejo y agroforestería de la especie. Los objetivos del presente trabajo fueron: (i) Estudiar la estructura poblacional y fenología de la palma en dos tipos de hábitat (pastizal y bosque secundario), y (ii) Describir la productividad de estructuras vegetativas y reproductivas y los potenciales beneficios económicos generados por sus recursos derivados (cade y tagua). Metodología Se realizaron 6 salidas al campo, durante la primera semana de los meses de julio, septiembre y noviembre de 2011 y febrero, mayo y julio de 2012. Se estudiaron dos poblaciones de P. aequatorialis en Manabí. La primera poblacion se ubica sobre una planicie con pastizales en la parroquia Canuto del canton Chone (00º49'18" S 80º08'43" W; 37 msnm), con área total de 5184 m2. En este sitio las hojas de cade se cosechan anualmente y las semillas de tagua se recogen al caer al suelo. El terreno, y las palmas en general, se exponen a una alta incidencia de luz solar (categoría 4.5 en una escala de 1–5; siendo 5 la exposicion total). La segunda población se ubica en la localidad Balsa Tumbada del canton Junin (00º56'20" S 80º14'24" W; 93–123 msnm); el terreno constituye un bosque secundario en recuperación sobre una colina con pendiente de ~39º y área total de 4608 m2. La incidencia de luz solar es de 2.5. En el terreno de Balsa Tumbada la tagua y el cade no se cosechan con regularidad. Demografía de las poblaciones: se censaron y georeferenciarion todos los individuos según la siguiente clasificación de estadios regenerativos: P-plántula (altura de 0–0.5 m), J1-juvenil 1 (0.5–1 m), J2 (1–1.5 m), J3 (1.5–2 m), J4 (2–2.5 m), J5 (2.5–3 m), J6 (>3 m), A1-adulto 1 (tronco de 0–2 m), A2 (2–4 m), A3 (4–6 m) y A4 (6–8 m). Producción de hojas: durante la primera salida al campo se marcaron todos los cogollos de los adultos (N=68 individuos en Canuto; N=171 en Balsa Tumbada), en cada uno de los censos posteriores se registró el número de hojas expandidas (hojas nuevas) a partir de cada cogollo (sección apical de la palma). Se obtuvieron valores totales de pérdida foliar en el año (hojas muertas en toda la población), así como el número hojas que cada planta perdió. Adicionalmente se realizó una estimación del crecimiento anual de las palmas, para lo cual se determinó la cantidad de cicatrices foliares en una porción de 30 cm del tronco; se obtuvo un número promedio de cicatrices para todos los individuos (machos y hembras), así como uno específico para las hembras y uno para los machos de cada localidad. El crecimiento anual del individuo fue calculado en función de los números de cicatrices foliares y la tasas de producción anual de hojas. Adicionalmente se consultó a varios actores locales involucrados en el comercio del cade para conocer el precio actual de la unidad de comercialización (ciento de hojas) en cada localidad; de tal manera que 170 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 posteriormente se pudo valorar económicamente la totalidad de hojas producidas durante el tiempo de estudio, es decir, la producción anual. Producción de semillas de tagua: a partir de la segunda salida al campo se registró el número de infrutescencias nuevas, así como las que liberaron tagua en cada hembra (N=38 hembras en Canuto; N=70 en Balsa Tumbada). Se obtuvo un valor aproximado del total de semillas producidas en cada localidad durante el año. Se determinaron valores promedio de semillas por fruto y de frutos por infrutescencia, así como de tamaño y peso de las semillas que salen a la venta, y cuántas de éstas componen un saco de semillas de tagua. Se propuso una valoración económica de los terrenos en términos de productividad de tagua. Fenología floral: se construyó un calendario fenológico del desarrollo total de la infrutescencia. La muestra fue de 200 estructuras reproductivas elegidas independientemente de la hembra a la que pertenecían; se escogieron 20 estructuras de cada fenofase: • • • • • • • • • • Brote en profilo (BP): brote floral inmaduro recubierto por la bráctea y por el profilo (hoja protectora externa). Brote en bráctea (BB): brote floral maduro recubierto por la bráctea (hoja protectora interna), misma que está expuesta debido a la apertura del profilo. Inflorescencia fresca (IF): la bráctea se ha abierto y la inflorescencia emerge, ocurre la antesis. Inflorescencia marchita (IM): se inicia un proceso de oxidación de los tejidos florales. Inflorescencia seca (IS): los óvulos ya fecundados se comienzan a desarrollar dentro de los ovarios. Infrutescencia con semillas en agua (A): el endospermo es líquido y bebible. Infrutescencia con semillas en gelatina (G): el endospermo comienza a solidificar y es comestible. Infrutescencia con semillas en mocoche (M): el endospermo es más compacto y blanquecino, aún es comestible. Infrutescencia con semillas en tagua blanda (TB): el crecimiento de la infrutescencia se detiene. La semilla ya no es comestible. Infrutescencia con semillas en tagua dura hasta la caída de las mismas (TD): el endospermo ha endurado en su totalidad. Los frutos caen y liberan la tagua. Resultados y discusión La estructura poblacional en Canuto (pastizal) se conforma por 51 plántulas, 30 juveniles (J1) y 68 adultos (38 hembras y 30 machos; A2–A3). Antes del inicio del experimento se reportó la tala ~10 machos en el área de estudio; lo que provocó la desviacion del ratio sexual esperado (1:1). La tala selectiva de los individuos en funcion del sexo y del recurso a aprovecharse es una técnica comun en muchas especies dioicas de los trópicos (Bernal et al. 2011). La tercera parte de la poblacion de Canuto corresponde a plantulas; sin embargo, es probable que estos individuos no alcancen el estadio de juvenil 1. La alta tasa de mortalidad durante los estadíos de plántula y juvenil 1 es generada por los efectos negativos de (i) la herbivoría, (ii) la alta tasa de exposición solar que destruye los tejidos foliares, y/o (iii) el efecto del desbroce del terreno por parte de los campesinos. Las poblaciones de tagua en pastizales ofrecen un panorama desalentador debido a la ausencia de regeneración natural; éstas poblaciones dentro de algunas décadas podrían desaparecer de los sistemas silvopastoriles, generando una invaluable pérdida de su diversidad genética (Borgtoft-Pedersen y Balslev 1992). 171 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 En Balsa Tumbada (bosque secundario en recuperación) la población se compone de 546 plántulas, 1987 juveniles (J1–J6) y 171 adultos (70 hembras, 98 machos y 3 individuos sin sexo definido; A1–A4). Informadores locales no reportan eventos de tala selectiva en el área, por lo menos hace 50 anos. Aun asi, la proporcion de sexos se desvia de la esperada (1:1); la densidad de machos fue mayor en la parte mas alta de la colina, donde hay más incidencia de luz solar respecto al resto del terreno. Se ha reportado que las poblaciones de plantas dioicas suelen exhibir desviaciones del ratio sexual como resultado de la segregacion espacial de sexos en relacion a la disponibilidad de recursos limitados (Lloyd y Webb 1977; Allen y Antos 1992; Vasiliauskas y Aarseen 1992). La mayor densidad de machos en la parte alta de la colina sugiere un patrón de segregación sexual, en donde los machos son más aptos para establecerse en microhábitats con mayor incidencia de luz solar (Bierzychudek y Eckhart 1988; Dawson y Blis 1989; Krischik y Denno 1990). Hay una alta proporcion de juveniles 1 en comparacion con otros estadios, y a partir de esta categoria la cantidad de individuos decrece conforme avanza el estadio. Esta conformacion de J invertida denota una mayor estabilidad poblacional, y ha sido observada también por Velasquez-Runk (1998), tanto en ambientes naturales como disturbados. Es así que a diferencia de Canuto, la población de Balsa Tumbada es viable a futuro, pues presenta, la mayoría de los estadios regenerativos. En Canuto se produjeron 592 hojas durante el año (N=68 individuos); en Balsa Tumbada, 1097 hojas (N=171). Cada individuo de Canuto crece a razon de 9.8 cm/ano y produce en promedio 8.7 1.9 hojas/año; cada individuo de Balsa Tumbada crece a razon de 8.6 cm/ano y produce en promedio 6.5 1.7 hojas/año. Al analizar por separado cada sexo, se estimó que los machos de Canuto crecen a razón de 9.9 cm/ano y producen en promedio 9.1 2.2 hojas/año; las hembras crecen a razón de 9.7 cm/ano y producen 8.4 1.5 hojas/año en promedio. En Balsa Tumbada los machos crecen a razón de 8.8 cm/an o y producen en promedio 6.9 1.9 hojas/año, y las hembras crecen a razón de 7.9 cm/ano y producen 5.8 1.2 hojas/año en promedio. La diferencia entre las medias (promedios) de machos y hembras es mayor en Balsa Tumbada; no obstante, los resultados de ambos sitios sugieren que los machos son mas productivos en cuanto a cade. Borgtoft-Pedersen (1993) obtuvo similares resultados en un pastizal ubicado en la Comuna Rio Santiago, provincia de Esmeraldas, en donde los machos produjeron 6.9 0.9 hojas al ano, en tanto que las hembras produjeron 5.4 1.0. En el pastizal (Canuto) la cobertura vegetal del terreno es baja (~1m), por lo que la mayoria de palmas de tagua no tienen impedimento en la captura de luz solar; en cambio, en el bosque secundario (Balsa Tumbada) la cobertura vegetal es de mayor altura (~6m), razón por la cual se obstaculiza el paso de la luz al interior del bosque y se genera competición inter e intra-específica. Borgtoft-Pedersen (1993) encontro una correlacion positiva significativa entre la incidencia de luz solar y el crecimiento de palmas de tagua en sistemas agroforestales de tierras bajas. El nivel de luz solar tambien marca la diferencia en la calidad de las hojas de cade (Borgtoft-Pedersen 1993; Velasquez-Runk 1998). En Canuto la alta incidencia de luz solar genera una mayor produccion foliar, pero las hojas son de menor tamano y robustez. Lo opuesto sucede con el cade de Balsa Tumbada, en donde hay menor incidencia de luz y por ende se generan hojas de mayor tamaño y con tejidos foliares más robustos; estas hojas son más apreciadas para la elaboración de techos. Se estimó la producción anual de tagua de Canuto en 18144 semillas idoneas para la venta, provenientes de 168 infrutescencias abiertas; cada hembra produjo en promedio 477.5 semillas/año. La produccion anual de Balsa Tumbada se estimó en 21276 semillas idoneas, provenientes de 197 infrutescencias abiertas; cada hembra produjo en promedio 303.9 semillas/año. Segun Borgtoft-Pedersen (1993) existe una correlacion positiva significativa entre la intensidad de luz solar y la productividad de infrutescencias en tierras bajas. A pesar de que en el presente estudio no se analizó este parametro en cada una de las 172 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 hembras, los resultados muestran que en Balsa Tumbada, donde las plantas se encuentran menos expuestas al sol en relacion a Canuto, se produce en promedio un menor numero de semillas por infrutescencia; esta correlacion se ha observado tambien en otras especies de palmas, como es el caso de Rhopalostylis sapida (Enright y Watson 1992). Sin embargo, es probable que con las semillas de tagua se de un patrón similar al del cade, es decir, que las semillas provenientes de plantas que crecen bajo la sombra sean de mejor calidad debido a la solidez del endospermo. En el presente estudio no se evaluo el desarrollo continuo de las infrutescencias, más se midió independientemente el tiempo promedio que toma cada fenofase, desde el aparecimiento del brote floral hasta la maduración del endospermo y caída de las semillas; estos tiempos se sumaron para estimar la duración del desarrollo total de los frutos. De acuerdo a datos de Borgtoft-Pedersen (1993) las infrutescencias demoran 3 anos en desarrollarse (Velasquez-Runk 1998); los resultados obtenidos en el presente estudio corroboran este enunciado. El tiempo promedio de desarrollo fue de 2.9 anos en Canuto y 3.0 en Balsa Tumbada, es decir, 977.2 66.4 días y 998.2 57.2 días respectivamente. Aun asi, se encontro que el desarrollo total de las infrutescencias en ambas localidades varia entre 32 y 40 meses, es decir, entre 2 anos 8 meses y 3 anos 4 meses; las fenofases con más duración fueron: TB (infrutescencia con semillas en tagua blanda) con un tiempo promedio de 32 semanas, y TD (infrutescencia con semillas en tagua dura hasta la caída de las mismas), con un tiempo de 36 semanas. En cuanto a las ganancias obtenidas por la venta de recursos provenientes de P. aequatorialis, en el pastizal (Canuto) se estimaron ingresos monetarios de 210.00 USD por la venta de 592 hojas de cade (~6 cientos); cada ciento (100 hojas) al año 2012 tuvo un valor de 35.00 USD; en el bosque secundario (Balsa Tumbada) las ganancias fueron de 440.00 USD por la venta de 1097 hojas (~11 cientos), pues el ciento tuvo un costo de 40.00 USD. En cuanto a la venta de semillas, debido a que el saco de tagua (unidad de comercialización) se compone de ~680 semillas y se vendió en promedio a 9.00 USD en el año 2012 en ambas localidades, la produccion anual de Canuto (18144 semillas; ~27 sacos) generó ingresos monetarios de ~240.00 USD; la produccion anual de Balsa Tumbada (21276 semillas; ~31 sacos) generó ganancias de ~280.00 USD. En términos generales, los réditos económicos estimados por palma fueron mayores en el pastizal. En cuanto a la venta de cade cada palma generó 3.09 USD (0.52 ha; 210.00 USD/68 individuos) en Canuto, y 2.57 USD (0.46 ha; 440.00 USD/171 individuos) en Balsa Tumbada. Un patrón similar fue reportado para la producción de semillas; cada palma generó 6.32 USD (0.52 ha; 240.00 USD/38 hembras) en Canuto, y 4.00 USD (0.46 ha; 280.00 USD/70 hembras) en Balsa Tumbada. A pesar de que las ganancias totales fueron superiores en Balsa Tumbada, los ingresos que cada palma generó en promedio fueron mayores en Canuto, ya que el pastizal es un área con menos individuos pero más productiva que el bosque secundario, tanto para cade como para tagua. El presente estudio constituye una primera aproximación a la valoración económica y manejo de la especie. La zona en la que se realizó el estudio es, tradicionalmente, productora de recursos derivados de la especie, y tiene un gran potencial para el aprovechamiento de estos recursos, debido a la creciente demanda de los productos derivados de las semillas de tagua a nivel nacional e internacional y, en menor medida, a la demanda que el cade genera a nivel local y provincial. Agradecimientos La presente investigación fue financiada por la Pontificia Universidad Católica del Ecuador y por el proyecto europeo PALMS FP7 – Palms Harvest Impacts in Tropical Forest. 173 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Bibliografía Allen, G; Antos, G. 1992. Sex ratio variation in the dioecious shrub Oemleria cerasiformis. American Naturalist. 141:537-553. Barfod, A. 1991. A monographic study of the subfamily Phytelephantoideae (Arecaceae). Opera Botanica. 105:1-73. Bernal, R; Torres, C; Garcia, N; Isaza, C; Navarro, J; Vallejo, M; Galeano, G; Balslev, H. 2011. Palm management in South America. Botanical Review. 77:607-646. Bierzychudek, P; Eckhart, V. 1988. Spatial segregation of the sexes of dioecious plants. American Naturalist. 132:34-43. Borchsenius, F; Borgtoft-Pedersen, H; Balslev, H. 1998. Manual to the palms of Ecuador. AAU Reports 37. Aarhus University, Pontificia Universidad Catolica del Ecuador. Aarhus, DK. 217 p. Borgtoft-Pedersen, H. 1992. Uses and management of Aphandra natalia (Palmae) in Ecuador. Bulletin de l’Institut Francais d’ Etudes Andines. 21(2):741-753. Borgtoft-Pedersen, H. 1993. Ivory nuts, fruits, and thatch: Use and management of Phytelephas aequatorialis (Palmae) in Ecuador. Part 3d in Extractivism in Ecuador with special emphasis on management and economic exploitation of native palms. Tesis PhD. Aarhus, DK, University of Aarhus. Borgtoft-Pedersen, H; Balslev, H. 1992. The economic botany of Ecuadorian palms. Sustainable harvesting and marketing of rain forest products. 173-191. Borgtoft-Pedersen, H; Skov, F. 2001. Mapping palm extractivism in Ecuador using pair-wise comparisons and bioclimatic modeling. Economic Botany. 55(1):63-71. Byg, A; Balslev, H. 2006. Palms in indigenous and settler communities in southeastern Ecuador: Farmers’ perceptions and cultivation practices. Agroforestry Systems. 67(2):147-158. Dawson, T; Bliss, L. 1989. Patterns of water use and the tissue water relations in the dioecious shrub, Salix arctica: The physiological basis for habitat partitioning between the sexes. Oecologia. 79:332-343. Dransfield, J; Uhl, N. 2008. Genera Palmarum: The evolution and classification of palms. Second edition. Royal Botanic Gardens, Kew, UK. 732 p. Enright, N; Watson, A. 1992. Population dynamics of the nikau palm, Rhopalostylis sapida (Wendl. et Drude), in a temperate forest remnant near Auckland, New Zealand. New Zealand Journal of Botany. 30:29-43. Freckleton, R; Silva, D; Bovi, M; Watkinson, A. 2003. Predicting the impacts of harvesting using structured population models: The importance of density-dependence and timing of harvest for a tropical palm tree. Journal of Applied Ecology. 40:846-858. Koziol, M; Borgtoft-Pedersen, H. 1993. Phytelephas aequatorialis (Arecaceae) in human and animal nutrition. Economic Botany. 47:401-407. Krischik, V; Denno, R. 1990. Patterns of growth, reproduction, defense, and herbivory in the dioecious shrub Baccharis halimifolia (Compositae). Oecologia. 83:182-190. 174 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Kvist, L; Aguirre, Z; Sanchez, O. 2006. Bosques montanos bajos occidentales en Ecuador y sus plantas utiles. Botanica Economica de los Andes Centrales. 205-223. Lloyd, D; Webb, J. 1977. Secondary sex characters in plants. Botanical Review. 43:177-216. Olmsted, I; Alvarez, E. 1995. Sustainable harvesting of tropical trees: Demography and matrix models of two palm species in Mexico. Ecological Applications. 5(2):484-500. Pinard, M; Putz, F. 1992. Population matrix models and palm resource management. Bulletin de l’Institut Francais d’ Etudes Andines. 21:637-649. Uma, R; Ganeshaiah, K; Nageswara, M; Aravind, N. 2004. Ecological consequences of forest use: From genes to ecosystem - A case study in the Biligiri Rangaswamy Temple Wildlife Sanctuary, south India. Conservation & Society. 2:347-363. Vasiliauskas, S; Aarseen, L. 1992. Sex ratio and neighbor effects in monospecific stands of Juniperus virginiana. Ecology. 73:622-632. Velasquez-Runk, J. 1998. Productivity and sustainability of a vegetable ivory palm (Phytelephas aequatorialis, Arecaceae) under three management regimes in northwestern Ecuador. Economic Botany. 52:168-182. 175 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 PROPUESTA METODOLÓGICA DE CRITERIOS E INDICADORES DEL MANEJO FORESTAL SOSTENIBLE EN LAS CONDICIONES DE CUBA Jorge Luis Cué García1, Mariol Morejón García2, Juan A. Herrero Echevarría3, Iván Pino Estopiñales3 1Universidad Técnica del Norte. Ecuador de Pinar del Río, Cuba 3Ministerio de la Agricultura, Cuba. 2Universidad [email protected] Resumen. El trabajo se desarrolla en el contexto de la producción forestal cubana teniendo en cuenta la metodología vigente en Cuba de criterios e indicadores del manejo forestal sostenible, Herrero, (2005) y la Guía de Campo para la Validación de Criterios e Indicadores para la Ordenación Forestal Sostenible a nivel de Unidad de Manejo, en Honduras, 2005. Se emplea la revisión documental, Método Delphi para criterios expertos y talleres participativos con actores y decisores de las Unidades Empresariales de Base Silvícolas, Empresa Forestal Integral, del Servicio Estatal Forestal y de organismos afines. La propuesta de criterios e indicadores de manejo forestal sostenible en las unidades empresariales se complementa con la adecuación del Barómetro de Sostenibilidad de Prescott-Allen, (1997). La adopción de un sistema jerárquico facilita la estandarización de los indicadores y su agregación a índices. Se propone la adecuación de la guía de procedimiento de los indicadores y la lista de chequeo de los mismos. Los resultados del analisis de la fiabilidad del instrumento aplicado a los expertos son aceptables pues el valor T2 de Hotelling fue de 322.46 para p = 0.007, lo que indica que los elementos de la escala no poseen la misma media, existiendo una concordancia significativa fuerte entre los elementos del instrumento, expresado en la correlación intraclase con un coeficiente de 0.756 para p = 0.038, coeficiente de confiabilidad Alpha de Cronbach de 0,806, Coeficiente de Spearman-Brown igual a 0,656 y Coeficiente Guttman para mitades partidas de 0,606. Esta alternativa metodológica evalúa la tendencia del manejo forestal sostenible desde las dimensiones de la sostenibilidad y se adapta a las particularidades de cada sistema y unidad de producción forestal, lo que posibilita una observancia más integral y holística de su desarrollo satisfaciendo los atributos valorativos de un sistema de indicadores de sostenibilidad en el ámbito del sector empresarial forestal especializado cubano. Palabras claves: Criterios e indicadores, manejo forestal sostenible. Introducción. La Cumbre de Río, 1992, consagró el concepto de Desarrollo Sostenible11 establecido por la comisión Brundtland, Nuestro futuro común de la Comisión Mundial sobre el Medio Ambiente y el Desarrollo de 1987, promoviéndolo al plano oficial a nivel internacional como marco de referencia obligada en el quehacer de las políticas de gobiernos, asociaciones, empresas. Esta nueva visión del desarrollo, denominado sostenible, redefine la relación entre el sistema económico y el natural y “se entiende a aquel como subsistema del sistema natural global” (Olabe 2003). 11 “El desarrollo sostenible es aquel que satisface las necesidades del presente sin comprometer la capacidad de las futuras generaciones para satisfacer las suyas” 176 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Es relevante el análisis de Yurjeric, (1996), en cuanto a los niveles de visión respecto a lo que expresa el concepto de desarrollo sostenible, para el planeta preocupa la estabilidad de la biósfera; para el país adquiere las características propias de su modelo de desarrollo y que pueda ser transformado en políticas por las autoridades públicas y para el ciudadano tiene sentido si conduce a una calidad de vida que responda a las necesidades y deseos como ser humano. Dirigirse hacia la sostenibilidad implica asumir el cambio como un hecho y un reto en lo conceptual, siendo clave la planificación para la gestión de los recursos naturales, financieros y del potencial humano, así como su expresión en las políticas a nivel internacional y de estados en función del desarrollo. La complejidad de la concepción de la sostenibilidad está dada en el hecho de que trasciende al futuro. Un estado final de sostenibilidad no existe, por eso los indicadores de sostenibilidad pueden permitir de forma realista solo una estimación sobre la dirección correcta de una determinada evolución de un sistema. (Oficina Federal de Agricultura, 2002) Los indicadores de sostenibilidad son la relación entre las diferentes dimensión de la sostenibilidad, reflejando la significación de las dinámicas desarrolladas en la complejidad del sistema más allá de lo reportado por cada dimensión del sistema separadamente IISD,2000, citado por Farsari y Prastacos (2002); brindan señales que facilitan la evaluación del progreso hacia los objetivos que contribuyen a la meta de alcanzar el bienestar humano y del ecosistema de manera armónica. La definición y desarrollo de indicadores de sostenibilidad no es solamente una materia de la ciencia, esto debe ser del consenso de todos, en procesos de carácter participativo, involucrando a la ciencia, los hacedores de políticas y la sociedad civil según Pinter, (2002). La promoción del desarrollo ha sido siempre una tarea a realizar en un territorio concreto, siendo el resultado de la acción de diferentes actores, con distintos intereses que se manifiestan y concretan en procesos económicos, sociales y ambientales. El monitoreo y evaluación del Manejo Forestal Sostenible es un reto de los gobiernos, empresas y sociedad en general en tanto incide en la calidad de vida de las personas, en la calidad de los ecosistemas terrestres y en el buen funcionamiento de la biósfera en el planeta. Cabe preguntase entonces ¿Cuáles son las alternativas más adecuadas para realizar el monitoreo del manejo forestal sostenible? Es la aspiración del presente documento presentar una alternativa metodológica para el seguimiento de los Criterios e Indicadores del Manejo Forestal Sostenible en las condiciones del sector estatal especializado en Cuba. Desarrollo. La presente experiencia se desarrolló en el periodo comprendido de 2006 al 2009 en la provincia de Cienfuegos, ubicada en la región centro sur del archipiélago cubano, limita al norte con la provincia de Villa Clara, al sur con el Mar Caribe, al este con la provincia de Santi Spíritus y al oeste con la provincia de Matanzas. Para la aplicación del Método Delphi se manejó el procedimiento propuesto por el Instituto de Ingeniería de España (2001), que modifica el número de circulaciones de los cuestionarios de cuatro veces a solo dos. Se puso a consideración de los expertos la propuesta de criterios e indicadores siendo validado estadísticamente mediante la evaluación de la fiabilidad del mismo, a través de la determinación de los coeficientes: Correlación entre mitades, Guttman, Spearman – Brown (igual longitud), Spearman – Brown 177 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 (desigual longitud) y el Coeficiente de Confiabilidad Alpha – Cronbach. Además fueron analizados otros estadísticos, entre ellos: Coeficiente de Correlación Intraclase (genera medidas sobre la consistencia o sobre el acuerdo de los valores entre los propios casos), y la Prueba T2 de Hotelling (genera un contraste multivariado sobre la hipótesis nula de que todos los elementos de la escala tienen la misma media), propuestos por Socorro (2002), y los puntos de cortes y el Coeficiente de Concordancia de Kendall. El paquete estadístico empleado fue SPSS versión 13.0 para Windows. El marco metodológico para diseñar el sistema de principios, criterios e indicadores de Manejo Forestal Sostenible se estructuró considerando los siguientes elementos: 1. Concepto de sostenibilidad aplicado. 2. Objetivos del análisis de los principios, criterios e indicadores. 3. Usuarios de la información. 4. Procedimiento de definición y selección de los principios, criterios e indicadores y sus umbrales. 5. Procedimiento de agregación para la evaluación de la tendencia del Manejo Forestal Sostenible. Se asume el concepto de sostenibilidad de Gómez, (2007), etimológicamente el término sostenibilidad se refiere a la garantía de continuidad en el tiempo de las características estructurales y funcionales, de los valores y de los atributos, de aquello a lo que se refiere: sistema, recurso, actividad o proceso. Principios básicos para la construcción del sistema de indicadores elaborados por los autores. 1. Correspondencia con el concepto de Manejo Forestal Sostenible expresado en los principios que lo regirán. 2. Correspondencia con los Criterios e Indicadores establecidos por el Servicio Estatal Forestal de Cuba. 3. Correspondencia de su diseño con la información de las variables e indicadores que se monitorean en el sector forestal y organismos afines. 4. Los indicadores que se conciben no requieren de nuevas y significativas inversiones para su monitoreo, sino solamente, aprovechar la infraestructura y los recursos humanos existentes de acuerdo al funcionamiento de las unidades silvícolas dentro del sector forestal y organismos afines. La Guía de Campo para la Validación de Criterios e Indicadores para la Ordenación Forestal Sostenible a nivel de Unidad de Manejo, en Honduras, 2005, así como, los Criterios e indicadores de Manejo Forestal Sostenible para Cuba, 2005, sirven de referente principales para la propuesta de sistemas de Criterios e Indicadores que se muestra y la Guía de Procedimiento. La adecuación de los indicadores del Manejo Forestal Sostenible al modelo de Prescott – Allen 1997, agrupados en Bienestar Humano y Bienestar del Ecosistema, se adopta por consenso durante el desarrollo de las sesiones de trabajo en cada Unidad Empresarial de Base Silvícola, atendiendo a su mayor relevancia en cuanto a su impacto en los bienestares. Se propone el proceso de agregación de los indicadores a los Índices de Bienestar Humano y del Ecosistema y su representación en el Barómetro de Sostenibilidad. Resultados y discusión. Se acepta como adecuada la propuesta de jerarquización de Bueren y Blom (1997), metodología basada en un sistema de pensamiento jerárquico llamada genéricamente Criterios e Indicadores, que permite la formulación de un sistema de evaluación o la propuesta de metas a alcanzar en el manejo forestal sostenible y en especial, una base de discusión sobre aspectos tanto filosóficos como científicos, técnicos 178 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 y operativos del mismo. Incrementa la posibilidad de cubrir los aspectos más relevantes a evaluar, evita la redundancia, limita el conjunto de principios, criterios e indicadores (P, C e I), permite también tener una referencia constante del fin hacia el cual está dirigido el manejo forestal sostenible y posibilita chequear la pertinencia de las políticas ejecutadas, (Bueren y Blom, 1997, Rusch y Sarasola, 1999). Principio Criterio Indicador Verificador Figura 1. Esquema jerárquico para la formulación de estándares para el manejo forestal sostenible Bueren y Blom, (1997). Experiencia internacional Contexto cultural y político Talleres Validación Visión Técnicos Primera Expectativa y visión actores propuesta Actores claves Principios básicos. Dinámica Trabajadores Directivos y especialistas Académicos y Científicos Revisión Continua Criterios e Monitoreo Indicadores Retroalimentación Guía de procedimientos Figura 2. Metodología de Rayen Quiroga, (2003). (Adaptada por el autor) A cada indicador le fue confeccionada una Guía de Procedimiento para los indicadores, elaborada por los autores, que los caracteriza y detalla el método de trabajo del mismo, la que contiene los siguientes aspectos: nombre del indicador, definición, ubicación según bienestar, tipo de escala, comportamiento ascendente favorable: Positivo o Negativo, unidad de medida del indicador, fórmula del indicador, cobertura de la información, fuente de los datos, periodicidad de los datos, responsable de la medición. 179 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 La escala del valor del puntaje del indicador se adecuó por los autores, quedando de la siguiente manera los intervalos: 0-30; 30.01-60 y 60.01-90, que se corresponde con la adecuación del Barómetro de Sostenibilidad de Prescoot-Allen desarrollado por Cué et al., (2008) propuesta que permite visualizar la valoración integral de la tendencia a la sostenibilidad de un sistema dado desde la perspectiva de la interpolación de los resultados para los Índices de Bienestar, destacando, por tanto, cual es el subsistema de mayor influencia sobre la sostenibilidad. Los índices agregados, en su totalidad, se calculan mediante la siguiente formulación Dónde: I(x): Índices agregados n I ( x) PI / n i 1 PI: Valor del puntaje de los indicadores que comprende. N: Número de indicadores evaluados. Los resultados del análisis de la fiabilidad del instrumento aplicado a los expertos fueron favorables, los que pueden apreciarse en los estadígrafos analizados según tabla 1. El valor T2 de Hotelling fue de 322.46 para p = 0.007, lo que indica que los elementos de la escala no poseen la misma media, existiendo una concordancia significativamente fuerte entre los elementos del instrumento, expresado en la correlación intraclase con un coeficiente de 0.756 para p=0.038. Tabla 1. Coeficientes de confiabilidad del instrumento para la consulta a expertos acerca de los sistemas de Criterios e Indicadores. Coeficiente de confiabilidad Alpha de Cronbach Parte 1 0,806 Parte 2 0,632 Correlación entre mitades 0,488 Coeficiente de Spearman-Brown Coeficiente Guttman para mitades partidas Igual longitud 0,656 Desigual longitud 0,656 0,606 Fuente: Elaborada por los autores. Los resultados de la consulta a expertos respecto a la propuesta del sistema de Criterios e Indicadores del Manejo Forestal Sostenible para las unidades silvícolas muestra un favorable comportamiento, así los puntos de cortes determinados demuestran que las cualidades de los sistemas de Criterios e Indicadores creados se califican como altamente relevante en la segunda circulación y el Coeficiente de Concordancia es de 0.81, según se observa en tabla 2., lo que permite inferir que existe una adecuada coincidencia de criterios entre los expertos. 180 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Tabla 2. Resultados del cálculo de los puntos de cortes y del Coeficiente de Concordancia de Kendall en la segunda circulación para los sistemas de Criterios e Indicadores en opinión de los expertos. Criterios C-1 C-2 C-3 C-4 SUMA P N-P Categoría. 1 0,6154 3,9000 3,9000 3,9000 12,3154 3,0789 -0,6478 A 2 0,0966 3,9000 3,9000 3,9000 11,7966 2,9492 -0,5181 A 3 0,5024 3,9000 3,9000 3,9000 12,2024 3,0506 -0,6196 A 4 0,1938 3,9000 3,9000 3,9000 11,8938 2,9735 -0,5424 A 5 0,8677 3,9000 3,9000 3,9000 12,5677 3,1419 -0,7109 A Puntos de corte 0,5690 4,8750 4,8750 4,8750 60,7759 2,4310 Coeficiente de Concordancia de Kendall 0.81 Fuente: Elaborada por los autores. La propuesta del sistema de Criterios e Indicadores consta de los siguientes principios, en coorcondancia con la Guía de Campo para la Validación de Criterios e Indicadores para la Ordenación Forestal Sostenible a nivel de Unidad de Manejo, en Hondura, 2005 1. Principio I. La planificación, la estructura normativa y un marco institucional son factores que favorecen el Manejo Forestal Sostenible. Posee tres criterios y 17 indicadores. 2. Principio II. Mantenimiento de la integridad del ecosistema. Muestra dos criterios y 13 indicadores. 3. Principio III. Sostenible rendimiento y calidad de los bienes y servicios ambientales. Abarca tres criterios y 22 indicadores 4. Principio IV El manejo forestal sostenido mantiene o realza el acceso equitativo intergeneracional a los recursos y beneficios económicos. Contiene tres criterios y 21 indicadores. Conclusiones 1. La Guía de Procedimiento de los indicadores operacionaliza el proceso de toma de datos y registro de los mismos de manera estandarizada y sistematizada. 2. La propuesta de sistema de Criterios e Indicadores del Manejo Forestal Sostenible para las unidades silvícolas de la Empresa Forestal Integral “Cienfuegos” muestra cuatro principios, 11 criterios y 73 indicadores que comprenden las dimensiones de las sostenibilidad. 3. Los puntos de cortes determinados demuestran que las cualidades de los Criterios e Indicadores creados se califican como altamente relevante con un Coeficiente de Concordancia de 0.81. Bibliografía. Bueren, EL van y Blom E. 1997. Esquema jerárquico para la formulación de estándares para el manejo forestal sostenible. Holanda, 54 p. http://www.piec.org/pathfinder/Pathfinder_portal/Instrumentos_Esp/A3/Print/A3-Esq-jerarquico-P.pdf. [Consulta: 22 de enero del 2007]. Cué G. JL., Morejón G. M., Pino E. I. y Herrero E. JA. 2008. Adecuación del Barómetro de Sostenibilidad a la valoración de los Criterios e Indicadores de Manejo Forestal Sostenible en las unidades silvícolas. Revista Electrónica “AVANCE”, CITMA, Pinar del Río, Cuba, http://www.ciget.pinar.cu/Revista/No.20112/articulo.htm [Consultado: 12 septiembre del 2013]. 181 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Cué G. JL., Morejón G. M., Pino E. I., León G. M. C., Cuellar G. M. y Herrero E. JA. 2008. Propuesta de Criterios e Indicadores de Manejo Forestal Sostenible en las unidades empresariales de base de la Empresa Forestal Integral “Cienfuegos”. Revista Electrónica “AVANCE”, CITMA, Pinar del Río, Cuba. http://www.ciget.pinar.cu/Revista/No.2011-2/articulo.htm [Consultado: 12 septiembre del 2013]. Farsari Yianna y Prastacos P. 2002. Sustainable Development Indicators: An overview, 16 p. En: http://www.iacm.forth.gr/papers/Asteras-English.pdf [Consulta: 5 de junio del 2005]. Gómez O., D. 2007. “Evaluación Ambiental Estratégica”. Mundi-Prensa Madrid 2007. Centro de Estudios y Experimentación de Obras Públicas, II Curso sobre "Evaluación Ambiental Estratégica: Un Instrumento para el Desarrollo Sostenible", Madrid, 3 al 6 de marzo del 2008. Guía de campo para la Validación de Criterios e Indicadores para la Ordenación Forestal Sostenible a nivel de Unidad de Manejo. 2005 Hondura. En: http://www.agendaforestalhn.org/documentos/C%20e%20I/Gu_a_metodol_gica_CeI.doc [Consulta: 17 de enero 2007]. Herrero E., JA. 2005. Criterios e indicadores de manejo forestal sostenible. Una visión de futuro. Agrinfor. Ministerio de la Agricultura, La Habana, Cuba. 55 p. Instituto de la Ingeniería de España. 2001 El método Delphi. http://www.gtic.ssr.upm.es/encuestas/delphi.htm [Consulta: 27 de octubre del 2005]. 6p. En: Oficina Federal de Agricultura. 2002. Valoración de la Sostenibilidad. En: Informe Agrario de la Oficina Federal de Agricultura. Extracto, Suiza. 12 p. Olabe A. 2003. Indicadores de sostenibilidad en el ámbito de la responsabilidad social de la empresa. [en línea]. http://www.grupcies.com/ [Consulta: 12 de enero del 2004] Pinter L. 2002. Seeking a true measure of sustainable developmente. [en línea]. Bulletin. 11(2) En: http://bulletin.rec.org/bull112/coversearch2.html [Consulta: 13 de marzo del 2005]. Prescott-Allen, R. 1997. Barometer of Sustainability: Measuring and communicating wellbeing and sustainable development. Cambridge: IUCN, En: “Sustainable development: presenting the main measurement methods”. Van Bellen, H. M. Ambient. Soc. [online]. ene./jun. 2004, 7(1):67-87. En: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1414-753X2004000100005&lng=es&nrm=iso>. [Consulta: 05 Junio 2005] Quiroga, R. 2003. Indicadores de sustentabilidad. Experiencia mundial y desafíos para América Latina. Presentación de Power Point. Documento de Trabajo. CEPAL. En: Curso: Desarrollo Sustentable Unidad 2. Evaluación de la sustentabilidad e indicadores de desarrollo sustentable. En: http://agenda.di.uaslp.mx/htms/biblio_U2.htm [Consulta: 28 de noviembre del 2006]. Rusch V. y Sarasola M., (2005) Indicadores de biodiversidad en bosques Nothofagus. En. Revista IDIA 21(8):8-14 http://www.inta.gov.ar/ediciones/idia/forest/dinamica01.pdf [Consulta: 12 de febrero 2007]. Socorro, A. 2002. Indicadores de la sostenibilidad de la gestión agraria en el territorio de la provincia Cienfuegos”. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas, Ciudad Habana, Cuba, 96 p. Yurjevic, A. 1996. El desarrollo sustentable: una mirada actualizada. CLADES – Chile Revista de CLADES No. 10 En: http://www.clades.cl/hacemos/10/rev10art2.htm [Consulta: 17 de enero 2005]. 182 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 EFECTO DE VARIABLES CLIMÁTICAS Y LITOLÓGICAS SOBRE EL CRECIMIENTO EN ALTURA DOMINANTE EN MASAS DE Pinus nigra Arn Vera, R.1; Bravo-Oviedo, A.1, 2; Rio, M.1, 2; Bravo, F.1; Montero, G.1, 2 1Instituto Universitario de Investigación en Gestión Forestal Sostenible Universidad de Valladolid-INIA. Avda. Madrid 44, 34071, Palencia, España. 2Departamento de Sistemas y Recursos Forestales. INIA-CIFOR. Autopista A6 Madrid-La Coruña, Salida 8. Madrid, España [email protected] Introducción Diversos factores bioclimáticos condicionan el crecimiento de una masa forestal. La búsqueda de una función matemática que se ajuste adecuadamente a esta variabilidad es el objeto de estudio de muchos modelizadores para ofrecer conclusiones cercanas a la realidad sobre una determinada gestión. En ese sentido, el ajuste de un modelo a una serie de datos de crecimiento de un determinado sistema forestal puede involucrar áreas de distribución amplias según la especie en estudio. En estos casos, el nivel de precisión puede variar en función de la calidad del sitio y su resultado puede ser una curva no adecuada. Esta variabilidad natural es causada en parte por el clima y suelo, por lo que su incorporación en los diferentes modelos podría resultar en un mejor ajuste. El uso de modelos dinámicos derivados por aproximación de diferencias algebraicas generalizadas GADA (Cieszewski & Bailey, 2000 conjuntamente con la incorporación de los factores causantes de la variabilidad en la estructura de cada masa pueden generar curvas con mayor especificidad y realismo. Para comprobar esta hipótesis, en el presente estudio se han incorporado variables climáticas y litológicas específicas de cada parcela en las ecuaciones GADA de crecimiento en altura dominante ajustadas a un conjunto de datos de distribución natural amplio de Pinus nigra Arn. Con ese antecedente, se ha planteado el objetivo de determinar si la inclusión de la variabilidad climática y litológica en los modelos de crecimiento en altura dominante, mediante el uso de una ecuación dinámica derivada por diferencias algebraicas generalizadas (GADA), puede mejorar el comportamiento de los mismos en el rango de distribución natural de Pinus nigra Arn en España. Metodología Los datos utilizados provienen de parcelas permanentes ubicadas en masas de Pinus nigra Arn que se encuentran dentro del rango de distribución natural en España. Son parte de la red de parcelas de producción del INIA-CIFOR que han sido inventariadas de forma sincrónica desde 1963, abarcando de esta manera un rango de edades entre 20 a 120 años en distintas calidades de estación. Las parcelas están ubicadas en Segura-Cazorla, Serranía de Cuenca, Els Ports y Sierra de Gúdar-Javalambre, y específicamente están asentadas en las provincias de Lérida, Tarragona, Cuenca, Guadalajara, Teruel y Jaén. Coinciden con las procedencias de Prepirineo Catalán, Alto Maestrazgo, Sistema Ibérico y Cordilleras Béticas (Catalán et al 1991). La información tanto de la altura dominante, edad, clima y origen litológico, fue proporcionada por el Centro de Investigación Forestal del Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA-CIFOR) para un total de 46 parcelas. El criterio de Assmann es considerado para determinar la altura dominante (Assmann, 1970). Sus edades varían por localidad, oscilando entre 31 y 175 años, mientras que las alturas dominantes varían de 7,2 a 30,5 m. Las características climáticas varían con cada parcela con rangos que varían de 549 a 1272 de precipitación media anual y 8,3 a 13ºC de temperatura media anual (AEMET, 2011). Por otra parte, existe una 183 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 tendencia del pino laricio en ubicarse sobre orígenes litológicos compuestos por dolomías, calizas y margas, y con especial preferencia de calizas + dolomías con un 48% de concentración de las masas12. Los modelos de Hossfeld, Korf, Richard, Weibull y Levakovic, seleccionados en Kiviste (2002) y el modelo de Lundqvist (1957) fueron derivados a 8 modelos dinámicos por diferencias algebraicas generalizadas GADA (Cieszewski & Bailey, 2000). En una primera fase estas ecuaciones se ajustaron al grupo de datos de las parcelas con el fin de seleccionar el mejor modelo para posteriormente incluir las variables ambientales. La relación entre los parámetros y el factor X de intensidad de crecimiento fueron asumidas según Cieszewski (2004) y Cieszewski & Strub (2008) Se introdujo un proceso autoregresivo de orden p (1), en función de los residuos del modelo ajustado con y sin un retardo de tiempo p, cuya estructura es la siguiente (Gregorie et al. 1995): (1) Donde es el error experimental que sigue un proceso autorregresivo, es 1 cuando j > p y es el parámetro autoregresivo continuo donde k=tij-tij-p y tj>tj-p para todo i. La selección del mejor modelo se realizó en función de los estadísticos de ajuste en términos de precisión y sesgo. Para determinar la precisión, se calculó el error de la predicción media absoluta (2) y el error del cuadrado medio de la predicción (3), para el sesgo se calculó el error promedio de la predicción de cada modelo (4), adicionalmente se determinó la eficiencia de cada modelo (5). (2) (3) (4) (5) Donde es la predicción residual, y son, en su orden, los valores observados y esperados, k es el número total de observaciones. Para la segunda parte del estudio, se realizó un análisis factorial con las variables climáticas, esta técnica permite encontrar una estructura subyacente de la matriz de datos en términos de factores inobservables (Crawley, 2007), donde las variables más correlacionadas son explicadas por dichos factores, el contraste de esfericidad de Bartlett basado en la matriz de correlación muestral R, determina si el análisis factorial es adecuado (Pérez, 2008); la selección de la variable está en función de la carga individual. Por otra parte, dado que la información litológica no es de característica cuantitativa, esta fue analizada de forma gráfica para observar la existencia de agrupaciones y demostrar variabilidad inter parcelas. Para ello se realizó un gráfico de la matriz de puntuaciones factoriales utilizando el método de mínimos cuadrados generalizados de Bartlett (Batlett 1937, 8) de los dos primeros factores resultantes del análisis factorial 12 Datos proporcionados por el INIA-CIFOR 184 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 con las variables climáticas y se introdujo esta información. La formación de agrupaciones determinará el origen litológico a seleccionar para ser ingresado en el modelo GADA en términos de presencia y ausencia. Para la expansión de los parámetros, se siguió el procedimiento descrito en Bravo-Oviedo et al (2008). Los modelos dinámicos fueron ajustados usando el PROC MODEL de SAS/ETS v 8 para Windows (SAS INC., 2004) y para la evaluación se utilizó el programa estadístico R (R Core Team, 2012). Resultados De acuerdo a los resultados obtenidos, todos los modelos presentan un alto grado de precisión y bajo nivel de sesgo y su eficiencia está por encima del 98%, sin embargo los estimadores de los parámetros en su mayoría son no significativos. El único modelo que tiene todos los parámetros significativos a un nivel de significación de 0,05 es la ecuación dinámica derivada del modelo de Hossfeld (E2), por lo tanto es el único considerado para ser evaluado y ajustado con las variables climáticas y litológicas. En cuanto al sesgo, el modelo tiende a sobreestimar por debajo de los 40 años y sobre los 120 años. Mientras, la eficiencia es superior al 97% para parcelas de 40 años en adelante, aunque a edades comprendidas entre 60 y 80 años, existe una ligera pérdida de eficiencia. De manera general este modelo es bastante preciso, sobre todo debido a que las edades de las masas de las parcelas de estudio son en su mayoría superiores a 40 años. El comportamiento biológico del modelo fue evaluado mediante un gráfico de curvas de calidad (fig. 1), cuya edad de referencia seleccionada fue de 100 años y cuatro índices de sitio fueron utilizados como altura de referencia. Se comprobó que las alturas alcanzadas en dependencia de la calidad de estación eran biológicamente adecuadas, por tanto este modelo cumple con las condiciones requeridas para predecir la altura de la masa. Figura 1. Curvas de calidad para Pinus nigra Arn utilizando la ecuación dinámica expandida del modelo de Hossfeld a una edad índice de 100 años De acuerdo al análisis factorial, se determinó que un 67,2% de la variabilidad puede ser explicada con los factores 1 y 2, los cuales aportan un 40,3% y 26,8% de la información respectivamente. El primer factor se caracteriza por agrupar los regímenes de precipitación de tal manera que las variables con mayor carga de aporte son en su orden: la precipitación total, superávit, índice hídrico, la precipitación en primavera, la precipitación en invierno y la precipitación en otoño. Por su parte, el factor 2 agrupa variables climáticas relacionadas con la temperatura, y las de mayor carga son, en su orden: evapotranspiración, la temperatura media y las temperaturas de verano e invierno respectivamente, por 185 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 otra parte, cuando fue incorporada la variabilidad litológica en los componentes del análisis factorial, no evidenció agrupaciones definidas entre las parcelas, por lo que no se consideró que la litología pudiera ejercer un aporte significativo a la explicación del crecimiento en altura dominante del pino laricio. Posteriormente se procedió a seleccionar la variable climática con mayor aporte para el modelo, para ello, se consideraron según su carga en el análisis factorial. La precipitación total tiene la carga más alta en el primer factor, sin embargo, dado que esta variable está en función de la suma de las precipitaciones de cada estación (inverno, verano, otoño y primavera), fue descartada en vista de que la precipitación de verano no se correspondía con el mismo factor explicativo de la precipitación total, pero la precipitación en primavera y la de invierno son de interés, las siguientes variables que le siguen en carga a la precipitación total son el superávit y el índice hídrico respectivamente. Al final, para la generación de hipótesis entre los parámetros de la ecuación dinámica y el clima, fueron seleccionadas las variables: precipitación en primavera, precipitación en invierno, índice hídrico y superávit. Dos estructuras fueron probadas para relacionar la altura dominante y las variables climáticas, una polinómica de segundo grado y una relación no lineal; para la relación no lineal se ha utilizado el modelo de Hossfeld I Modificado, estas estructuras reemplazan el parámetro b de la ecuación dinámica E2. Las estructuras (6) y (7) representan las relaciones polinómica y no lineal respectivamente, donde representa la variable climática a incorporar, bo, b1 y b2 son los parámetros a ser estimados. Para su incorporación a la ecuación dinámica E2, el parámetro b es expandido y su notación quedaría como en (8) y (9), donde se incrementa el parámetro c, Ho y to son la altura y edad de referencia, t es la edad conocida y H es la altura a determinar siguiendo el mismo procedimiento de ajuste explicado anteriormente. (6) (7) (8) (9) Se aprecia una mejora en la estimación del modelo global con todas las relaciones y con todas las variables, especialmente en la relación polinómica de segundo grado con el índice hídrico, con el cual disminuye en un 9% la raíz del error medio cuadrático (RMSE) en comparación con la ecuación sin la inclusión de la variabilidad climática (tabla 2). Sin embargo, en lo relacionado al sesgo medio ( ) y eficiencia globales, no se observa una mejora significativa. El índice hídrico en relación polinómica se muestra como la variable climática más adecuada, y su inclusión mejora también la estimación del modelo por clases de edad, tanto en sesgo (fig 2) como en eficiencia (fig 3). 186 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Tabla 2. Estadísticos de ajuste globales con las variables climáticas incluidas en la ecuación dinámica E2 Tipo de relación Polinómica de segundo grado Variable incluida en E2 RMSE %RE R-Ad-adj mse Eficiencia Índice Hídrico 0,4724 9 0,9901 0,0112 0,3288 0,1846 0,992 Prec. en Primavera 0,4837 7 0,9896 0,0095 0,3336 0,1936 0,991 Prec. en Invierno 0,4985 4 0,9890 0,0099 0,3441 0,2056 0,991 Superávit 0,4938 5 0,9892 0,0114 0,3380 0,2017 0,991 Índice Hídrico 0,4955 4 0,9891 0,0104 0,3290 0,2039 0,991 Prec. en Primavera 0,4999 3 0,9889 0,0095 0,3398 0,2075 0,991 Prec. en Invierno 0,5006 3 0,9889 0,0099 0,3392 0,2081 0,991 Superávit 0,4950 4 0,9892 0,0111 0,3311 0,2035 0,991 0,9881 0,0138 0,3417 0,2237 0,990 No lineal E2 Sin inclusión de variables climáticas 0,5179 0.98 0.97 0.96 Eficiencia 0.00 Sin variables climáticas Inclusión del Índice Hídrico 0.95 -0.05 Sesgo 0.05 0.99 0.10 1.00 %RE: Porcentaje de reducción en RMSE con la inclusión de la variable climática 40 60 80 0.94 -0.10 Sin variables climáticas Indice Hídrico 100 40 120 60 80 100 120 clases de edad (años) clases de edad (años) Figura 2. Sesgo por clases de edad con el índice hídrico incluido Figura 3. Eficiencia por clases de edad con el índice hídrico incluido Discusión De las 8 ecuaciones dinámicas derivadas de los modelos de Korf, Lundqvist, Hossfeld, Weibull, Richards y Levakovic II por procedimiento GADA (Cieszewski & Bailey, 2000), el mejor ajuste resultó con la ecuación dinámica del modelo base de Hossfeld. Esta ecuación dinámica derivada también describe mejor el crecimiento en altura para Pinus pinaster Ait (Bravo-Oviedo et al 2007), pero difiere de la ecuación encontrada por Martin-Benito et al (2008) para Pinus nigra, ya que en sus resultados indican que la ecuación con mejor ajuste para esta especie resultó ser la de Lundqvist (1957). Tanto el modelo de 187 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Lundqvist como el de Hossfeld son bastante buenos para predecir la altura dominante de Pinus nigra, sin embargo, la ecuación dinámica derivada de Hossfeld tiene una mayor eficiencia global y por clases de edad. Existe evidencia que indica que el patrón de crecimiento en altura dominante es afectado por las condiciones climáticas de cada sitio y que tiene que ver con el índice hídrico anual. Su inclusión mejora la explicación del modelo, sobre todo en clases de edad menores a 40 años y entre los 60 y 80 años. Esta variable climática está relacionada con la cantidad de agua disponible en función de la precipitación y la evapotranspiración; la relación encontrada entre esta variable climática con la respuesta del pino laricio en altura dominante, podría deberse a una relación más estrecha entre la precipitación, las características físicas del suelo y la sequía estival característica de climas mediterráneos, sin embargo, la diferenciación entre sitios no parece ser adecuadamente explicada por el sustrato sobre la cual están asentadas lo cual probablemente se deba a que la información disponible de orígenes litológicos a escala amplia determina que un 81% de individuos se establecen sobre terrenos con predominancia de calizas+dolomías, dolomías y calizas+margas. Esta gran agrupación de parcelas podría ser la causa de la insuficiente explicación de esta variabilidad en el modelo; información sobre características físico-químicas específicas de cada parcela podrían resultar en mejor detalle y con mayor aporte a la explicación tanto de la altura dominante. La altura dominante parece ser explicada adecuadamente con ecuaciones dinámicas, sin embargo, los resultados de este estudio indican que esta explicación es mejor para Pinus nigra que los encontrados para Pinus Pinaster por Bravo-Oviedo et al (2007) con la misma metodología GADA, esto podría deberse a que la variabilidad en el pino marítimo mediterráneo es mayor que en el pino laricio, lo cual se demuestra cuando el clima es incluido en el modelo, mejorando notablemente la explicación en todas las clases de edad en Pinus pinaster (Bravo-Oviedo et al 2008), cosa que no se logra en todas las clases de edad en Pinus nigra. La importancia de la predicción adecuada de la altura dominante mediante el uso de modelos de crecimiento, radica en observar las distintas tendencias del crecimiento de las masas, sobre todo cuando las variables climáticas son incluidas, esto permitiría por una parte, evaluar los posibles impactos del cambio climático y orientar a gestores sobre sus efectos probables en la gestión (Bravo, 2009) y por otra parte tomar decisiones adecuadas dirigidas a contribuir en la solución de este problema. Conclusiones Diferentes valores de humedad específicos de cada sitio afectan el crecimiento del pino laricio, y permite explicar adecuadamente las diferencias en altura dominante a lo largo de su distribución natural en España mediante la utilización de la ecuación dinámica de Hossfeld derivada por el procedimiento de diferencias algebraicas generalizadas. Por otra parte, aunque la información litológica no ofrece ninguna ventaja para la estimación de la altura dominante del Pinus nigra, las características físicas específicas del suelo podrían ser una fuente de variabilidad que mejorarían la explicación de su desarrollo. Bibliografía AEMET. 2011. Iberian climate atlas. Agencia Estatal de Meteorología (España) and Instituto de Meteorología (Portugal), Madrid, Spain. 188 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Assmann, E. 1970. The Principles of Forest Yield Study, English Edition. Pergamon Press Ltd. 506. Oxford. Bartlett, M. S. 1937. The statistical conception of mental factors. Brit. Jour. of Psych. 28, 97– 104. Bartlett, M. S. 1938. Methods of estimating mental factors. Nature, 141, 609–610. Bravo, Andrés. 2009. Variabilidad del crecimiento en altura dominante de Pinus pinaster Ait., en el interior peninsular. Tesis Doctoral por la Universidad de Valladolid. España. Bravo-Oviedo, A.; Rio, M.; Montero, G. 2007. Geographic variation and parameter assessment in generalized algebraic difference site index modeling. For. Ecol. and Manag. 247. 107–119 Bravo-Oviedo, A.; Tome, M.; Bravo, F.; Montero, G.; Rio, M. 2008. Dominant height growth equations including site attributes in the generalized algebraic difference approach. Can. J.For. Res. 38, 2348-2358. Catalán, G.; Gil, P.; Galera, R.; Martin, S.; Agundez, L.; Alia, R. 1991. Las regiones de procedencia de Pinus sylvestris L. y Pinus nigra Arn. Subsp. salzmannii (Dunal) Franco en España. ICONA. Cieszewski, C.J. 2004. GADA Derivation of Dynamic Site Equations with Polymorphism and Variable Asymptotes from Richards, Weibull, and other Exponential Functions. Plant. Manage. Res. Coop., p. 10. Cieszewski, C.J.; Bailey, R.L. 2000. Generalized Algebraic Difference approach: theory based derivation of dynamic site equations with polymorphism and variable asymptotes. For. Sci. 46, 116–126. Cieszewski, C.J; Strub, M. 2008. Generalized Algebraic Difference Approach Derivation of Dynamic Site Equations with Polymorphism and Variable Asymptotes from Exponential and Logarithmic Functions. For. Sci. 54, 303–315. Crawley, M. 2007. The R book. Wiley. 950. England Gregorie, T.G.; Schabenberger, O.; Barret, J.P. 1995. Linear modelling of irregularly spaced, unbalanced, longitudinal data from permanent-plot measurements. Can. J. For. Res. 25, 137–156. Kiviste, A.; Álvarez-González, G.; Rojo-Alboreca, A.; Ruiz-González, D. 2002. Funciones de crecimiento de aplicación en el ámbito forestal. Ministerio de Ciencia y Tecnología, INIA. 190. Madrid. Lundqvist, B. 1957. On the height growth of cultivated stands of pine and spruce in Northern Sweden. Fran Statens Skogforsk. band 47. 1 – 64. Martín-Benito, D.; Gea-Izquierdo, G.; Rio, M.; Cañellas, I. 2008. Long-term trends in dominant-height growth of black pine using dynamic models. For. Ecol. Manag. 256. 1230–1238. Pérez, C. 2008. Técnicas de Análisis Multivariante de Datos. Pearson Educación. 672, Madrid R Core Team. 2012. A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna. SAS Institute Inc. 2004. SAS/ETS 9.1 User’s Guide. SAS Institute Inc., Cary, NC, USA. 189 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 ADS80 AND MODIS BASED AND REMOTE SENSING APPLICATIONS IN FOREST MANAGEMENT Tomislav Sapic Faculty of Natural Resources Management, Lakehead University, Thunder Bay, Canada 955 Oliver Rd., Thunder Bay, Ontario, P7B 5E1, Canada, [email protected] Introduction A Geographic Information System (GIS) is today a main component of forest management in Canada and its role continues to broaden. The role of GIS has already been broadened in large part through the increasing addition of digitized, pre-processed, aerial imagery to GIS databases, while one of the newest expansions of GIS is the area of GIS Web services, in particular Web-delivered, near real-time, satellite imagery derivatives. In the province of Ontario, Leica Airborne Digital Sensor (ADS), aerial imagery has been commissioned by the provincial government and integrated into forest management province-wide, while MODIS satellite based derivatives, delivered near real-time as Web services are emerging as practical GIS forest management data sources. The use of Leica ADS aerial imagery and MODIS data in forest management in Ontario and in teaching at the Faculty of Natural Resources Management, Lakehead University, as well as their potential uses in the context of forest management in Ecuador, will be the focus of this paper. Leica ADS Imagery in Ontario Forest Management Ontario is the largest province in Canada, 107.6 M ha in total area (OMNR 2011a), within which regulated forest management is carried out in Forest Management Units (FMUs) inside an area called Area of Undertaking -- a wide band stretching east - west through the central portion of Ontario and covering 43.8 M ha (Fig. 1). Provincial policy is to create for each FMU a new aerial-imagery-based forest resource inventory every 10 years (OMNR 2013). Figure 1. Forest Management Units within the province of Ontario Leica ADS sensors have been developed over several technological generations (Leica Geosystems 2013). Leica ADS40 preceded Leica ADS80, which in turn is followed by Leica ADS100. In 2006, Ontario 190 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Ministry of Natural Resources commissioned North West Geomatics to produce a set of imagery and geomatics products for the area of all FMUs within the AOU, and extending out of it, adding up to 50 M ha (North West Geomatics 2013). At the time the undertaking was the largest of its kind in the world. The technology used for the project was at first ADS40 and was subsequently replaced by ADS80. The products delivered and made available to forest management tenure holders were orthorectified tiles including panchromatic 0.2 m (GSD) and multispectral (RGB + NIR) 0.4 m (GSD); filmstrip stereo imagery at 0.2 m (GSD) for panchromatic and 0.4 m (GSD) for multispectral (RGB + NIR) , and a 5 m point grid Digital Surface Model (DSM) with points image-classified into three land type classes: deep water, bare ground, and vegetation. Horizontal accuracy for the imagery and the derived DSM is ±1 m (LIO 2013).The project took four years and cost C$25 M (Industry Canada 2013). ADS80 has a CCD sensor recording electromagnetic reflectance along a 12-bit radiometric scale. The CCD sensor head SH92 contains two 4-band beamsplitters, one in nadir and one at 16º backwards. In total there are 12 CCD lines, each containing 12,000 pixels size 6.5 μm: two lines are single panchromatic lines, one pair of panchromatic lines is staggered by half a pixel, and out of the eight remaining lines, two are recording red, two green, two blue and two near-infrared (Leica Geosystems 2013). ADS80 is equipped with an Inertial Measurement Unit (IMU); a gyro-stabilized mount with a stabilization range in roll -7º - +7º, in pitch -8º - +6º, and in drift -30º - +30º (Leica Geosystems 2008); a control and data processing unit that includes the Inertial Positioning Attitude System and a Global Navigation Satellite System (GNSS) receiver; the Mass Memory unit that can store up to 980 GB over up to 9.7 hours of flying (Figs. 2, 3, and 4); and the Flight and Sensor Control Management System (FCMS) (Figs. 5 and 6) that can be used to create a flight plan, while it also monitors and reports on the execution of the flight plan, monitors and controls the sensor, and monitors GNSS quality. All the ADS80 elements together make up a sensor system that in a short time can scan large areas with high spatial and radiometric resolution large-format stereo images, at high accuracy and ready for a rapid post-processing and a creation of stereomodels, orthophotography, and DSM models (Leica Geosystems 2013). Figure 2. Onboard setup of a Leica ADS system (Leica ADS40 in this case): 1 - Camera head; 2 - Control Unit; 3 - Mass Memory; 4 - Operator interface; 5 - Pilot guidance indicator; 6 - Gyro-stabilized mount (Fricker 2001). 191 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figure 3. A Leica ADS system, ADS40, on board of a Cessna Caravan plane (Leica Geosystems 2008). Figure 4. Leica ADS80 system, with 1 - Camera, 2 - gyro-stabilized mount, 3 - Control Unit, and 4 – Operator interface (Leica Geosystems 2008). Figure 5. An example of an FCMS, north-up view display (Leica Geosystems 2011). 192 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figure 6. An example of an FCMS, nose-up, pilot view display (Leica Geosystems 2011). Imagery created by ADS80 is processed over three levels, L0, L1 and L2. L0 represents raw, unrectified imagery that is usually not delivered to the client. L1 represents stereo multispectral imagery with a 100% overlap but taken at two different angles, nadir and 16º backwards (Fig. 7), and ready to be used in a stereo model, and L2 represents orthorectified imagery. Figure 7. Pushbroom scanner ADS80 recording of nadir and 16º backwards multispectral imagery (Leica Geosystems 2008). ADS80 L1, stereo imagery for the Ontario FRI was delivered in filmstrips (Fig. 8) and L2, orthorectified, imagery in 5 x 5 km tiles (Fig. 9). Both, L1 and L2 imagery were delivered in panchromatic and R+G+B+NIR bands (Figs 10, 11, and 12). In addition, Digital Surface Model (DSM) points were extracted from the stereo, L1, models into a 5 m grid and classified (Fig. 13). The DSM is different from the Digital Terrain Model, in that it represents the surface seen from above (e.g., tree canopy, building top, ground) rather than only the ground (terrain). As a result, each DSM point is assigned the surface elevation and a class as bare ground, vegetation, or open water. In total, 280 TB of data were delivered for the entire area scanned for the Ontario FRI (Leica Geosystems 2008). 193 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 a) b) Figure 8. An example of ADS filmstrip imagery taken in Ontario. a) the geographic location of the filmstrips , b) the filmstrip images themselves, indicating overlapping flightlines (Leica Geosystems 2008). Figure 9. An ADS orthorectified, 5x5 km tile, in true colour. 194 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figure 10. ADS orthorectified image of the Lakehead University campus, in true colour. Figure 11. ADS orthorectified image of the Lakehead University campus, in false colour. Figure 12. Details in an ADS orthorectified image of the Lakehead University campus. 195 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figure 13. ADS DSM 5x5 m points classified in bare earth (red), vegetation (green), and deep water (blue). ADS L1 stereo imagery is used in Ontario in stereomodels mainly for FRI-based tree species and ecosite identification, but increasingly non-FRI uses have been found, such as in road layout, mining, terrain analysis, and so on. FRI, being a GIS vector dataset, is created by bringing ADS stereomodels into the GIS environment, through, for example, an addition of a suitable stereo-viewing and editing extension to ArcGIS. Here, stands are delineated into polygons based on tree species, forest structure, and ecosites, and average tree heights measured (Fig. 14). In doing so, the NIR channel-based false-colour display has become the display of choice for most of photointerpreters, allowing them to more easily distinguish conifer from deciduous trees. At the same time, other, non-forested areas, are delineated into specific ecosite and land classes, such as various wetland types, rocky outcrops, roads, residential areas, etc., also by relying on stereo-viewing. Open water is delineated through image classification of L2, orthorectified, images. Ontario FRI is produced for each FMU at the turn of its 10-year FRI cycle. Figure 14. A person stereo-viewing aerial imagery (OMNR 2011b). The DSM point, GIS, layer is used to orthorectify L1 images, bring the z (height) axis in the stereomodels and enable height (e.g. tree height) measurements, and is available for any terrain (surface) based analysis. As well, since each point is classified into one of the three aforementioned classes, the DSM layer can also be used for broad land-type analyses. However, based on a small sample inspection, DSM points delivered for FMUs early in the four-year project contained lower classification accuracy and lower height accuracy around the edges of buildings and forest canopies. Lower classification accuracy was 196 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 exhibited when dark, tree-shaded, areas would be misclassified as deep-water (Fig. 13). Lower height accuracy, would occur when points’ z values would not follow abrupt changes of the surface height, e.g., along forest canopy edges, but would gradually change, causing a) misses of small canopy openings and b) gradual taper-off of height values within 15 m of vertical objects 18 to 22 meters high (Lusignan 2011). Lower accuracy of ADS derived DSM points around tree and building edges were reported by Gehrke et al. (2010). ADS Imagery Advantages and Potentials in Forest Management Province of Ontario has traditionally relied on aerial, 9x9 inch, 1:20,000 scale, analog photos to estimate the FRI. The Ontario ADS imagery coverage has now changed to state-of-the-art aerial imagery that is at higher (practical) spatial resolution; it includes the NIR channel; it allows digital viewing and manipulation -including image classification -- thus enormously shortening the post-processing time; and it is a readily available source of a high resolution and high accuracy DSM. Likely because of the great size of the scanned area in Ontario, the improvements have been accomplished at a low per-area-unit cost of ~C$0.50 per ha. Besides the savings in cost and time, advantages of the new imagery are several: a) higher spatial resolution, viewed and manipulated within computer stereo-viewing systems will enable photo interpreters to produce FRI of much higher accuracy; b) availability of the NIR band will allow for a better tree species identification when viewed in false colour; c) already, the imagery is yielding a much more accurate and precise classification and digitization of basic features such as lakes and roads (Fig. 15); and d) the high accuracy DSM layer is expected to lead to higher accuracy in tree height estimates. Figure 15. An example of a higher precision and accuracy of a GIS feature based on the ADS imagery. The blue line represents the outline of a GIS water feature created based on the new ADS imagery; the purple line represents the old, pre-ADS imagery, GIS water feature. Beyond the advantages of ADS imagery, a large potential advantage can also be to use the DSM as an alternative to LIDAR, the state-of-the-art source of DTM today. This emergence of image-based DSM as an alternative to LIDAR has been accomplished mainly through the recent development of the Semi Global Matching computer algorithm that computes a very accurate three-dimensional (x,y,z) position for each point in the images involved in a photogrammetric stereopair (Gehrke et al. 2010, White et al. 2013). The alternative can be potent in particular in areas with no or with very sparse forest coverage. For forested areas with a full canopy, elevations measured from stereo imagery, i.e., DSM, represent only the canopy heights and not the ground heights underneath. In contrast, LIDAR elevations provide both due to the capability of LIDAR pulses to penetrate the canopy through small openings (White et al. 2013). Still, when it comes to non-forested areas, image-based DSM elevations are comparable to LIDAR based DTM elevations, with average differences reported by Gehrke et al. (2010) at 0.8 ± 5.4 cm in height estimates 197 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 for a semi-urban, generally flat, area with sporadic trees. On the opposite side of the terrain spectrum in a high alpine terrain, likely exhibiting similarities with the mountainous regions of Ecuador, Bühler et al. (2012) measured image-based DSM accuracy at < 0.5 m RMSE for slope inclinations < 50º and rapidly decreasing for steeper slopes, dropping to Q95 > 10 m for slope inclinations > 70º. Such high accuracy scores of image-based DSM might make the cost of ADS imagery more acceptable, if it turns out that expensive LIDAR data acquisition is not required, or at least not for areas without full tree canopy. MODIS Data in Forest Management MODIS, Moderate Resolution Imaging Spectroradiometer, is a sensor on board of the NASA’s Earth Observing System’s Terra and Aqua satellites. Through MODIS, the entire Earth’s surface is viewable every 1 - 2 days and 12 bits images are acquired in 36 spectral bands ranging in wavelength from 0.4 μm to 14.4 μm and ranging in spatial resolution from 250 m to 1000 m (NASA 2013). MODIS data are available for free and provided through 65 data products, from electromagnetic reflectance values to derived environmental and vegetation variables, and in terms of temporal representation, from dailies (i.e., near real-time data) to up to a year n-day composites (NASA LP DAAC 2012, 2013). Included in the set of derived environmental and vegetation variables are products identifying active fire locations and vegetation indices. The active fire location products, MOD14 and MYD14 are derived from MODIS 4- and 11-micrometer radiances (NASA LP DAAC 2013a), based on the Thermal Anomalies and Fire algorithm, and indicate detections of 1 or more fires within a 1 km pixel. These active fire location products are managed by the Fire Information and Resource Management System (FIRMS), which provides them in several different file formats and as near real-time or archived data (EOSDIS 2013). The file formats include shapefiles, KML and text files, and WMS (Web Map Service). FIRMS active fire KML (Keyhole Markup Language) files are point derivatives, each point positioned in the centre of a 1x1 km pixel and representing a detected fire or multiple fires within the pixel. FIRMS points are described with the time and confidence level of the fire detection. FIRMS KML fire files are daily updated and can be added as a map service to any GIS Web service enabled map around the globe, or the archived KML files can be downloaded, parsed and analyzed as point fire data. MODIS vegetation indices (VI), represented by MOD13 and MYD13 MODIS products, provide consistent comparisons of vegetation conditions, and are intended for global monitoring of vegetation condition and analyzing global biogeochemical and hydrologic processes, but also for describing local, land surface biophysical properties and properties, including primary production and land cover conversion (NASA LP DAAC 2013b). The VI are derived from 469 nm, 645 nm, and 858 nm bands and are provided as 16-day or monthly averages, at spatial resolutions from 250 m to 5600 m. Due to their frequency, regularity and availability cost-free, VI have been recognized as very useful in models assessing impact of forest management on water quality, as demonstrated in the study by Li et al. (2008). Summary and Conclusions Continuing technological developments in aerial cameras, aerial and satellite imagery acquisition and processing, as well as Web GIS service standards, have made it possible for image and image-based data to be efficiently integrated into forest management. These image data can be on one hand very accurate and precise, such as the case with Leica ADS imagery-based data, and on the other hand, temporally consistent and frequent, down to near real-time. Apart from being used to create more accurate and precise forest management core information, these image-based data can be used to analyze the impact of forest management on the environment in which it takes place. As well, image-based data such as DSM can be used to create highly accurate terrain models (albeit outside of full tree canopy) and potentially used in hydrological and perhaps climate-change directed glacial analyses. 198 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 At the Faculty of Natural Resources Management, Lakehead University, Ontario FRI ADS80 imagery and DSM is used in teaching GIS, Photogrammetry and Remote Sensing, and in research. Students are taught on a) how to use the imagery in a GIS and stereo environment (Fig. 16), b) how to manipulate the imagery and its bands to most suitable and effective colour display for photointerpretation; c) how to recognize tree species in the imagery; and d) how to recognize ecosites and use this knowledge in creating an FRI according to Ontario’s guidelines. 1a 3 1b 2 Figure 16. ERDAS Stereo Analyst extension in ArcMap 10.0 (Intergraph 2013, ESRI 2013), used for teaching at Lakehead University. 1a&1b - left and right L1 ADS filmstrip images; 2 – left and right L1 ADS filmstrip images in a 2D display; 3 -- left and right L1 ADS filmstrip images in a 3D, stereo display. MODIS FIRMS KML active fires points are used as a Web GIS service and displayed within Web GIS applications and made available to students and researchers for their use (Fig. 17) 1 2 Figure 17. A Web GIS application using Google Maps (Google 2013) as the background and displaying 1 -FIRMS current fires and 2 -Ontario FMUs. (Sapic 2013). 199 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Acknowledgment The author deeply thanks Dr. Brian McLaren from Lakehead University for his encouragement in writing the paper and his greatly helpful and crucial revision of it. Literature Cited Bühler Y., Ginzler C, and Marty M. 2012. High Resolution DEM Generation in High-Alpine Terrain Using Airborne Remote Sensing Techniques. Transactions in GIS 16(5):635–647. EOSDIS (NASA’s Earth Observing System - Data and Information System). 2013. Near Real-Time Data FIRMS (Fire Information for Resource Management System). https://earthdata.nasa.gov/data/near-realtime-data/firms . November 1, 2013. ESRI. 2013. ArcGIS Desktop 10. http://resources.arcgis.com/content/arcgisdesktop/10.0/about. November 4, 2013. Fricker P. 2001. ADS40 – Progress in digital aerial data collection. Photogrammetric Week 01, Institute of Photogrammetry. Heidelberg, Germany. Gehrke, S., Morin, K., Downey, M., Boehrer, N., and Fuchs, T. 2010. Semi-global matching: An alternative to LIDAR for DSM generation. In Proceedings of the 2010 Canadian Geomatics Conference and Symposium of Commission I. Google. 2013. Google Maps. https://maps.google.com/. November 6, 203 Industry Canada. 2013. Company Information: North West Geomatics Ltd. http://www.ic.gc.ca/app/ccc/srch/nvgt.do?lang=eng&prtl=1&sbPrtl=&estblmntNo=123456112610&profile=c mpltPrfl&profileId=781&app=sold October 20, 2013. Intergraph. 2013. ERDAS Stereo Analyst extension for ArcGIS. http://geospatial.intergraph.com/products/ERDASExtensionsforArcGIS/StereoAnalystforArcGIS/Details.as px. November 4, 2013. Leica Geosystems. 2008. Leica - 3rd Generation Airborne Digital Sensors: Features/Benefits for Remote Sensing & Environmental Application. http://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=1&ved=0CCoQFjAA&url=http%3A%2 F%2Fmars.jrc.ec.europa.eu%2Fmars%2Fcontent%2Fdownload%2F1176%2F6943%2Ffile%2FT1_Rohrb ach_Rohrbach-AirborneSensors-3rd_Leica.pdf&ei=35NkUvrqMaE2gWWqoCwBw&usg=AFQjCNFxEdk3vWVOSM_CAXFELGDLjceZNg&bvm=bv.54934254,d.b2I&cad=rja . October 10, 2013. Leica Geosystems. 2011. Product description Leica FCMS Release 3.3. http://www.leicageosystems.us/downloads123/zz/airborne/general/documentations/FCMS_Product-Descripton_EN.pdf. November 1, 2013. Leica Geosystems. 2013. Leica ADS80 Airborne Digital Sensor. http://www.leica-geosystems.com/en/Leica-ADS80-Airborne-Digital-Sensor_86846.htm October 20, 2013. Li X., Nour M.H, Smith D.W., Prepas E.E., Putz G., and Watson B.M. 2008. Incorporating water quantity and quality modelling into forest management. The Forestry Chronicle 84:3. Lusignan E. 2011. Ontario eFRI's digital surface model: characteristics and uses in forest management related measurements and visualizations. Undergraduate thesis. Lakehead University. 200 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 LIO (Land Information Ontario). 2013. Forest Resource Inventory Digital Aerial Imagery Metadata. https://www.appliometadata.lrc.gov.on.ca/geonetwork/srv/en/main.home. November 1, 2013. NASA LP DAAC (National Aeronautics and Space Administration Land Processes Distributed Active Archive Center). 2012. MODIS Land Products Quality Assurance Tutorial: Part - 1. 2012. NASA LP DAAC, USGS EROS Center, Sioux Falls, South Dakota, US. NASA (National Aeronautics and Space Administration). 2013. MODIS. http://modis.gsfc.nasa.gov/about/. October 29, 2013. NASA LP DAAC (National Aeronautics and Space Administration Land Processes Distributed Active Archive Center). 2013. MODIS Data Products Table. https://lpdaac.usgs.gov/products/modis_products_table. November 1, 2013. NASA LP DAAC (National Aeronautics and Space Administration Land Processes Distributed Active Archive Center). 2013a. Thermal Anomalies & Fire Daily L3 Global 1 km. https://lpdaac.usgs.gov/products/modis_products_table/myd14a1. November 1, 2013. NASA LP DAAC (National Aeronautics and Space Administration Land Processes Distributed Active Archive Center). 2013b. Vegetation Indices 16-Day L3 Global 250 m. https://lpdaac.usgs.gov/products/modis_products_table/mod13q1. November 1, 2013. Ontario Ministry of Natural Resources (OMNR). 2011a. The Forest Resources of Ontario. Queen’s Printer for Ontario. http://www.mnr.gov.on.ca/stdprodconsume/groups/lr/@mnr/@forests/docºuments/document/stdprod_0929 22.pdf Ontario Ministry of Natural Resources (OMNR). 2011b. The enhanced forest resources inventory. Canadian Institute of Forestry Conference. Presented by Hugh Devon. Sep. 19, 2011. Ontario Ministry of Natural Resources (MNR). 2013. Forest Resource Inventory. http://www.mnr.gov.on.ca/en/Business/Forests/2ColumnSubPage/199556.html . October 20, 2013. Sapic, T. 2013. Current FIRMS fires in a Web GIS http://flash.lakeheadu.ca/~forspatial/webgis/Canada_fires_and_Ontario_FMU.html . application. White, C.J., Wulder M. A., Vastaranta M, Coops N.C, Pitt D., and Woods M. 2013. The Utility of ImageBased Point Clouds for Forest Inventory: A Comparison with Airborne Laser Scanning. Forests 4:518-536. 201 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 VIABILIDAD TÉCNICA, PRODUCTIVA, COMERCIAL Y AMBIENTAL DEL JABÓN ARTESANAL “Yakumama” ELABORADO CON PRODUCTOS FORESTALES NO MADERABLES DE LA COMUNIDAD KICHWA RÍO BLANCO Fanny Gabriela Jami Toca, Oscar Iván Navarrete Jungal Instituto Nacional Autónomo de Investigaciones Agropecuarias. Estación Experimental Santa Catalina. Programa Nacional de Forestería. Panamericana Sur Km1, sector Cutuglahua, Quito [email protected] ANTECEDENTES Los productos forestales no maderables son un recurso natural importante para el desarrollo económicosocial de las comunidades rurales (FAO 2010). Al respecto, las comunidades Kichwas de la Amazonía utilizan tradicionalmente la diversidad proveniente de los bosques para diversos usos: medicinales, artesanales, preparación de brebajes, cosmetología, culinarias. La pobreza y conservación de recursos naturales, fue el incentivo para que el INIAP (Instituto Nacional Autónomo de Investigaciones Agropecuarias) gestione y ejecute el proyecto GENFORESTAL “Conservación y Uso Sostenible de Recursos Genéticos Forestales en áreas críticas de bosques húmedos y secos de los Andes y Amazonía“, a fin de contribuir a la conservación y el uso sostenible de los recursos de los bosques en Amazonía (INIAP 2012). La comunidad kichwa Río Blanco que forma parte de las poblaciones participantes del proyecto, tomó la decisión de emprender una alternativa económica basada en el aprovechamiento de los recursos no maderables de sus bosques para la producción de jabones en base de especies como medida sustitutiva de los jabones convencionales. Para ese fin, la presente investigación contribuye al análisis de esa alternativa, mediante un estudio de mercado, cuyos resultados facilitarán la toma de decisiones por parte de la comunidad. (Miranda, J. 2005). OBJETIVOS OBJETIVO GENERAL Analizar la viabilidad técnica, productiva, comercial y ambiental del jabón artesanal amazónico “Yakumama”, elaborado con productos forestales no maderables de los bosques de la comunidad Kichwa Rio Blanco, cantón Tena, provincia Napo, que permita su desarrollo sustentable. OBJETIVOS ESPECÍFICOS Realizar el estudio de factibilidad del jabón artesanal amazónico “Yakumama” elaborado con productos forestales no maderable de los bosques de la comunidad Kichwa Río Blanco, mediante el análisis de los requerimientos técnicos, financieros, económicos y ambientales. Identificar el modelo de gestión más pertinente para el desarrollo del proyecto mediante el uso de estrategias consideradas por la propia comunidad, que permitan garantizar la sostenibilidad en el tiempo de este emprendimiento productivo. 202 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Metodología para los estudios: técnico, de mercado, financiero y evaluación económica-financiera y ambiental. Para el estudio de viabilidad técnica, productiva, comercial y ambiental del jabón artesanal “YAKUMAMA”, se seguirá el procedimiento metodológico propuesto en la obra Preparación y Evaluación de Proyectos de Inversión (Andrade, P. 2012). Estudio técnico del proyecto El estudio técnico del proyecto comprende el análisis del tamaño o capacidad de producción, identifica la localización óptima, define la ingeniería del proyecto y finalmente el proceso productivo. Tamaño o capacidad de producción Se calculará la producción óptima de acuerdo a la capacidad instalada existente en la comunidad. Localización Para la localización se procederá identificando al nivel de: Macro localización y Micro localización. Ingeniería del proyecto Condiciones para la producción del jabón artesanal Se realizará un análisis de infraestructura (plano de la fábrica de jabón), en donde se detallarán espacios de uso para elaboración de jabón: área de producción, área de maduración, área de almacenamiento, área de limpieza, área de empaquetado, bodega, área administrativa, área de ventas. Descripción técnica del producto Para establecer un producto de calidad se debe tener muy en cuenta los siguientes aspectos: Presentación, Unidad de Medida, Formas de Almacenamiento, Insumos, Mano de Obra, Transporte, Vida Útil y perecibilidad, Maquinaria. Proceso productivo.-Se realizará por medio del flujograma de producción, a través del diseño de la cadena productiva (jabón artesanal “YAKUMAMA”). Estudio de mercado El estudio de mercado pretende determinar la demanda potencial del jabón artesanal “YAKUMAMA”, en el mercado hotelero del cantón Tena, para lo cual es preciso analizar el precio y las técnicas de comercialización que permitan introducir con éxito dicho producto. Tamaño del Universo Se tomará en cuenta el número de establecimientos con capacidad de alojamiento, existentes en el cantón Tena, según el Catastro de los Establecimientos Turísticos 2013 facilitado por la Unidad de Turismo y Biodiversidad del Gobierno Autónomo Descentralizado Municipal de Tena. 203 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Cálculo de la Demanda Real Análisis de la Demanda La demanda del producto será obtenida de los datos de la encuesta, que serán calculados anualmente, según el siguiente artificio. Proyección de la Demanda Para la estimación de la proyección demanda se considerará que el incremento de la demanda está relacionado con el crecimiento del mercado hotelero (número de hoteles y afluencia de turistas) del cantón Tena. Análisis de la Oferta Para la oferta de los jabones, en el Cantón Tena, existen empresas que producen jabones para el mercado hotelero, los mismos que no son artesanales y son de una sola esencia, y no presentan mayor variedad. Proyección de la Oferta Para realizar la proyección de la oferta del jabón artesanal se utilizará el porcentaje de crecimiento de la actividad económica. Demanda Real Debemos establecer la estimación de la demanda real de mercado en el momento actual, así como también realizar las estimaciones de demanda potencial para nuestro producto. La demanda potencial será la cantidad sobre la que se trabajará, dado por la producción de jabón artesanal “YAKUMAMA”. Análisis de precio Análisis de Precios de la Competencia Se realizará el análisis de los precios de los jabones para el mercado hotelero que existen actualmente ya que son competencia para el producto del estudio. Proyección de los Precios al Público Los precios al público serán calculados según el costo de producción y el porcentaje de ganancia. Análisis de comercialización Para la comercialización del jabón artesanal “YAKUMAMA”, se plantearán las siguientes estrategias: Estrategia de promoción, Estrategia de presentación, Estrategia de distribución. 204 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Estudio financiero El estudio financiero determina los niveles de ingresos y egresos que permitirán verificar los resultados que genere el proyecto, al igual que la liquidez que tiene el emprendimiento para cumplir con sus obligaciones operacionales y no operacionales. Para este estudio, se elaborarán cuadros de costos, balances y flujos de caja que permitan evaluar los elementos del estudio financiero. Estimación de ingresos y costos Presupuesto de Costos.- El presupuesto de costos estará basado en los costos que están directamente ligados a la producción, así como los que se relacionan indirectamente. Presupuesto de Ingresos.-El presupuesto de ingresos corresponderá a un precio de venta de cada producto por unidad, así como al número de unidades vendidas por año estimadas en la proyección de mercado. Plan de inversiones El costo del proyecto.- Se elaborará cuadros del total de inversiones para el proyecto y capital de trabajo. Resultados y Situación Financiera Proyectados.- se trabajará con: Estado de pérdidas y ganancias, Balance general, Flujo de Caja Evaluación económico - financiera En la evaluación económico-financiera se calculará y analizará la Tasa Interna de Retorno (TIR), el Valor Actual Neto (VAN) e índices financieros del proyecto, relacionando aspectos sociales, económicos que contribuyan al desarrollo de la comunidad. Cálculo del Valor Actual Neto (VAN) El VAN es una de las herramientas más usuales para calcular la rentabilidad del proyecto. Cálculo de la Tasa Interna de Retorno (TIR) La TIR es aquella tasa de interés que iguala el valor actualizado del flujo de los ingresos con el valor actualizado del flujo de costos. La tasa interna de retorno (TIR) mide la rentabilidad del dinero que se mantendrá dentro del proyecto. Punto de equilibrio Se calculará el punto de equilibrio de acuerdo a los datos financieros obtenidos, para saber cuál es la cantidad mínima que se debe producir y vender para cubrir costos y gastos. Análisis de sensibilidad El análisis de sensibilidad servirá para medir cuán sensible es el proyecto frente a cambios en el mercado (positivo, negativo), como variación en: cantidad, precio, costos fijos, costos variables, etc. Estudio Ambiental 205 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Para realizar la evaluación ambiental es necesario conocer claramente el proceso productivo de la elaboración del jabón artesanal “YAKUMAMA”, para identificar si las fuentes de materia prima son amigables o perjudiciales para el medio ambiente y elaborar medidas que ayuden a minimizar o anular las posibles consecuencias ambientales. La evaluación ambiental es una herramienta que asegurará que los recursos invertidos darán al proyecto la sostenibilidad a largo plazo que es esencial. (FAO, 2005) Se realizará la recopilación de información de proyectos similares que sirvan como referencia de análisis de los principales impactos ambientales Se utilizará: libros, revistas, informes, documentos. Para el desarrollo del Estudio de impacto ambiental, se utilizará como herramienta de apoyo: Ficha de Evaluación Ambiental-Anexo 2 (CFN, 2002). Metodología para diseñar el Modelo de Gestión El modelo de gestión óptimo para la comunidad Río Blanco permitirá garantizar la sostenibilidad en el tiempo de este emprendimiento productivo, que mejore su calidad de vida. Se utilizará técnica documental, se buscará información de uno de los expertos en el tema, y que hayan manejado modelos de gestión en comunidades, mediante la preparación previa de una guía de entrevista. Para el análisis general de la comunidad Río Blanco se utilizará la herramienta: matriz FODA (Fortalezas, Oportunidades, Debilidades, Amenazas), para el éxito del proyecto. BIBLIOGRAFÍA Andrade, P. 2012. Preparación y Evaluación de Proyectos de Inversión. Quito, Ecuador. p. 6-67. Arévalo, V. 2009. Chakras, bosques y ríos. El entramado de la Biocultura Amazónica. Publicación miscelánea No 148, INIAP. Editorial Abya – Yala. Quito, Ecuador. p. 62. FAO (Organización de las Naciones Unidas para la Agricultura y la Alimentación) 2000. Productos Forestales No Maderables. Guía de apoyo para el desarrollo de actividades empresariales. Publicación FAO. Quito, Ecuador. p. 7-15. FAO (Organización de las Naciones Unidas para la Agricultura y la Alimentación). 2010. Consulta de expertos sobre productos forestales no madereros. Grijalva, J.; Arévalo V.; Wood Ch. 2004. Expansión y trayectorias de la ganadería en la Amazonía del Ecuador. Publicación miscelánea del INIAP. Editorial Tecnigrava. Quito, Ecuador. p. 201. INIAP (Instituto Nacional Autónomo de Investigaciones Agropecuarias). 2012. Proyecto Genforestal: Conservación y Uso Sostenible de Recursos Genéticos Forestales en áreas críticas de bosques húmedos y secos de los Andes y Amazonía. Publicación INIAP. Quito, Ecuador. p. 26. MAE (Ministerio de Ambiente). 2010. Aprovechamiento de recursos forestales en el Ecuador y proceso de fracciones y decomisos. Publicación OIMT. Quito, Ecuador. p. 3:14. Miranda, J. 2005. Gestión de Proyectos: evaluación financiera económica, social, ambiental. 5 ed. Bogotá, Colombia. MM editores. p.114. 206 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Ospina. J.; Aldana, H. 1995. Economía, Administración y Mercadeo Agropecuarios: Análisis Financiero. Santa Fe, Colombia. Terranova. p. 133. Unidad de Turismo y Biodiversidad del Gobierno Autónomo Descentralizado Municipal de Tena. 2013. Catastro de Establecimientos Turisticos. Tena, Ecuador. s.p. 207 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 VALIDACIÓN DE UN PROTOCOLO PARA LA PROPAGACIÓN MASIVA IN VITRO DE Paulownia elongata, Paulownia fortunei y un híbrido (Paulownia fortunei x Paulownia elongata) BAJO SISTEMAS DE PROPAGACIÓN CONVENCIONAL E INMERSIÓN TEMPORAL Cárdenas, A; Grijalva, J; Morillo, E; Ramos, R; Meneses, S Instituto Nacional Autónomo de Investigaciones Agropecuarias. Estación Experimental Santa Catalina. Programa Nacional de Forestería. Panamericana Sur Km1, sector Cutuglahua, Quito [email protected] RESUMEN Ecuador está experimentando una evolución en la explotación maderera, el promedio de ingresos de las exportaciones en el período 1995 a 2000 fue de US$ 100 millones, pero el problema radica en que las exportaciones no sobrepasan el 2,2 % y las importaciones el 4,2%, existiendo así un déficit que no cubre el mercado del sector maderero. Por este motivo la introducción de especies de rápido crecimiento como son las del género Paulownia spp. es indispensable para abastecer de productos forestales al mercado nacional y para la exportación, para así disminuir la presión sobre bosques naturales. Pero una de las limitantes, es la disponibilidad de material genético homogéneo; nace así la necesidad de implementar herramientas biotecnológicas como la propagación in vitro a fin de obtener plántulas iniciales libres de organismos patógenos, homogeneidad del material, mayor vigor de las plantas regeneradas, multiplicación durante todo el año, ahorro de espacio, fácil transporte e intercambio de material vegetal. Con esta investigación se evaluará bajo las condiciones de los laboratorios en Santa Catalina la propagación in vitro convencional mediante organogénesis directa, proponiéndose además evaluar la respuesta bajo biorreactores o Sistemas de Inmersión temporal que es una tecnología alternativa para la multiplicación acelerada de especies vegetales. Palabras clave: Paulownia, propagación in vitro, organogénesis directa, sistemas de inmersión, maderable. INTRODUCCIÓN Paulownia spp. es el único género con especies arbóreas de la familia Scrophulariaceae. Las nueve especies de este género son originarias de China excepto Paulownia fortunei y Paulownia tomentosa. Específicamente para proyectos forestales, las especies más utilizadas son Paulownia elongata, Paulownia fortunei y Paulownia kawakamii (Lucas et al., 2011). Este árbol empezó a ser desarrollado genéticamente, a comienzos de la década de 1990, para que se adaptara a distintos climas a fin de promover su cultivo en el mundo, tanto para reforestación como para uso maderable y energético. Este género se adapta a una gran variedad de climas pues el rango de temperaturas al que pueden adecuarse las especies varia ampliamente, llegando a soportar mínimas absolutas de -20º C y máximas absolutas de 45ºC. Algunas experiencias han demostrado que el rango óptimo de temperaturas para el crecimiento en altura y diámetro se localiza usualmente entre 24º C y 29ºC de temperatura media diaria. Pawlonia spp. se reproduce a través de semillas y esquejes de tallo y raíz. En cuanto a la primera forma, uno de los principales problemas es que pueden presentar dormancia, además de que el crecimiento de las plántulas es menor que las derivadas de esquejes o multiplicadas in vitro. Respecto a la segunda, los 208 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 esquejes de tallo, en general, son más difíciles de obtener; no obstante, en los de raíz el daño físico causado a la epidermis y al cortex provoca el ataque de patógenos. Dentro de las técnicas de cultivo in vitro utilizadas en el género Paulownia se han reportado la organogénesis directa e indirecta, la embriogénesis somática directa e indirecta, de éstas, la organogénesis directa usando yemas axilares es la técnica que comúnmente se emplea para la propagación masiva comercial, mientras que la embriogénesis somática se maneja más en protocolos de mejoramiento genético. Existe interés particular en conocer las características y bondades de estas especies de rápido crecimiento para su uso futuro en sistemas de producción agroforestal y en plantaciones forestales en el Ecuador, por lo que la validación y generación de tecnologías in vitro para la multiplicación de materiales será de gran utilidad en esta iniciativa del Gobierno Nacional. JUSTIFICACIÓN La introducción de especies de rápido crecimiento es indispensable para abastecer de productos forestales al mercado nacional y para la exportación para así disminuir la presión sobre bosques naturales. Debido a las características de Paulownia, la Presidencia de la República del Ecuador ha solicitado de manera prioritaria su introducción como alternativa a problemas forestales que tiene el Ecuador ya que se explotan especies nativas e introducidas para cubrir la demanda interna y externa de madera de artesanos, industrial o energía de biomasa, todo esto debido a que en los últimos años el Ecuador está experimentando una evolución en la explotación maderera; es así que según las estadísticas del MAE el promedio de ingresos de las exportaciones en el período 1995 a 2000 fue de alrededor de US$ 100 millones, pero el problema radica en que las exportaciones no sobrepasan el 2,2 % y las importaciones el 4,2% existiendo así un déficit que no cubre el mercado del sector maderero. Una de las limitantes para el éxito de la introducción de esta especie al Ecuador, es la disponibilidad de material genético homogéneo, libre de enfermedades, debido a que el uso convencional de propagación de dichas especies es por semillas, y estacas. En este ámbito la aplicación de biotecnologías como la propagación in vitro permitirá la obtención de plántulas libres de organismos patógenos, homogeneidad del material, mayor vigor de las plantas regeneradas, multiplicación durante todo el año, ahorro de espacio, y fácil transporte e intercambio de material vegetal. La biotecnología permite al sector agrícola y forestal participar en el éxito del sistema productivo aplicando técnicas que consisten en el aislamiento de partes específicas de una planta los cuales son insertados en un medio de cultivo con los nutrimentos hormonales bajo condiciones de asepsia. OBJETIVO Validar varios protocolos de propagación in vitro realizadas para las especies pertenecientes al género Paulownia spp. para establecer la mejor metodología y poder implementar en programas de producción de plantas para la reforestación. METODOLOGÍA El estudio se realizará en cuatro fases: Fase I: Introducción in vitro de Paulownia elongata, Paulownia fortunei y un híbrido (Paulownia fortunei x Paulownia elongata). 209 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Se recolectará explantes de ramas jóvenes con nudos a partir de plantas madre que se encontrarán en el invernadero. Los explantes recolectados se colocarán inmediatamente en agua, para su posterior lavado con jabón líquido y abundante agua para eliminar posibles contaminantes. Luego se someterán a una desinfección con alcohol al 70%, hipoclorito de sodio y fungicidas para proceder a su introducción a diferentes medios de cultivo. Entre las variables que se evaluarán consta: Porcentaje de brotación Esta variable se evaluará con observaciones a los 10, 20, 30 días y se determinará el crecimiento de los brotes a partir de la siembra. Porcentaje de contaminación Se determinará el porcentaje de contaminación con observaciones frecuentes de las muestras a partir de la siembra hasta los 15 días después de la siembra. Se registrará los explantes contaminados de acuerdo a la fórmula: Porcentaje de oxidación Se contabilizará los explantes que presentaron oxidación a partir del primer día de siembra. Tasa total de pérdida Se contabilizará el número de explantes contaminados, número de explantes muertos que presento a los 15 días a partir de la siembra se utilizará la siguiente formula: Fase II: Multiplicación in vitro de Paulownia elongata, Paulownia fortunei y un híbrido (Paulownia fortunei x Paulownia elongata) bajo el sistema convencional e inmersión temporal. Se prepararán diferentes medios de cultivo con reguladores de crecimiento a diferentes concentraciones entre ellas BAP y ANA, y se probarán bajo los dos sistemas de propagación masiva. Entre las variables que se evaluarán para el sistema convencional serán: Altura de los brotes La evaluación se realizará a los 10, 20 y 30 días a partir de la siembra, se registrará los datos en mm con la ayuda de una regla milimetrada. Número de nudos Se contabilizará el número de nudos que presentará cada uno de los explantes a partir de la siembra a los 10, 20 y 30 días. Índice de multiplicación 210 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 El índice de multiplicación se evaluará a los 10, 20 y 30 días, posteriores a la siembra, se obtendrá contando el número de nudos obtenidos cada mes dividido para el número de plantas, para lo cual se utilizará la siguiente fórmula: Tasa de pérdida Se contabilizará el número de explantes contaminados, número de explantes muertos a partir de la siembra hasta los 10, 20 y 30 días para lo cual se utilizará la siguiente formula: Entre las variables que se evaluarán para los sistemas de inmersión temporal serán: Por observación visual, se registrará la brotación de los explantes a los 15 y 30 días a partir de la siembra. Tamaño de brotes Se determinará la longitud en centímetros de los brotes y se obtendrá un promedio de la longitud de brotes por cada tratamiento a los 30 días a partir de la siembra. Porcentaje de brotes hiperhidratados Se establecerá mediante relación porcentual entre el número brotes con hiperhidratación y el número total de brotes. Se evaluará a los 30 días. Índice de multiplicación Se registrará el número de explantes que produce una planta a los 30 días a partir de la siembra. Índice de multiplicación = (nº explantes * nº nudos) / nº de explantes iniciales Fase III: Enraizamiento in vitro de Paulownia elongata, Paulownia fortunei y un híbrido (Paulownia fortunei x Paulownia elongata), en medio sólido de explantes provenientes de la segunda fase. Se prepararán diferentes medios de cultivo con reguladores de crecimiento a diferentes concentraciones entre ellas IBA y ANA. Entre las variables que se evaluarán para el sistema convencional serán: Número de plántulas con raíces Se contabilizará el número de plántulas que desarrollen raíces a los 15 y 30 días de haber sido sembrado el brote en medio enraizante 211 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Número de raíces Se contará el número de raíces de cada brote, a los 15 y 30 días de haber sido sembrados. Longitud de las raíces Se determinará la longitud de la raíz principal a los 15 y 30 días haber sido sembrado el brote en medio enraizante. Se utilizará una regla milimetrada, y la medida se expresará en milímetros (mm). Fase IV: Adaptación a condiciones de invernadero de Paulownia elongata, Paulownia fortunei y un híbrido (Paulownia fortunei x Paulownia elongata) obtenidas in vitro. Se colocarán las plántulas en diferentes sustratos como son la turba y la pomina, y se evaluarán por observación visual, se registrará la sobrevivencia de las plantas a los 10, 20 y 30 días a partir de la siembra en invernadero. Se calificará de acuerdo a la siguiente escala, donde, 1 corresponderá a las plantas vivas y 0 a las plantas muertas. RESULTADOS ESPERADOS Se espera obtener un porcentaje mínimo del 86% de brotación aplicando los tratamientos de cada protocolo que se evaluará en cada etapa de experimentación, bajo el sistema convencional e inmersión temporal empleando como tejido ideal los segmentos internodales según la bibliografía consultada. BIBLIOGRAFÍA Aguilar, M., Villalobos, A., Salgado, R. 2001. Cultivo in vitro de Paulonia (Paulownia tomentosa). Revista Mexicana de ciencias forestales. 19 p. Consultado 5 de sep.2013. Disponible en: http://agroforestal.com.mx/content/cultivo-vitro-de-paulonia Bergmann, B., and Moon, H. 1997. In vitro adventitious shoot production in Paulownia. Plant Cell Rep. 16(5):315-318. Consultado13sep. 2013. Disponible en: http://link.springer.com/article/10.1007%2FBF01088288#page-1 Castellanos, O., Rodríguez, A., Rodríguez, J., Rodríguez, B. 2006. Organogénesis indirecta y enraizamiento in vitro de Paulownia elongata. Red de Revistas Científicas de América Latina, el Caribe, España y Portugal. 4(15):13. Consultado 9 sep. 2013. Disponible en: http://www.redalyc.org/pdf/730/73000415.pdf. Corredoira, E., Cernadas, J., San José M. 2010. Inducción de yemas adventicias en Paulownia tomentosa estudio anatómico del proceso caulogénico. Santiago de Compostela, España. Revista Real Academia Galega de Ciencias 29: 5-22. Consultado 15 sep.2013. Disponible en: http://www.ragc.cesga.es/RRAGC /revista2010/pdf/Inducciondeyemas.pdf. Grijalva, J., X. Checa, R. Ramos, P. Barrera y R. Limongi. 2012. Situación de los Recursos Genéticos Forestales – Informe País Ecuador. Preparado por el Programa Nacional de Forestería del INIAP con aval del INIAP/FAO/MAE/MAGAP/MMRREE. Documento sometido a la Comisión Forestal de la FAO-Roma, para preparación del Primer Informe sobre el Estado de los Recursos Genéticos Forestales en el Mundo. 95 p. Ipecki, Z., Gozukirmizi, N. 2004. Indirect somatic embryogenesis and plant regeneration from leaf and internodal explants of Paulownia elongata. Plant Cell Tissue and Organ Culture. 79(3):341-345. 212 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 MAE. 2011. Estimación de la Tasa de Deforestación del Ecuador continental. Disponible en:http://www.ambiente.gob.ec/sites/default/files/users/mponce/TasasDeforestacionEcuador.Ver_.03.05.1 1.pdf. Zhao-Hua, Z., X. YaoGuo y L. Xin-Yu, 1986. Paulownia in China; cultivation and utilization.The Chinese Academy of Forestry. Asian Development for Biological Science and International Development Research Centre, Beiging, China. 213 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 ZONIFICACIÓN AGROECOLÓGICA DEL CULTIVO DE CAUCHO Hevea brasiliensis EN EL ECUADOR EN BASE A LA INCIDENCIA DEL MAL SURAMERICANO DE LAS HOJAS Lucrecia Maldonado, Víctor Cevallos, Franck Rivano, Blanca Simbaña, Trajano Yugcha , Rigoberto Lucero INIAP, Estación Experimental Santo Domingo, Km 38 Vía Santo Domingo-Quinindé. Apartado postal:17-24-101 [email protected] INTRODUCCIÓN El caucho Hevea brasiliensis es originario de la Amazonía. El primer registro científico del látex y su uso fue descrito por el científico francés La Condamine durante la Misión Geodésica en Ecuador hace más de 250 años. A pesar de ser nativo de la Amazonía, presenta un mejor comportamiento fuera de su lugar de origen ante el ataque de la Enfermedad Suramericana de las Hojas, principal problema fitosanitario que afecta al cultivo, causada por el hongo Microcyclus ulei, responsable de provocar sucesivas defoliaciones en clones susceptibles, hasta producir la muerte. En la actualidad los cultivos comerciales se extienden entre los 22°N y 25°S lo cual demuestra su capacidad de adaptación a una diversidad de condiciones edafoclimáticas. El origen de las plantaciones en el Ecuador se remonta al año 1966 en una propiedad de la provincia de Pichincha, con una superficie de 25 ha, plantación que la podríamos calificar como una prueba de adaptación del cultivo a las condiciones agroclimáticas de nuestro país. Comercialmente se difundió a partir de 1970; el “boom” de su crecimiento ocurre a finales de los años ochenta donde se incrementó la demanda por parte de la pequeña industria, lo que despertó el interés en los agricultores, quienes comenzaron a sembrar superficies considerables con fondos propios, lo que permitió alcanzar en 1993 una superficie total de 4 068 ha (Malo et al., 1993), y que en el 2003 alcanzó 9 246 ha, de las cuales aproximadamente el 37% habrían sido eliminadas por su avanzada edad y por problemas patológicos (Rivano, 2004). Desde el fenómeno de El Niño entre 1997-1998 las plantaciones han atravesado limitaciones de producción y expansión, principalmente debido a la Enfermedad Suramericana de las Hojas, donde es ineludible la siembra de materiales con resistencia genética al patógeno y de alta productividad. De acuerdo al último censo cauchero existen 4 942 ha de cultivo distribuidas en diferentes provincias del país: Los Ríos con un 40% de la plantación nacional, Santo Domingo de Los Tsáchilas con el 32%, Esmeraldas con el 15%, Pichincha con el 11%, Cotopaxi 1% y Guayas 1% (Torres, 2009). El objetivo del presente estudio fue determinar las zonas con características edafo-climáticas favorables para el cultivo del caucho, con respecto a la incidencia del Mal Suramericano de las Hojas. MATERIALES Y MÉTODOS Para realizar la zonificación agroecológica del cultivo, se utilizó como herramienta los Sistemas de Información Geográfica SIG con el software ArcGIS 9.3 y sus diferentes aplicaciones ArcMap, ArcCatalog y ArcToolbox. Para la generación del mapa de zonas aptas se consideraron las exigencias climáticas del cultivo de caucho y del hongo Microcylus ulei; para lo cual se adoptaron los diferentes parámetros: geomorfológicos 214 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 (pendiente), edáficos (textura del suelo, pedregosidad, pH, nivel freático, profundidad), climáticos (precipitación, temperatura) y altitud. La información climática corresponde a un historial desde 1980 a 2005, con una serie de veinticinco años, que fue recopilada de estaciones meteorológicas pertenecientes al Instituto Nacional de Meteorología, obtenidos del Sistema de Información Nacional de Agricultura, Ganadería, Acuacultura Y Pesca – SINAGAP (ex-SIGAGRO) – Dirección de Investigación y Generación de Datos Multisectoriales, mientras que la cartografía de Áreas Naturales se adquirió del Ministerio del Ambiente. Todas las variables consideradas en la investigación (pendiente, suelos, precipitación, temperatura, altitud, etc.) corresponden a capas temáticas que se relacionaron entre sí hasta concluir en un mapa de zonas aptas, mediante la utilización de herramientas de geoprocesamiento y Analysis Tools de la aplicación ArcToolbox. Con los requerimientos climáticos del cultivo y también para la ocurrencia de la Enfermedad Suramericana de las hojas en condiciones naturales, se seleccionaron las zonas que cumplieron con esas exigencias, éste proceso con las herramientas Select Layer By Atribute y la expresión SQL (Query Builder) de la aplicación ArcToolbox. En la elaboración del mapa se excluyeron las zonas naturales empleando la herramienta Analysis Tools, ya que son consideradas como zonas de protección y conservación, obteniéndose así el Mapa de Zonificación Agroecológica para el Cultivo de Caucho en Ecuador. Los diferentes requerimientos agroecológicos del cultivo (Cevallos et al., 2012; Ministerio de Agricultura & CONIF, 1997; Ortolani et al., 1983; Pilau et al., 2007) se presentan en la Tabla 1. Tabla 1. Requerimientos Agroecológicos del Cultivo de Caucho Requerimientos Rango Pendiente Menor a 25 % Arenoso franco, Franco arenoso, Franco limoso, Franco, Franco arcillo, Franco Textura arcillo arenoso, Franco arcillo limoso Pedregosidad Menor a 25 % pH 4,5 a 6,5 Nivel Freático Mayor a 100 cm Profundidad Mayor a 50 cm Precipitación 1.250 mm a 3.500 mm Temperatura Mayor a 22° C Altitud Menor a 1.000 m.s.n.m. Para generar el mapa de Zonas de Escape, se elaboró el modelo cartográfico considerando las condiciones climáticas que no favorecen el desarrollo del hongo Microcyclus ulei, luego se procedió a realizar la superposición de las variables: número de meses con precipitación ≤ 50 mm, número de meses con precipitación ≤ 30 mm, precipitación y déficit hídrico, con los que se ubicó las zonas que cumplan con esos requerimientos. Este proceso se realizó con ayuda de la herramienta Select Layer By Atribute y la expresión SQL (Query Builder) de la aplicación ArcToolbox, obteniéndose de esta manera el Mapa. Los diferentes parámetros climáticos para la heveicultura las zonas de escape (Cevallos et al., 2012; Ortolani et al. 1983; Pilau et al. 2007) se presentan en la Tabla 2. 215 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Tabla 2. Características climáticas para la heveicultura bajo condiciones de escape a Microcyclus ulei Característica Numero de meses con precipitación ≤ 50 mm Numero de meses con precipitación ≤ 30 mm Precipitación Déficit Hídrico Rango 3-6 3-6 mayor a 1.250 mm 100 mm a 600 mm RESULTADOS Y DISCUSIÓN La Figura 1 representa el Mapa de Zonas Aptas del cultivo de caucho bajo condiciones naturales, considerando las exigencias climáticas del caucho y del hongo Microcyclus ulei. Comparando este mapa con el trabajo realizado por otros autores se observa claramente un aumento en la precisión y el detalle, conferidos por la mayor escala grafica utilizada y el mayor número de variables empleados en este estudio. Las zonas aptas para este cultivo alcanzan 749 824 ha (sin considerar áreas naturales). Entre en las zonas geográficas de impacto se encuentran las Provincias de Esmeraldas, Pichincha, Santo Domingo, Los Ríos, Guayas, Cotopaxi, Orellana y Sucumbíos con las cabeceras cantonales Muisne, Rosa Zárate, La Concordia, Puerto Quito, Pedro Vicente Maldonado, El Carmen, Santo Domingo, Yaguachi Nuevo, Milagro, parte de Colimes, Santa Lucía, Balzar, Vinces, El Salitre, Baquerizo Moreno, Santa Ana de Vuelta Larga, Olmedo, Shushufindi, Puerto Francisco de Orellana, Joya de los Sachas, Loreto, Nueva Loja, Putumayo. Figura 1. Mapa de zonas aptas para el cultivo de caucho en el Ecuador. En este mapa se puede distinguir zonas localizadas en la Región Litoral Norte (La Concordia, Santo Domingo, Quinindé, entre otras) que presentan condiciones muy favorables para la infección de las hojas por el hongo Microcyclus ulei, con una temperatura mínima promedio de 21°C, humedad relativa promedio anual de 88%, precipitación anual promedio de 3000 mm. Conforme se acerca a la zona central 216 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 del litoral se diferencia una zona con condiciones intermedias de presión de la enfermedad (Patricia Pilar, Los Ángeles, etc.) con las siguientes características: precipitación media anual de 2500 mm, temperatura mínima promedio de 21°C, humedad relativa promedio anual 85%, lo que favorece la esporulación de Microcyclus ulei. La Figura 2a representa la zona apta (color verde) y la zona de escape (color naranja) que comprende 1 242 641 ha., que incluye las áreas con alta restricción para la siembra de caucho bajo condiciones naturales; es así como sólo se considera verdaderamente zona de escape apta para el cultivo de caucho al área donde se intersectan los dos colores verde y naranja; de lo cual resultó la Figura 2b, que representa el Mapa de Zonas de Escape, con un área de 80 174 ha. Las zonas geográficas están localizadas en las provincias de Los Ríos, Guayas y una pequeña parte en Manabí, con las cabeceras cantonales Velasco Ibarra, Pichincha, Quevedo, Quinsaloma, Las Naves, Echandía, Pueblo Viejo, Catarama, Cumandá, El Triunfo, La Troncal, Simón Bolívar, que presentan las siguientes características climáticas: humedad relativa promedio de los dos meses más secos del año entre 77 y 78%, precipitación anual promedio de alrededor de 1900 mm, con un período seco de cinco meses del año, precipitación media inferior a 50mm, durante el cual ocurre la defoliación-refoliación de los árboles, condiciones que interrumpen la esporulación del hongo Microcyclus ulei, evitando que se produzca una infección por el bajo potencial de inóculo, lo que identifica la zona de escape. Figura 2. Mapa de Zonificación Agroecológica del cultivo de caucho en el Ecuador bajo condiciones naturales (a) Mapa de zonas aptas y de escape para el cultivo de caucho en el Ecuador, (b) Mapa de zonas de escape al Microcyclus ulei en el Ecuador. CONCLUSIÓN El Mapa de Zonificación Agroecológica del cultivo de caucho confirma la existencia de áreas con potencial para su expansión, lo que avizora una verdadera oportunidad para el desarrollo del cultivo en condiciones edafo-climáticas aceptables; sea zona de incidencia o de escape a la Enfermedad Suramericana de las hojas. BIBLIOGRAFÍA 217 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Cevallos, V. Maldonado L. Rivano, F. 2012. Estudio del comportamiento de tres clones comerciales de caucho Hevea brasiliensis en tres zonas caucheras del Litoral Ecuatoriano. Boletin Divulgativo N° 414. INIAP. La Concordia, EC. Ministerio de Agricultura & Corporación Nacional de Investigación y Fomento Forestal CONIF. 1997. Zonificación de áreas aptas para el cultivo de caucho, Santafé de Bogota, CO. Malo, F., Remache, M. & Sigua, J. 1993. El cultivo del caucho en el Ecuador. Agricultura Comercial S. A. Boletín Divulgativo Nº 3. 89 p, Quito, EC. Ortolani, A. A., Pedro Jr., M. J., Alfonsi, R. R., Camargo, N. B. D., Brunini, O .1983. Aptidão o agroclimática para regionalização da heveicultura no Brazil. In: Anais do Seminario Brasileiro para recomendação de clones de seringueira, Brasilia, BR, p. 19-28. Pilau F. G., Marin, F. R., Paes de Camargo M. B., Delgado Assad E., Silveira Pinto, H., Barbarisi, B. F. 2007. Zoneamento agroclimático da heveicultura para as regiões Sudeste e Centro-Oeste do Brasil. Revista Brasileira de Agrometeorologia 15 (2): 161-168 Rivano, F. Informe de Misión en el Ecuador. CIRAD-ASONHEV (Del 19 al 27 de enero 2004). Torres, D. 2009. Diagnóstico de la población, producción y rendimiento del caucho (Hevea brasiliensis). Tesis de Ingeniería Agropecuaria. Universidad Técnica de Quevedo. Quevedo, EC. 60 pp. 218 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 ANÁLISIS DE LOS PATRONES DE CRECIMIENTO RADIAL DE MASAS DE Pinus pinaster Ait., EN RESPUESTA A LA VARIABILIDAD CLLIMÁTICA EN EL CENTRO-SUR DE ESPAÑA Riofrío, J1. Bravo-Oviedo, A1,2. Bravo, F.1 1Instituto Universitario de Investigación en Gestión Forestal Sostenible, Universidad de Valladolid-INIA, Avda. Madrid 44, 34071, Palencia, España. 2Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria. Centro de Investigación Forestal (INIA-CIFOR). [email protected] Resumen La región Mediterránea se define como una zona de transición climática con alta sensibilidad ante los efectos del cambio climático. Las distintas respuestas que dicho cambio provoca sobre el crecimiento de las masas forestales se pueden evidenciar al estudiar la variación de las relaciones entre el crecimiento radial y el cambio de los factores climáticos durante los últimos años. Con el objetivo de evaluar si la relación clima-crecimiento es dependiente de la edad y determinar si existe un efecto significativo en los patrones de crecimiento inicial, se realizó el análisis dendroclimático de masas procedentes de repoblaciones de Pinus pinaster Ait., jóvenes (18 años) y adultos (53 años), ubicadas en el centro-sur de España. Se tomaron barrenas en las masas adultas y rodajas en las masas jóvenes de 10 individuos. Ambos grupos de edad tuvieron una respuesta diferenciada frente a las condiciones climáticas y mostraron un incremento de la variabilidad del crecimiento radial a partir de 1995. La correlación y función de respuesta de las variables climáticas estacionales sobre el crecimiento mostró una dependencia de la edad y del período de análisis, lo que indica una relación cambiante con el clima a través del tiempo. Los árboles adultos fueron más sensibles a la influencia climática que los árboles jóvenes. Finalmente, el análisis de respuestas mediante ventanas móviles permitió detectar un incremento en la sensibilidad de los árboles adultos a los eventos de sequía previos y en el inicio de la estación de crecimiento. Palabras clave: dendrocronología, variabilidad climática, patrones de crecimiento, análisis temporal, P. pinaster. Introducción Entre las principales ventajas del uso de anillos de crecimiento como indicadores del cambio o variabilidad climática están su capacidad para fechar con precisión las perturbaciones, la existencia de patrones de variabilidad interanual sincronizados a escala espacial y temporal, una la creciente red de cronologías en diferentes ecorregiones, y la posibilidad de utilizar modelos lineales simples para relacionar el clima con el crecimiento (Hughes 2002). Sin embargo, aún persisten algunas incertidumbres relacionadas con los complejos mecanismos fisiológicos mediante los cuales los parámetros climáticos se traducen en variaciones del crecimiento (Fritts 1976), además la asociación estadística entre el clima y el crecimiento puede cambiar en el tiempo (D’Arrigo et al. 2008). A pesar que se conoce que durante el crecimiento algunas características morfológicas y fisiológicas en los árboles difieren entre los estados juveniles y adultos (Bond 2000), aún no se conoce con certeza cómo los cambios en los procesos fisiológicos asociados al envejecimiento pueden alterar estas relaciones. Las investigaciones que han probado si la respuesta crecimiento-clima es estable a través de 219 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 diferentes grupos de edad, no son concluyentes debido a la amplia variedad de los resultados reportados (Carrer and Urbinati 2004, Rozas et al. 2009). El calentamiento climático está influyendo negativamente en el crecimiento y el vigor de los pinos mediterráneos, desencadenando episodios de decaimiento (Andreu et al. 2007). Los bosques de coníferas de montaña en el centro-sur de España podrían ser algunos de los más vulnerables a este efecto inducido por el cambio climático (Linares et al. 2009). El pino negral (Pinus pinaster Ait.) es una especie característica de los ambientes mediterráneos, donde forma masas monoespecíficas y mixtas en pinares tanto naturales como en repoblaciones, y a pesar que se ve afectada por eventos de sequía es mucho más resistente que otras especies de pinos distribuidos en la región, lo que la convierten en especialmente valiosa para la producción forestal (Sánchez-Salguero et al. 2012). El entendimiento de cómo la variabilidad climática de las últimas décadas ha afectado los patrones de crecimiento de los bosques mediterráneos es relevante para evaluaciones de impacto y proyecciones a largo plazo (Sabaté et al. 2002). En este sentido conocer si conforme se incrementa la edad de los individuos los cambios estructurales y funcionales relacionados con la ontogenia modifican las respuestas clima - crecimiento puede ser determinante. Para este estudio se establecieron dos hipótesis de trabajo; i) la sensibilidad y respuesta de los patrones crecimiento de P. pinaster a las variables climáticas son dependientes de la edad, y ii) existe un cambio significativo en los patrones de crecimiento inicial de P. pinaster debido a cambios en las condiciones climáticas. Materiales y métodos Sitio de estudio El sitio de estudio corresponde a los montes de Fuencaliente, ubicados en la Sierra Morena de la provincia de Ciudad Real, en el centro–sur de España. Las condiciones climáticas son típicas del clima Mediterráneo con veranos secos y precipitación máxima entre otoño e invierno, la temperatura media anual es de 15.2ºC y la precipitación media anual de 480mm (Fig. 1). Figura. 1. Ubicación del sitio de estudio sobre la distribución P. pinaster. Se seleccionaron dos masas de repoblaciones de Pinus pinaster Ait., localizadas a 1.76 km de distancia, de edades diferentes, adultos de 50 años y jóvenes de 20 años aproximadamente. Diez árboles de cada masa fueron muestreados durante mayo del 2012, se tomaron testigos o canutillos pareados con la barrena Pressler en las masas adultas y se cortaron rodajas de los árboles jóvenes. Construcción de cronologías 220 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Tanto canutillos y rodajas se secaron al aire y se lijaron progresivamente, con el uso de un escáner Epson Expresion 1640XL se obtuvieron imágenes digitales a una resolución 1100pdi y precisión de 0,01mm, el ancho de los anillos se midió con el programa WinDENDRO (REGENT INSTRUMENT INC., 2002). Las muestras fueron datadas siguiendo el procedimiento de datación cruzada estándar. La datación cruzada fue validada con el programa COFECHA V6.06P, que calcula el grado de sincronía o correlación de cada series individual con la cronología maestra (Speer 2010). El crecimiento radial de los árboles jóvenes tiende a ser más rápido que el de los árboles maduros, por lo que las series deben ser estandarizadas. La estandarización consiste en ajustar una función matemática que recoja dicha tendencia, la serie residual obtenida es un índice adimensional o índice de crecimiento, cuya variabilidad se atribuye principalmente al clima (Fritts 1976). Las series de árboles jóvenes y adultos se estandarizaron con una función spline suavizada, la rigidez de la función se ajustó de acuerdo a la longitud de las cronologías. Para la estandarización de las series se utilizó el programa estadístico R v.2.15.2 con la librería dplR para aplicaciones dendrocronológicas. Se aplicó un modelo autoregresivo a cada serie para descartar la autocorrelación relacionada con el crecimiento del año previo. Finalmente las cronologías medias de cada grupo de edad se obtuvieron del promedio de los índices de crecimiento usando una media robusta doblemente ponderada (Speer 2010). La calidad de las cronologías del ancho de anillos se evaluó con la media del grosor de los anillos, la autocorrelación (AC) y (r) la correlación media entre todas las series con la cronología maestra. Con los índices de crecimiento se calcularon: la sensibilidad media (MS) como medida de la variabilidad interanual, (rbt) la similitud en el crecimiento entre árboles y (EPS) la señal expresada por la población que es una medida de la calidad estadística de la cronología (Cook and Pederson 2011). Datos climáticos Las variables climáticas usadas fueron extraídas de la base de datos Spain02 (Herrera et al. 2012) obtenida de la interpolación de los datos diarios de precipitación y temperatura de Agencia Estatal de Meteorología (AEMET) para el período 1950-2007. Se calcularon los valores estacionales de precipitación y temperatura de invierno (W, Diciembre, Enero y Febrero), primavera (SP, Marzo, Abril y Mayo), verano (SM, Junio, Julio y Agosto) y otoño (AU, Septiembre, Octubre y Noviembre). Adicionalmente se calculó el índice de aridez (DRI) (Thornthwaite 1948). Relaciones entre variables climáticas y crecimiento Para determinar las variables climáticas que controlan el crecimiento, se utilizó la función de correlación y función de respuesta, que consisten en una secuencia de coeficientes calculados entre los índices de crecimiento y las variables climáticas (Biondi and Waikul 2004). Un método de remuestreo “bootstrapping” es comúnmente utilizado para obtener resultados precisos para las funciones de respuesta y correlación (Guiot 1991). El análisis de la relación entre clima y crecimiento se realizó con la librería bootRes disponible dentro de R (R Develomment Core Team 2010). Se consideró como período de análisis desde marzo (primavera) del año previo hasta noviembre (otoño) del año de crecimiento. Adicionalmente se evaluó la estabilidad de las relaciones entre crecimiento y clima en el tiempo, se obtuvieron los coeficientes en ventanas móviles de 20 y 10 años para los árboles adultos y jóvenes respectivamente. Resultados y discusión 221 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Descripción de cronologías En nuestros resultados las cronologías de ambos grupos de edad de P. pinaster, presentaron altos valores de MS (0,296 y 0,278 para árboles adultos y jóvenes) y EPS (>0,93 en ambos casos) (Tabla 1), lo que indica una respuesta fiable general de los patrones de crecimiento a factores ambientales (Cook and Pederson 2011), esto muestra que a pesar del corto período de análisis dendroclimático los individuos tuvieron la suficiente sensibilidad a las variables climáticas para manifestar respuestas en sus patrones de crecimiento. Patrones de crecimiento. En la cronología de ancho de anillo e índices de crecimiento de los árboles adultos (Fig. 2a, b) se observó un incremento de la variabilidad interanual a partir de las año 1995 en comparación con décadas pasadas, además los índices de crecimiento más extremos se presentaron en este período, patrones también observados por Bogino & Bravo (2009) y Vieira et al. (2010). Para el período común de crecimiento (1994 – 2011) ambas cronologías estuvieron débilmente correlacionados (r 0,219 – 0,277). Estos cambios son evidencias del efecto del cambio climático sobre el crecimiento de los bosques, tanto en la reducción del crecimiento como en el incremento de la variabilidad interanual. Tabla 1. Estadísticos descriptivos de las cronologías de ancho de anillos y cronologías residuales de masas adultas y jóvenes de P. pinaster. Adultos Intervalo Jóvenes 1959 - 2011 1994 - 2011 10 (20) 10 (19) Ancho medio de anillos (mm) 2,512 3,583 Sensibilidad media MS 0,295 0,277 Autocorrelación de primer orden 0,510 0,338 r 0,681 0,576 EPS 0,972 0,965 rbt 0,582 0,554 Nº árboles (cores) a ) b ) c ) 222 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figura. 2. a) Cronologías medias de ancho de anillos, b) Índices de crecimiento y c) Número de series en cada cronología de árboles adultos (línea gris) y árboles jóvenes (línea negra) de P. pinaster. Relación entre clima y crecimiento Las correlaciones obtenidas con el índice de aridez de los árboles jóvenes presentaron una correlación significativamente positiva con el DRI de otoño del año previo, mientras que correlaciones dispares se observaron en los árboles adultos en los dos períodos analizados (Fig 3). Los coeficientes significativos de la función de respuesta para DRI se obtuvieron en invierno y primavera del año de crecimiento en los árboles adultos entre 1994-2007, mientras para el período de crecimiento inicial los valores de DRI en verano del año de crecimiento presentaron una relación negativa, además en los árboles jóvenes no se observó ninguna relación significativa. En general las cronologías residuales de los árboles adultos fueron más sensibles a la influencia climática que la de los árboles jóvenes. Además la precipitación y el índice de aridez fueron las variables climáticas más determinantes y mostraron tendencias similares aunque con distintos niveles de significación. P. pinaster como otras especies en la región mediterránea es vulnerable a los períodos de sequía (SánchezSalguero et al. 2012), cuyos efectos dependen de la intensidad y recurrencia de los eventos. En nuestros resultados, los efectos de la sequía sobre el crecimiento también son dependientes de la edad, y al parecer a medida que aumenta la edad de los árboles los eventos de sequía previos y en el inicio de la estación de crecimiento se tornan más relevantes. Figura. 3. Coeficientes de correlación (a) y función de respuesta (b) entre cronologías residuales de árboles jóvenes y adultos con valores de DRI (barras blancas árboles jóvenes, barras gris claro árboles adultos 1959-1976, barras gris oscuro árboles adultos 1994-2007). Letras minúsculas indican el año previo y letras mayúsculas el año de crecimiento. Las relaciones significativas se representan con círculos blancos (p < 0,05). Esta mayor sensibilidad de árboles adultos se evidenció durante los períodos de sequía intensa (1999 y 2005), donde el índice de crecimiento de los árboles adultos fue menor que en los jóvenes, en cambio durante los años con alta disponibilidad hídrica su crecimiento se aceleró rápidamente (Fig. 2b), esto se puede interpretar como un comportamiento oportunista relacionado con la edad (Rozas et al. 2009) lo que indica mayor plasticidad de los árboles adultos, este comportamiento post-sequía ha sido observado en otras especies de pinos en España (Martínez-Vilalta et al. 2012). 223 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figura. 4. Correlaciones temporales del crecimiento al DRI calculadas en ventanas móviles. Se representan las series estacionales que fueron significativas. Líneas entrecortadas indican los límites a partir de los cuales las correlaciones son significativas. Análisis dinámico de las respuestas al clima Los cambios en la sensibilidad al clima durante los últimos años ha sido descrita por varios autores para P. pinaster y otras especies de pinos mediterráneos (Andreu et al. 2007). En nuestro caso, con ninguna de las tres variables climáticas analizadas se pudo simplificar una respuesta homogénea, ya que la asociación entre crecimiento y las variables climáticas cambió de acuerdo con al momento de análisis (Fig. 4), estos cambios se observaron en el nivel de significación y signo de las correlaciones. La relación con DRI reflejó un incremento general en la respuesta especialmente previo y en el inicio de la estación de crecimiento (W y SP) a medida que aumenta la edad, sin embargo también se apreciaron oscilaciones de las correlaciones durante todo el período analizado, esto indica que el clima podría haberse convertido en el factor más limitante para el crecimiento por el aumento del estrés hídrico y debido a la variabilidad de las precipitaciones que se ha registrado durante la segunda mitad del siglo XX (De Luis et al. 2009), ya que estos cambios aparecen vinculados a cambios en los patrones de crecimiento de los árboles. Conclusiones Las relaciones clima-crecimiento en P. pinaster son parcialmente controladas por la edad, ya que presentaron alta sensibilidad al período de análisis maximizando la señal climática en los árboles adultos. Estas relaciones fueron altamente sensibles a las fluctuaciones de las condiciones de disponibilidad hídrica previa y en el inicio de la estación de crecimiento, tornándose mucho más fuertes a medida que aumenta la edad de los árboles. El incremento de la sensibilidad media de los árboles adultos produce un mayor impacto y capacidad de reacción durante eventos de sequía y post-sequía respectivamente, esta sensibilidad podría aumentar si se incrementan la frecuencia e intensidad de las condiciones de sequía. Debido a que estos resultados se refieren a un área geográfica limitada y a un conjunto de datos relativamente pequeño, deben considerarse como un indicio para estudios más amplios que consideren diferentes regiones y especies, a fin de encontrar respuestas más generales que se incluyan en futuras evaluaciones de impacto de cambio climático sobre la dinámica de sistemas forestales. Bibliografía Andreu, L; Gutiérrez, E; Macias, M; Ribas, M; Bosch, O; Camarero, JJ; Macías, M. 2007. Climate increases regional tree-growth variability in Iberian pine forests. Global Change Biology 13. Biondi, F; Waikul, K. 2004. DENDROCLIM2002: A C++ program for statistical calibration of climate signals in tree-ring chronologies. Computers & Geosciences 30(3): 303–311. 224 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Bogino, S; Bravo, F. 2009. Climate and intraannual density fluctuations in Pinus pinaster subsp. mesogeensis in Spanish woodlands. Canadian Journal of Forest Research 39(8): 1557–1565. Bond, BJ. 2000. Age-related changes in photosynthesis of woody plants. Trends in plant science 5(8): 349–53. Carrer, M; Urbinati, C. 2004. Age-Dependent Tree-Ring Growth Responses To Climate in Larix Decidua and Pinus Cembra. Ecology 85(3): 730–740. Cook, ER; Pederson, N. 2011. Uncertainty, Emergence, and Statistics in Dendrochronology. Dendroclimatology. Dordrecht, Springer Netherlands, v.11, p.77–112. . D’Arrigo, R; Wilson, R; Liepert, B; Cherubini, P. 2008. On the “Divergence Problem” in Northern Forests: A review of the tree-ring evidence and possible causes. Global and Planetary Change 60(3-4): 289–305. Fritts, HC. 1976. Tree Rings and Climate. s.l., The Blackburn Press, p.567. . Guiot, J. 1991. The bootstrapped response function. Tree-ring bulletin 51: 39–41. Herrera, S; Gutiérrez, JM; Ancell, R; Pons, MR; Frías, MD; Fernández, J. 2012. Development and analysis of a 50-year high-resolution daily gridded precipitation dataset over Spain (Spain02). International Journal of Climatology 32(1): 74–85. Hughes, MK. 2002. Dendrochronology in climatology – the state of the art. Dendrochronologia 20(1-2): 95–116. Linares, JC; Camarero, JJ; Carreira, JA. 2009. Interacting effects of changes in climate and forest cover on mortality and growth of the southernmost European fir forests. Global Ecology and Biogeography 18(4): 485–497. De Luis, M; González-Hidalgo, JC; Longares, LA; Stepánek, P. 2009. Seasonal precipitation trends in the Mediterranean Iberian Peninsula in second half of 20th century. International Journal of Climatology 29(9): 1312–1323. Martínez-Vilalta, J; López, BC; Loepfe, L; Lloret, F. 2012. Stand- and tree-level determinants of the drought response of Scots pine radial growth. Oecologia 168(3): 877–88. R Develomment Core Team. 2010. R: A language and environment for statistical computing. R Foundation Statistical Computing . Rozas, V; DeSoto, L; Olano, JM. 2009. Sex-specific, age-dependent sensitivity of tree-ring growth to climate in the dioecious tree Juniperus thurifera. The New phytologist 182(3): 687–97. Sabaté, S; Gracia, C; Sánchez, A. 2002. Likely effects of Climate Change on growth of Quercus ilex, Pinus halepensis, Pinus pinaster, Pinus sylvestris and Fagus sylvatica forests in the Mediterranean Region. Forest Ecology and Management 162: 23–37. Sánchez-Salguero, R; Navarro-Cerrillo, RM; Camarero, JJ; Fernández-Cancio, Á. 2012. Selective drought-induced decline of pine species in southeastern Spain. Climatic Change 113(3-4): 767–785. Speer, JH. 2010. Fundamentals of Tree-Ring Research. Tucson, University of Arizona Press, p.333. . Thornthwaite, C. 1948. An approach toward a rational classification of climate. Geographical review 38(1):55–94 Vieira, J; Campelo, F; Nabais, C. 2010. Intra-annual density fluctuations of Pinus pinaster are a record of climatic changes in the western Mediterranean region. Canadian Journal of Forest Research 40(8): 1567– 1575. 225 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 226 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 POBLACIONES RURALES Y MEDIOS DE VIDA ASOCIADAS AL BOSQUE Doris Chalampuente SEGURIDAD ALIMENTARIA EN COMUNIDADES INDÍGENAS DE COSTA RICA: EL CASO DE COMUNIDADES CABÉCAR DE ALTO CHIRRIPÓ Galo Fernando Pillajo BIOINDICADORES: “SEGUROS LOCALES” PARA PEQUEÑOS PRODUCTORES AGROPECUARIOS 227 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 SEGURIDAD ALIMENTARIA EN COMUNIDADES INDÍGENAS DE COSTA RICA: EL CASO DE COMUNIDADES CABÉCAR DE ALTO CHIRRIPÓ M.Sc. Doris Chalampuente Centro Agronómico Tropical de Investigación y Enseñanza-CATIE [email protected] PROBLEMÁTICA Según la Organización para la Agricultura y Alimentación mencionan que los pueblos indígenas son creadores, proveedores y conservadores de la biodiversidad y su cultura, además de ser responsables de algunas prácticas más sostenibles desde el punto de vista ecológico; sus sistemas de subsistencia como el pastoreo, recolección, caza y pesca los ha hecho interdependientes de los recursos naturales y del medio ambiente, constituyen actividades que forman parte de sus medios de vida, pero que a la vez se ven afectados por la degradación ambiental, el mismo que se asocia a cambios climático lo que ha alterado la relación de los pueblos indígenas y el medio ambiente (FAO 2009). Las prácticas y conocimientos tradicionales de los pueblos indígenas son únicos, viables y validados a través del tiempo, el poder usar y conservar su biodiversidad permite tener alternativas de alimentación que contribuyen a la seguridad alimentaria y promueven su bienestar individual y colectivo. A través del enfoque de medios de vida, se ha llegado a conocer las actividades productivas y reproductivas que realizan los indígenas de tres comunidades de Alto Chirripó (Costa Rica), con lo cual llegan a satisfacer sus necesidades básicas. A través de la presente investigación se ha identificado los recursos y actividades de la comunidad, permitiendo conocer factores internos y externos así como aspectos de vulnerabilidad que afectan o contribuyen a la seguridad alimentaria local, siendo punto de partida para la creación de propuestas en base a las potencialidades e intereses de la comunidad. OBJETIVO GENERAL o Analizar los medios de vida en relación con la seguridad alimentaria de las comunidades indígenas Cabécar de Alto Chirripó, Costa Rica y formular propuestas para reducir su vulnerabilidad. Objetivos específicos o Caracterizar los medios y estrategias de vida que aportan a la seguridad alimentaria en comunidades indígenas Cabécar de Alto Chirripó. o Identificar las condiciones y factores de vulnerabilidad que podrían afectar la seguridad alimentaria en el corto y mediano plazo para las comunidades indígenas Cabécar de Alto Chirripó. o Proponer líneas de trabajo para reducir la vulnerabilidad alimentaria de las poblaciones estudiadas. 228 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 METODOLOGÍA Selección de las comunidades: se trabajó con tres comunidades pertenecientes a la Reserva Indígena de Chirripó (Alto Quetzal, Paso Marcos de Simiriñak y Simiriñak) para lo cual se tomó en consideración aspectos como: accesibilidad a la zona (tiempo y distancia), información previa, disponibilidad de actores locales claves como los Asistentes de Atención Primaria (ATAP) y profesores de la zona; en total se trabajó con 37 familias que corresponde al 27% del total registrado en la zona. Análisis de Medios y Estrategias de Vida: se aplicó una entrevista semi estructurada sobre medios de vida y necesidades fundamentales, se realizaron talleres con grupos focales, observación participante y diálogo con representantes de Instituciones, además se tomaron medidas antropométricas (talla y peso). Análisis de vulnerabilidad de seguridad alimentaria: basado en la información obtenida en entrevistas y encuestas de los medios de vida, se consideró las variables más representativas de cada capital y se construyeron índices siendo el valor más alto el de menor vulnerabilidad, mientras que los más bajos indican una alta vulnerabilidad (alto=1, medio=2 y bajo=3). Los resultados obtenidos fueron representados en un gráfico radial (telaraña). Análisis Estadístico: Para el análisis de resultados se empleó estadística descriptiva a través del uso de Info Stat-Statistical Software (Di Rienzo et ál. 2012) RESULTADOS Y DISCUSIÓN Descripción de los medios de vida y capitales o recursos presentes en las comunidades indígenas de Alto Chirripó En las comunidades Alto Quetzal, Paso Marcos de Simiriñak y Simiriñak se identificaron actividades o medios de vida que pueden ser productivos, extractivos y de reproducción. Los principales medios de vida son: a) actividades agropecuarias, b) venta de productos excedentes de la finca, c) extracción de productos del bosque, d) prestación de servicios como elaboración de alimentos, educadores, otros, e) venta de mano de obra, f) remesas o ayudas del estado. Las personas entrevistadas han diversificado sus actividades para lograr satisfacer sus necesidades fundamentales básicas como es el acceso a alimentación, salud, educación, entre otras. Los medios de vida reproductivos están relacionados con la disponibilidad de tierras (capital físico/financiero), acceso a educación y medicina (capital humano), pertenencia a una organización (capital social), actividades culturales y recreativas (capital cultural), maternidad e identidad ciudadana (capital humano y cultural). A través del análisis de componentes principales, se pudo identificar las combinaciones de medios de vida que se relacionan a cada comunidad (Figura 1). 229 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 1.21 Colecta café Eje 2 (31.69%) 0.75 Trab. Temporal Paso Marcos S. 0.28 Becas Suplementos alimenticios Pensión Alto Quetzal -0.18 Venta prod.agr MO carreteras Simiriñak -0.65 -0.76 -0.33 0.10 0.53 0.97 Eje 1 (68.31%) Comunidad Medios de vida Figura1. Relación entre Medios de Vida y Comunidades de Alto Chirrripó (Alto Quetzal, Paso Marcos de Simiriñak y Simiriñak). Las familias indígenas de las comunidades en mención, cuentan con estrategias de vida mixtas, donde se combinan las actividades agrícolas, pecuarias, remesas, comercio y demás; existen tres medios que son comunes dentro del grupo de entrevistados y están relacionados con la producción agrícola, pecuaria y extracción de leña, adicional a estos medios las familias pueden disponer de entre un medio de vida hasta once medios de vida. Las prácticas de recolección y de cultivo son parte de estrategias de manejo ambiental que promueve la sostenibilidad de los recursos y continuidad de las estrategias de vida de las comunidades (Montoya 2011); la pérdida y/o disminución de estos recursos ha ido transformando la agricultura de autoconsumo a una comercial con dependencia exclusiva de dos cultivos (frijol y banano), que a la vez resulta tener limitaciones por las condiciones físicas en las que se encuentran (comunidades económicamente aisladas) además de tener una escasa relación comercial entre ellos mismos o con terceros. Percepción de las Necesidades Fundamentales Las familias de las comunidades indígena de Alto Chirripó han logrado satisfacer algunas de sus necesidades humanas fundamentales en base a las actividades o medios de vida disponibles y en base a su percepción estas necesidades (básicas, de la persona, de entorno y de acción) fueron calificadas como: muy bueno o satisfechas (5), bueno o aceptablemente satisfechas (4), regular o medianamente satisfechas (3), malo o ligeramente satisfecha (2) y muy malo o insatisfechas (1). A nivel comunitario, existen diferencias en cuanto a necesidades básicas satisfechas que para unas resultan ser más satisfechas y para otras no, en este sentido para Paso Marcos de Simiriñak y Simiriñak, las necesidades de la persona (relación entre vecinos, familiares y el estado de felicidad) son 100% por ciento satisfechas. Un aspecto ligeramente satisfecho es la generación de ingresos, algunos lo atribuyen a la falta de acceso a mercados y otros a la falta de capacitación. Las necesidades de alimentación son percibidas como buenas o satisfechas en las tres comunidades (Figura 2). 230 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Alimentación Comunicación Organización 5 Estado de salud 4 Vivienda y vestimenta 3 Recreación Reproducción 2 1 Tradiciones y cultura Relación vecinos 0 Generación ingresos Relación intrafamiliar Elección de representantes Identidad con la comunidad Manejo de basura Felicidad Ambiente saludable Alto Quetzal Paso Marcos de S. Simiriñak Figura 2. Relación entre Medios de Vida y Comunidades de Alto Chirripó (Alto Quetzal, Paso Marcos de Simiriñak y Simiriñak). El capital natural es uno de los más importantes para la población Cabécar, el bosque y la agricultura juegan un papel primordial dentro de la reproducción familiar, además tiene una relación directa con aspectos culturales y biológicos. La seguridad alimentaria depende del estado de este capital y de cuando este aporte a la salud y nutrición de las familias indígenas de la zona; existe una gran diversidad de productos agrícolas, pese a esto la diversidad de dietas es baja por lo que su alimentación se limita al consumo de frijol, banano y arroz; las prohibiciones respecto al uso de productos del bosque como la cacería han conducido a cambios en los patrones alimentarios. Análisis de la seguridad alimentaria en comunidades Cabécares de Alto Chirripó La seguridad alimentaria existe cuando todas las personas tienen, en todo momento, acceso físico, social y económico a alimentos suficientes, inocuos y nutritivos que satisfacen sus necesidades energéticas diarias y preferencias alimentarias para llevar una vida activa y sana (Cumbre Mundial sobre la Alimentación, 1996). a) Disponibilidad Diversidad de alimentos producidos y adquiridos por las familias indígenas de Alto Chirripó Según las investigaciones realizadas por López (2004) y Ríos (2011), mencionan que las comunidades indígenas de Alto Chirripó se dedican a la producción agrícola y pecuaria empleada básicamente para autoconsumo, así disponen de granos básicos, tubérculos tropicales, frutales y algunas hortalizas. La presente investigación permitió identificar que cada familia dispone entre 3 y 20 cultivos siendo los cultivos básicos: maíz, frijol y banano (Figura 3). 231 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 ñampi tiquizque ñame malanga yuca hortalizas chayote chirimoya caña de azúcar guaba cas mandarina piña papaya guanábana cacao aguacate café l_dulce naranja l_agrio plátano pejibaye banano maíz frijol 0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100% Figura 3. Diversidad agrícola presente en las comunidades de Alto Quetzal, Paso Marcos de Simiriñak y Simiriñak Las ayudas alimentarias proporcionadas por el Gobierno, si bien por un lado solventan las necesidades alimenticias y nutricionales, también han sido considerados agentes de cambio para los patrones culturales de las zonas indígenas; se evidencian el alto consumo de enlatados, embutidos, pastas y gaseosas o frescos de paquete. b) Acceso En términos de acceso a tierra, las familias indígenas de Alto Chirripó, disponen entre una y 50 ha, distribuidas en distintas zonas, estos territorios son destinados para vivienda, producción y conservación del bosque. Al mantener la tradición de herencia matriarcal, la redistribución de tierras se asigna por parte del padre de la mujer quien entrega sus tierras a la nueva pareja, la misma que puede estar cerca o lejos del rancho de la familia materna (Ministerio De Salud, 2005). El uso de productos del bosque como la extracción de hongos, hojas, la caza son actividades que se han ido perdiendo a nivel de la zona indígena, las razones son múltiples, por un lado está las prohibiciones respecto a la cacería y por otra la falta de interés que demuestran las familias ante estas actividades. El uso de plantas medicinales es algo que aún conservan, el conocimiento respecto a la forma de preparación y uso está dado más en las personas adultas que en los jóvenes. c) Consumo y/o utilización biológica Diversidad de dietas en las familias indígenas de Alto Chirripó La encuesta de recordatoria de 24 horas (HDDS), permitió conocer la diversidad de dietas presentes en la zona; de 12 grupos alimenticios, se identificó que las familias consumen entre cinco y 12 (Cuadro 1), es decir que poseen de mediana a alta diversidad de alimentos, con un promedio de ocho grupos alimenticios por familia, esto da a entender que las familias indígenas poseen dietas diversificadas. Según Swindale y Bilinsky (2006) la diversidad de dietas es un indicador indirecto (proxy) que demuestra que una dieta más diversificada está asociada con buenos pesos de nacimiento, medidas antropométricas, concentraciones de hemoglobina, además de tener una correlación con factores como ingesta calórica y proteínica así como con los ingresos familiares. Este tipo de encuestas permite analizar la seguridad alimentaria desde un punto de vista familiar o intrafamiliar. 232 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Cuadro 1. Cantidad de grupos alimentarios presentes en 25familias entrevistadas en las comunidades de Alto Quetzal, Paso Marcos de Simiriñak y Simiriñak N° grupos alimenticios 8 a 12 Categorías Alta diversidad de alimentos Media diversidad de alimentos 5a7 Baja diversidad de alimentos 1a4 N° familias 19 6 Total 25 Fuente: Elaboración propia A pesar de mostrar que la dieta es diversificada, existe la tendencia hacia alto consumo de carbohidratos, dados por el aporte de cereales donde se destaca el consumo de arroz (96%) y frijol en un 92%, en algunos casos se mencionan productos como pan, galletas, maíz y sus derivados. En general para los costarricenses la base de la alimentación es el arroz y los frijoles, esta combinación permite obtener proteína de buena calidad, similar a la de la carne y a bajo costo, por lo que se recomienda consumir por cada cucharada de frijol dos de arroz (Ministerio de Salud 2010), además que el frijol es considerado como un alimento funcional por ser fuente de proteínas, almidones, hierro, ácido fólico, magnesio y potasio y de fibra (Sedó Masís 2002). Estado nutricional según el Índice de Masa Corporal (IMC) en los grupos de estudiantes de Alto Chirripó El estado nutricional del grupo evaluado está dentro de los rangos de normalidad, el déficit nutricional está presente en el 3% y 2% de mujeres y hombres respectivamente; sobrepeso es más notorio a nivel de mujeres que de hombres y los problemas de obesidad está presente en menos del 7% de la población en estudio. A nivel de grupos etarios, la presencia de sobrepeso es notorio dentro del grupo de mujeres en edades entre 15 y 19 años (24%) y también dentro del grupo de hombres en edades de 5 a 9 años (18%); problemas de nutrición (infrapeso) está presente en el 6% tanto de hombres como mujeres en edades de 10 a 14 años (pubertad) (Figura 4). 100% 88% 86% 77% 73% 24% 18% 5% G1 (5-9 años) M 67% 6% 3% 6% 6% 9%12% 4% 8% 10% G1 (5-9 años) G2 (10-14 años) G2 (10-14 años) G3 (15-19 año) G3 (15-19 años) F M Infrapeso F Normal Obesidad M F Sobrepeso Figura 43. Estado nutricional por sexo y edad según IMC en la zona de Alto Chirripó El estado nutricional en personas adultas fue medido únicamente a nivel de la comunidad de Alto Quetzal y se tomó como referencia estudiantes del colegio (10 hombres y nueve mujeres) que están en edades entre 20 y 34 años; el principal problema observado fue en el grupo de mujeres donde el 56% presenta obesidad tipo I, y los valores de sobrepeso y normalidad están por debajo del 25%; para el caso de los hombres hay un 50% que está en estado normal y un 50% que presenta problemas de sobrepeso. 233 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 En general la muestra evaluada presenta un estado nutricional normal en todas los grupos etarios, existe dos casos tanto para hombres como mujeres con problemas de delgadez; según el informe de la CCSSTurrialba dentro del territorio indígena Cabécar de Chirripó la frecuencia de desnutrición infantil afecta de alguna manera al 40% de los niños menores de 6 años y hasta un 50% de la población escolar (Aragón y Garro 2010). Si bien los problemas de desnutrición están presentes más en niños menores de 6 años, los problemas de sobrepeso y obesidad también están presentes a nivel de zona indígena debido al cambio de la estructura alimentaria tradicional, factor que está asociado a la incorporación de la alimentación escolar con productos no locales, lo que ha provocado cambios en las dietas y preferencias alimenticias por lo que es cada vez más común ver el consumo de enlatados, embutidos, comida cada vez más condimentada y refrescos de paquete, entre otros. d) Estabilidad Acceso permanente a alimentos Se tiene estabilidad cuando hay acceso a alimentos adecuados y en todo momento, el almacenamiento es una de las actividades que promueven la seguridad alimentaria, así por ejemplo el 80% (30) de las familias entrevistadas guardan semillas de frijol y maíz, los granos cosechados son almacenados en sacos y el tiempo de uso dependerá de si realizan o no un segundo ciclo de cultivo y la frecuencia de consumo de los mismos; en el año existe disponibilidad continua de banano, plátano y varios tubérculos tropicales y también de productos estacionales como ciertos frutales (naranjas, limón dulce, entre otros). BIBLIOGRAFÍA Aragón, V; Garro, J. 2010. Análisis de la Situación de Salud: Territorio indígena Cabécar de Chirripó. Meneses, L. Turrialba, CR, Caja Costarricense de Seguro Social (CCSS). 72 p. FAO, (Organización de las Naciones Unidas para la Agricultura y la Alimentación) 1996. Cumbre Mundial sobre la alimentación: Declaración de roma sobre la Seguridad Alimentaria Mundial y Plan de Acción sobre Alimentación 1996. (en línea). Disponible en http://www.fao.org/wfs/index_es.htm ______2009. Pueblos indígenas y tribales: construyendo la diversidad biológica y cultural para la seguridad alimentaria y de los medios de vida. El tesoro alimentario de los indígenas en peligro (en línea). Disponible en http://www.fao.org/news/story/es/item/29629/icode/ López, M. 2004. Papel de los productos forestales en las estrategias de vida de los Indígenas Cabécar, Cantón de Turrialba, Costa Rica. Tesis Mag Sc. . Turrialba, Costa Rica, CATIE. 116 p. Ministerio de Salud, C. 2005. Aproximación a la condición de salud los Pueblos Indígenas de Costa Rica.10 p. Disponible en ttp://www.ministeriodesalud.go.cr/ops/documentos/docPerfil%20de%20Salud%20%20de%20Pue blos%20Indigenas%20de%20CR.pdflos José Van der Laat A. ________. 2010. Análisis de Estado Nutricional Población Infantil del Distrito de Chirripó. Informe área de salud Turrialba-Jimenez.7 p. Montoya, F; Drews, S. 2007. Medios de vida, bienestar comunitario y conservación de especies. Una guía para entender, evaluar y mejorar los vínculos en el contexto de los programas de tortugas marinas. WWF. Programa marino y de Especies para Latinoamérica y el Caribe. San José, CR 234 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Ríos, S. 2011. Vulnerabilidad al cambio climático de tres grupos de productores agropecuarios en el área de infuencia del Bosque Modelo Reventazón (BMR), Costa Rica. Tesis Mag. Sc. Turrialba, CR, CATIE. 134 p. Sedó Masís, P. 2002. El mercado de los alimentos funcionales y los nuevos retos para la educación alimentaria - nutricional. Revista Costarricense de Salud Pública 11:18-25. Consultado 9 oct. 2012. Disponible en http://www.scielo.sa.cr/scielo.php?script=sci_arttext&pid=S140914292002000100004&nrm=iso Swindale, A; Bilinsky, P. 2006. Puntaje de Diversidad Dietética en el Hogar (HDDS) para la Medición del Acceso a los Alimentos en el Hogar: Guía de Indicadores (v.2). Proyecto de Asistencia Técnica sobre Alimentos y Nutrición (FANTA), Academia para el Desarrollo Educativo (AED):40 p. 235 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 BIOINDICADORES: “SEGUROS LOCALES” PARA PEQUEÑOS PRODUCTORES AGROPECUARIOS Galo Fernando Pillajo Anchatuña Ministerio de Agricultura, Ganadería, Acuacultura y Pesca, Subsecretaría de Ganadería [email protected] Introducción La agricultura necesita adaptarse en muchas zonas a cambios rápidos e intensos del clima (temperaturas, precipitaciones) y a cambios relacionados con los esquemas de distribución de plagas y enfermedades. Mientras en algunas zonas los rendimientos podrían inicialmente mejorarse, se espera que el impacto general del Cambio Climático en el sector agropecuario y en la Soberanía y Seguridad alimentaria sea cada vez más negativo, especialmente en zonas propensas a sufrir desastres relacionados con el clima como sequías, heladas e inundaciones, y a padecer inseguridad alimentaria. Los pequeños productores agropecuarios identifican signos y comportamientos en las plantas, animales y astros que pudiésemos denominarlos “predictores del clima” ó “Bioindicadores”. Estos predictores están sustentados en la Cosmovisión del Universo. El productor agropecuario, especialmente el Andino (léase habitante de los países cruzados por la Cordillera de los Andes), da valor y significados al comportamiento de las plantas, los animales, el cielo, la tierra –entre otros– con los que establece un diálogo, de modo que en él se desarrolla un conocimiento predictivo de los cambios súbitos de clima. Según Patrick Stewart, biólogo y consultor ambiental (1990), un Bioindicador es un organismo que funciona como una señal o calendario, indicando factores ambientales, como los cambios de estación o la proximidad de un recurso vital como las lluvias. Este reconocimiento a la sabiduría ancestral junto a la moderna tecnología de predicción climática pueden usarse como estrategia básica y alternativa que permita prevenir las pérdidas de recursos y vidas humanas que ocasionan las Variaciones Climáticas y Cambio Climático y de esta manera convertirse en “Seguros Locales” para las poblaciones más pobres y vulnerables de las zonas rurales ecuatorianas. Resultados Este proceso de sistematización ha permitido identificar Bioindicadores (llamados también indicadores biológicos o climáticos) en base al conocimiento ancestrales en altitudes desde los 8 a 3476 metros sobre el nivel del mar, que van desde la región litoral (provincias de Manabí, Guayas, Los Ríos) y Amazónica (provincia de Orellana) hasta la provincia de Chimborazo. Se identificaron en total 369 Bioindicadores, la mayoría, es decir el 36% de ellos corresponden a comportamientos de animales, el 35% al de los astros y otros elementos meteorológicos y el 29% restante, al comportamiento de las plantas. 236 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Gráfico Nº 1: Porcentajes totales de Bioindicadores de la sistematización La edad promedio total de los informantes fue de 51,24 años El mes representativo para interpretar el comportamiento de las plantas como Bioindicadores, es octubre para la región amazónica e interandina y diciembre para la región del litoral. El mes de noviembre es el mas representaivo para interpretar el comportamiento de los animales en la región interandina y litoral y para la región amazónica, en cualquier época del año. Los meses más representativos para interpretar el comportamiento de los astros y otros elementos meteorológicos son: noviembre para la región interandina, diciembre para la región litoral y en cualquier época para la región amazónica. Ejemplos de Bioindicadores identificados: PLANTAS ELEMENTO BIOINDICADOR EPOCA ceibo "vestido" enero palma real octubre árbol de tamarindo capulí tilo "tangarana" (árbol) QUÉ OBSERVAR árbol con abundantes hojas de color verde caña 3 gajos de coquitos o corozo INTERPRETACIÓN Invierno próximo a iniciar Invierno próximo a iniciar noviembre árbol "vestido" de En mes y hojas verde claro medio empieza el invierno octubre floración en inicio de las árboles jóvenes lluvias (menos de 15 años) octubre, floración en inicio de las noviembre árboles lluvias noviembre hoja se hace rojo se acercan las 237 NOMBRE EDAD Miguel Ángel Conforme Cuzme 59 Walter Farías Jiménez 42 Germán Cayetano Zambrano Villa Francisco Guamán Caranqui 45 José Yausen León Fermín Tapuy 75 75 54 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 lluvias ASTROS Y OTROS ANIMALES "caperona" (árbol) "gallareta azul" -ave enero caída de hojas marzo diciembre presencia presencia del Eduardo Pozo verano El invierno es Edison Neptalí fuerte cuando Zúñiga llagan éstas Sánchez aves por ciento por las noches "cordeleros"- libélulas enero volando bajo y en se inicia el Eduardo Mera abundancia invierno y va a Lucas ser regular alacranes noviembre, abundantes el invierno va a Gladis Antonia diciembre, alacranes en los ser de lluvias Mendoza enero tallos del plátano normales y se Solórzano extenderá hasta el mes de mayo golondrina noviembre gran cantidad época de Francisco siembra de Guamán habas, ocas, Caranqui papas, mashua "Ucuy" septiembre- salen de sus nidos presencia de Fernando ( hormiga arriera) noviembre lluvia Alvarado 7 estrellas en Cuando se forma de una alinean las "7 cabrillas"- octubre, Héctor José cabrilla estrellas, se estrellas noviembre Basurto aposentada en el aproxima el valle invierno " color negro, significa lluvia fuerte, sin viento; Fernando nubes invierno color de nubes color blancuzco, Alvarado cae lluvia y hace viento" Invierno está Las aguas noviembre lejos de marinas Jorge Orlando mar , empezar empiezan a bajar Bravo Lucas enero invierno de de temperatura pocas lluvias invierno noviembre María altas próximo a mar , Genoveva Ávila temperaturas entrar y mucha diciembre Farías lluvia Conclusiones 238 55 44 36 35 75 47 30 47 38 80 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 • Los Bioindicadores climáticos deben ser recopilados, sistematizados, validados, difundidos y puestos en práctica. El reto en los próximos años es recuperar este capital social comunal del sector rural y con la participación de otros actores locales, verificar si son válidos para otros territorios del sector agropecuario asentados en las zonas más vulnerables a los impactos negativos del clima. • Los Bioindicadores ancestrales son complementarios a las predicciones climáticas de los servicios nacionales de meteorología emitidos por INAMHI, CIIFEN, INOCAR, etc., pero en la lógica de acción-reacción ante eventos repentinos a nivel local, logran tener un nivel de efectividad dada la deficiencia de la comunicación en tiempo real de las alertas meteorológicas. • El conocimiento tradicional se convierte en este contexto en un mecanismo que permite a las poblaciones rurales disponer de avisos climáticos ante cualquier tipo de variación hidrometeorológicas. Literatura citada Terminología sobre Reducción del Riesgo de Desastres. Estrategia Internacional para la Reducción de Desastres. Naciones Unidad, 2009. UNISDR. Mauricio G. Paoletti. Departamento de Biología Universidad de Padova, vía U. Bassi, 58 / b, 35121 Padova, Italia. Vallejo, S; Quingaísa, E; Ortiz, P; Vinueza, L. 2007. El Agro y Vida Rural en Ecuador: Comportamiento 2000-2007 y Perspectivas 2008, IICA, Quito-Ecuador. Ministerio de Relaciones Exteriores del Ecuador, 2007. El Cambio Climático en Ecuador. 2007. Valdivieso, E. Quispe, M. Aguilar, L. Metodología de pequeños productores para mejorar la producción agrícola. Capacidades y Estrategias locales para la gestión de riesgos. Fundación Agrecol. Andes. La Paz-Bolivia. Junio, 2006. 239 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 SISTEMAS INTEGRADOS: SISTEMAS AGROFORESTALES Antonio Vera EVALUACIÓN DE ESPECIES FORRAJERAS BAJO SISTEMAS SILVOPASTORILES PARA LA SOSTENIBILIDAD DE LA GANADERÍA EN LA AMAZÓNÍA ECUATORIANA Gloria Cristina Luna MATERIAL DIDÁCTICO PARA EL APRENDIZAJE AGROFORESTAL Ingrid Martínez ESTABLECIMIENTO DE ESPECIES ARBÓREAS DE USO AGROFORESTAL MEDIANTE TÉCNICAS CONSERVACIONISTAS EN CONDICIONES DE SECANO Jenny Ordóñez ENLAZANDO CONOCIMIENTO LOCAL Y RASGOS FUNCIONALES DEL COMPONENTE ARBÓREO, BANANOS Y PALMAS EN SISTEMAS AGROFORESTALES CON CACAO Y CAFÉ EN COSTA RICA Juan F. Dueñas RUNA GUAYUSA – DESARROLLO DE UN SISTEMA DE CULTIVO AGROFORESTAL DE Ilex guayusa Loes. Nancy Criollo EVALUACIÓN DE ALTERNATIVAS SILVOPASTORILES QUE PROMUEVAN LA INTENSIFICACIÓN Y RECUPERACIÓN DE PASTURAS DEGRADADAS Y CONTRIBUYAN A REDUCIR EL IMPACTO AMBIENTAL DE LA ACTIVIDAD GANADERA EN LA AMAZONÍA ECUATORIANA AL SEGUNDO AÑO DE ESTABLECIMIENTO 240 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Aliana Piñeiro Ilex Guayusa COMO MOTOR PARA EL DESARROLLO SOSTENIBLE EN LOS SISTEMAS AGROFORESTALES EN LA AMAZONÍA OCCIDENTAL Roy Vera MEJORAMIENTO DE CHAKRAS, UNA ALTERNATIVA DE SISTEMA INTEGRADO PARA LA GESTIÓN SOSTENIBLE DE BOSQUES EN COMUNIDADES NATIVAS DE LA AMAZONÍA ECUATORIANA 241 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 EVALUACIÓN DE ESPECIES FORRAJERAS BAJO SISTEMAS SILVOPASTORILES PARA LA SOSTENIBILIDAD DE LA GANADERÍA EN LA AMAZÓNÍA ECUATORIANA Antonio Vera, Willan Caicedo, Luis Riera Instituto Nacional Autónomo de Investigaciones Agropecuarias, Estación Experimental Central de la Amazonía. Vía san Carlos a 3 km de la entrada a La Parker, Cantón Joya de los Sachas, Orellana. [email protected] Resumen En la Estación Experimental Central de la Amazonía del INIAP ubicada en el cantón La Joya de los Sachas, Provincia de Orellana, se evalúan especies forrajeras bajo sistemas silvopastoriles (SSP), como alternativa para la sostenibilidad de los recursos naturales y mejorar la productividad de los ganaderos promoviendo la recuperación de pasturas degradadas, contribuyendo a reducir el impacto ambiental en la Amazonía ecuatoriana. Las especie evaluadas son 12 gramíneas para pastoreo; 6 gramíneas de pastoreo 3 leguminosas herbáceas y 6 forrajeras leñosas; cada grupo se evaluaron bajo un diseño de Bloques Completos al Azar con tres repeticiones. Transcurrido tres años de evaluación las que presentan mayor producción y adaptación a las condiciones de la Amazonía son las siguientes: Brachiaria hibrido (mulato); Brachiaria brizantha (xaraes); Panicum maximun (mombaza) en pasto de corte King Grass (morado y blanco) y Pennisetum violaceun (maralfalfa); en leguminosas herbáceas Arachis pintoi (maní forrajero); y en especies forrajeras leñosas Morus alba (morera); Flemingia macrophylla (flemingia) y Leucaena leucocephala (leucaena); las gramíneas y leguminosas herbáceas han sido evaluadas en tres frecuencias de corte a los 30, 45 y 60 días y las forrajeras leñosas a los 60, 90 y 120 días. Concluyendo que existe potencial de diferentes materiales dentro de cada grupo. Introducción Las pasturas constituyen el principal uso productivo del suelo en la Región Amazonía Ecuatoriana (RAE), dominadas por pasturas en monocultivos y degradadas por malas prácticas desde el establecimiento y posterior manejo. La actividad ganadera es el principal medio de vida de más de 3000 familias en esta región (Nieto y Caicedo 2012). En el caso de la ganadería bovina, los niveles de producción y productividad son bajos: los promedios de producción de leche apenas alcanzan a 3.5 litros/vaca/día y la producción de carne acusa incrementos de apenas 0,25 kg/día, con capacidad de carga animal de 0.8 unidades bovinas adultas/ha (INIAP y MAGAP, 2010). Esta baja productividad de la ganadería bovina en la RAE, se explica entre otras, por las siguientes causas: baja fertilidad de los suelos y en procesos de degradación que tiene como consecuencia una baja productividad de biomasa de pasturas; las especies y variedades de pastos utilizados son susceptibles al ataque de plagas; poca o nula utilización de leguminosas (arbustivas o rastreras), que mejoren la fertilidad del suelo y disminuyan su erosión; presencia agresiva de malezas, que ocasiona un alto uso de la mano de obra; uso de razas o cruza de ganado de bajo potencial productivo y prácticas deficientes de manejo reproductivo y sanitario del ganado (INIAP y MAGAP, 2010). Todo lo anterior mencionado está relacionado con la deforestación y la emisión de gases de efecto invernadero considerados entre los factores más importantes de cambios climáticos globales (Wood y Porro 2002, Grijalva et al. 2004). 242 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Por lo manifestado, es muy necesario conocer especies forrajeras de buena producción de biomasa que se adapten a los diferentes ecosistemas y permite tener alternativas para la implementación de sistemas agro y silvopastoriles en la región amazónica, razones por las cuales el INIAP emprendió un programa de generación de tecnologías con especies forrajeras herbáceas y leñosas con el propósito de implementar alternativas silvopastoriles orientadas a la mejora de los sistemas de producción incrementando los niveles de producción y productividad pecuaria, contribuyendo a reducir el impacto ambiental de la actividad ganadera en la Amazonía ecuatoriana, Vera, A.1999 Materiales y Métodos La presente investigación se realiza en la Estación Experimental Central de la Amazonía (EECA), del INIAP, en la parroquia San Carlos, cantón La Joya de los Sachas, Provincia de Orellana, con una altitud de 249 msnm, precipitación de 3000 mm anual y una temperatura de 24 °C; bajo sistema silvopastoril que fue establecido en el año 2010 en la EECA; en una área que estuvo ocupada por más 10 años por el cultivo de limón, la que cumplió su ciclo de producción, en esta área se realizó una limpieza total de malezas dejando algunos árboles de especies maderables y limón. Para el establecimiento de las parcelas se realizó una recolección de las especies forrajeras a nivel de la Amazonía y Costa. El material recolectado fue sembrado por semilla sexual a una densidad de 0.8 x 0.8 m, y por semilla asexual las especies de corte y las leñosas a una densidad de 1 x 1 m, transcurridos los seis meses de establecidas las parcelas se realizó un corte de igualación a las pastura y se establecieron los corte con frecuencias de cortes a los 30, 45 y 60 días en las herbáceas y 60, 90 y 120 días en las leñosas. Las parcelas están implementadas bajo un Diseño de Bloques Completos al Azar con tres repeticiones por tratamiento, en un área experimental de 1 hectárea. Los datos experimentales obtenidos se tabularon y se analizaron utilizando el software InfoStat (2012). Se realizaron análisis de varianza y pruebas de Tukey (al 5%), como prueba de significación los tratamientos del experimento se muestran a continuación. Cuadro 1. Especies de gramíneas y leguminosas existente en el banco forrajero. EECA-2013. Gramíneas N. Común Leguminosas herbáceas N. Común Brachiaria decumbens Dallis, Arachis pintoi Maní forrajero Brachiaria brizanhta Marandu Clitoria sp Clitoria Brachiaria brizantha Xaraes Desmodium ovalifolium Trébol tropical Brachiaria hibrido Mulato Desmodium heterophyllum Desmodium Brachiaria humidicola Kikuyo amazónico Centrosema pubescens Centro Brachiaria brizantha Piata Macroptilium atropurpureum Siratro Brachiaria dictyoneura Llanero Stylosanthes guianensis Alfalfa tropical Panucum máximum Común Leguminosas leñosas 243 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Panucum máximum Enano Flemingia macrophylla Flemingia blanca Panucum máximum Mombaza Leucaena leucocephala Leucaena Panucum maximum Tanzania Trichantera gigantea Quiebra barriga Panucum maximum Pichilingue Gliricidia sepium Yuca de ratón Pennisetum purpureum Elefante Morus alba Morera Pennisetum violaceum Maralfalfa Flemingia macrophylla Flemingia morada King grass. Morado Tithonia diversifolia Botón de oro King grass. Verde Pennisetum 811 Pichilingue No identificado cuenca Se evaluaron las siguientes variables a) Producción de biomasa. El corte de las especies se realizó cada 30, 45 y 60 días de acuerdo al tratamiento, lo que se utilizó un cuadrante de 1 m2 y tomando 3 muestras por parcela, en cada tratamiento se determinó la producción de la herbácea; mientras la producción de forraje de las leñosas se determinó mediante la simulación del ramoneo de los animales cosechando la biomasa disponible en leñosas forrajeras previamente identificadas. Del material producto de las producciones se tomó una muestra representativa de 500 g y se envió al laboratorio de calidad y alimento del INIAP, para la determinación de materia seca, proteína cruda, digestibilidad, para poder observar las cualidades de cada una de las especies forrajeras en las diferentes frecuencias de corte. Resultados y Discusión Especies de pastos para pastoreo. El análisis de varianza mostró diferencias estadísticas en la producción de materia seca anual acumulada en los pastoreos registrados a los tres años de estudio, al comparar entre las especies. Transcurridos los tres años de evaluación, se observa la producción de biomasa y la que presenta los promedios de producción de forraje es el pasto xaraes (Brachiaria brizantha) con 1.14; 1.53 y 3.15 Ton/ha/año en frecuencias de 30, 45 y 60 días y mulato (Brachiaria hibrido) con 1.11; 1.96 y 3.1 Ton/ha/año en las tres frecuencias de corte. 244 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Gráfico. 1. Producción promedio de MS/ton/ha, de las especies de Brachiarias-EECA-.2013 En el comportamiento de proteína cruda el Brachiaria decumbens (dallis), presenta los mejores valores con 14.7, 12.5 y 11.2 en las diferentes frecuencias de corte sin embargo a los 30 días se evidencia mejor calidad, pero sin embargo si observamos a medida que aumenta la frecuencias de corte la calidad disminuye, lo ideal es aprovecharlos a los 40 o 45 días donde existe una buena producción de forraje, buen valor nutritivo. Grafico 2. Gráfico. 2. Comportamiento de la proteína en las especies de pastos de Brachiarias. EECA-2013 En digestibilidad se observa que a los 30 días presenta los mejores valores y a los 60 días disminuye drásticamente en todas las especies. Grafico 3. Porcentajes Digestibilidad (%) 70 Xaraes Mulato Marandú Dallis Kikuyo Amazónico Piata Llanero 60 50 40 30 30 días 45 días 60 días Frecuencia de aprovechamiento Gráfico. 3. Comportamiento de la digestibilidad en las especies de pastos de Brachiarias. EECA-2013 245 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Dentro de las especies de Panicum máximum, el de mayor producción es el cultivar mombaza con producción de materia seca Ton/ha/año de 0.97, 1.78 y 3.6 a los 30, 45 y 60 días. Gráficos 4. Gráfico. 4. Producción promedio de MS/ton/ha, de las especies de Panicum. EECA-.2013 En lo referente a proteína a los 30 días presentan los mejores valores y a los 60 días los valores más bajos, por lo tanto el aprovechamiento debe de realizarse a los 45 días donde se equilibre la producción con la calidad de las especies Gráfico 5. Gráfico. 5. Comportamiento de la proteína en las especies de Panicum. EECA-2013 En digestibilidad la tendencia es igual a la proteína entre mayor frecuencia de corte baja la digestibilidad exento el pasto tanzania que se mantiene en las tres frecuencias. Gráfico 6. 246 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Gráfico. 6. Comportamiento de la digestibilidad en las especies de Panicum. EECA-2013 Especies de pastos para corte. En las gramíneas de uso para corte, la mayor producción la acuso los pastos King grass morado y blanco con 0.78, 2.89, 7.04 y 1.17, 3.03 4.68 Ton/ha/año, respectivamente a los 30, 45 y 60 días Gráfico 7. Gráfico. 7. Producción promedio de MS/ton/ha, de las especies de pastos de corte. EECA-.2013 En proteína el King grass morados presenta los valores en las tres frecuencia sin mucha variabilidad con 11.14, 11.24 y 11.55 en cambio en las otras especies disminuye a los 45 y 60 días, lo que nos indica que se puede aprovechar cada 60 días cuando hay una mayor producción de forraje. Gráfico 8. Gráfico. 8. Comportamiento de la proteína en las especies de pastos de corte. EECA-2013 En digestibilidad tiene la misma tendencia que la proteína a medida que aumenta la frecuencia de corte disminuye el aprovechamiento por el animal. Especies de leguminosas herbáceas. En leguminosas herbáceas la que sobresale en producción en las tres frecuencias de corte es el maní forrajero con 0.86, 1.45 y 1.7 ton/ha/año. Gráfico 10. 247 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Gráfico. 10. Producción promedio de MS/ton/ha, de leguminosas herbáceas. EECA-.2013 En proteína la que mayor valor presenta es el Centrosema pubescens con 20.35, 20.50 y 20. 87 en las tres frecuencias de corte, pero sin embargo tiene problema con las frecuencias de corte. Gráfico 11. EECA-2013 Gráfico. 11. Comportamiento de la proteína en las leguminosas herbáceas. En digestibilidad el que presenta los mejores valores es el Arachis pintoi (maní forrajero), con 70.10, 70 y 69.3 en las tres frecuencias de corte, lo que lo hace una especie muy buena para los sistemas de producción sostenible. Gráfico 12. EECA-2013 248 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Gráfico. 12. Comportamiento de la digestibilidad en las leguminosas herbáceas. Especies de forrajeras leñosas. En las especies forrajeras leñosas la frecuencia de corte fueron a los 60, 90 y 120 días para observar la producción de forraje y calidad de cada una de las especies. La que presentó mayor producción fue Morus alba con, 2.64, 5.53 y 6.73 Ton/ha/año, en las tres frecuencias de corte, seguido por Flemingia macrophylla y Leucaena leucocephala). Gráficos 13. Gráfico. 13. Producción promedio de MS/ton/ha, de las especies de forrajeras leñosas. EECA-.2013 En proteína la que presenta los valores más altos es la Gliricidia sepium seguido de la Leucaena leucocephala con 30.2, 29.39, 27.02 y 29, 28.99, 26.42 a los 60, 90 y 120 días respectivamente. Gráfico 14. Gráfico. 14. Comportamiento de la proteína de las especies forrajeras leñosas. EECA-.2013 En digestibilidad los valores más altos los presenta la leucaena leucocephala y Gliricidia sepium. Gráfico 15. Porcentajes Digestibilidad (%) 90 Leucaena Gliricidia Flemingia Quiebra Barriga 70 50 30 60 días 90 días 120 días Frecuencia de aprovechamiento EECA-2013 249 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Gráfico.15. Comportamiento de la digestibilidad de las especies forrajeras leñosas. CONCLUSIONES Y RECOMENDACIONES Bajo las condiciones agroforestales en que se desarrolló la investigación, se establecen en base a los resultados de tres años de evaluacion se dan las siguientes conclusiones: Las especies de gramíneas para pastoreo que se presentan como opciones con mayor probabilidad de éxito para el desarrollo de los sistemas agro y silvopastoriles para la ganadería en la Amazonía ecuatoriana, debido a los mejores atributos en cuanto a rendimiento de forraje de la pastura, calidad, capacidad de adaptación y la persistencia a la frecuencia de cortes son (Brachiaria brizantha) cultivar xaraes, (Brachiaria hibrido) cultivar mulato y (Panicum maximun) cultivar mombaza.. En las especies de gramíneas para corte, King grass morado y blanco y maralfalfa (Pennisetum violaceum) por que presentan mayor producción de forraje, buena calidad, adaptabilidad y persistencia a las frecuencias de corte y se lo puede utilizar cada 60 días. En leguminosas herbáceas el maní forrajero (Arachis pintoi) es el que presenta mayor producción, adaptabilidad a la zonas media y baja, se asocia muy bien con la mayoría de gramíneas, es muy palatable, no causa timpanismo en los animales y tiene una buena calidad, persiste bajo sombra lo que es muy apropiado para los sistemas de producción ganadero. En las especies leñosas la de mayor producción es la morera y flemingia pero por su mejor calidad la leucaena es la de mayor recomendación por su buena adaptabilidad, resistencias a los cortes y calidad para ser utilizada en los sistemas agrosilvopastoriles, las otras especies funcionan muy bien como banco de proteínas, cercas vivas y mejorar la dieta animal e incentivar a la semiestabulación de los bovinos en la RAE. Con todas estas especies se tiene material suficiente para la implementación de los sistemas silvopastoriles y la sostenibilidad de la ganadería en la amazonía ecuatoriana. Se recomienda continuar la evaluación con animales para evaluar la productividad en carne y producción de leche. LITERATURA CITADA Caicedo, W. 2013. Evaluación de sistemas silvopastoriles como alternativa para la sostenibilidad de los recursos naturales, en la Estación Experimental Central de la Amazonía, del INIAP año 1. Tesis Ing. Zoot. Escuela Superior Politécnica de Chimborazo. Riobamba – Ecuador. Criollo, N. 2013. Evaluación de sistemas silvopastoriles como alternativa para la sostenibilidad de los recursos naturales, en la Estación Experimental Central de la Amazonía, del INIAP año 2. Tesis Ing. Agr. Escuela Superior Politécnica de Chimborazo. Riobamba – Ecuador. Grijalva, J., Arévalo, V., y Wood, Ch. 2004. Expansión y trayectorias de la ganadería en la Amazonía del Ecuador. Publicación miscelánea del INIAP. Editorial Tecnigrava, Quito Ecuador, p 201. 250 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Grijalva, J., Arévalo, V. y Barrera, V. 2002. Informe de investigación en el Valle de Quijos y Piedemonte del Ecuador. Proyecto IAI/NSF “Ganadería, uso de la tierra y deforestación en zonas amazónicas de Brasil, Perú y Ecuador: estudio comparativo”. U. de Florida-INIAP-UNAS-EMBRAPA-Universidad Federal do Pará-McGill University, p 99. INIAP, MAGAP. 2010. Mejoramiento de la productividad de los sistemas de producción de leche y carne bovina en áreas críticas de la costa, sierra y Amazonía ecuatoriana. Quito, Ecuador, 36 p. Nieto, C. y Caicedo, C. 2012. Análisis reflexivo sobre el Desarrollo Agropecuario Sostenible en la Amazonía Ecuatoriana. INIAP – EECA. Publicación Miscelánea Nº 405. Joya de los Sachas, Ecuador, p 102. Vera, A.1999. Proyecto “Rehabilitación de pastizales degradados y mejoramiento de la sostenibilidad de la ganadería en zonas intervenidas de la Amazonía Ecuatoriana”, Estación Experimental Napo-INIAP, Francisco de Orellana-Ecuador. Wood, Ch. y Porro, R. 2002. Deforestation and Land Use in the Amazon. Gainesville. University Press of Florida. p 385. 251 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 MATERIAL DIDÁCTICO PARA EL APRENDIZAJE AGROFORESTAL Gloria Cristina Luna, Hugo Ferney Leonel, Sandra Timaná Universidad de Nariño [email protected] Colombia es un país muy diverso en ambientes, climas y culturas, lo que permite que en cada región, la agroforestería ofrezca opciones ecológicas, sociales y económicamente sustentables. Especialmente los pequeños agricultores suelen implementar sistemas agroforestales para incrementar su eficiencia productiva, sin embargo, este conocimiento ha sido poco difundido, lo que requiere de una forma de facilitar la promoción, establecimiento y manejo de prácticas agroforestales, utilizando servicios especializados de extensión, con herramientas pertinentes para la educación de agricultores como el uso de las Tecnologías de la Información y la Comunicación – TIC y diversidad de materiales didácticos; en virtud de lo expuesto, se generó innovaciones didácticas para la construcción del conocimiento agroforestal, desde un enfoque de significancia y valoración social, donde el contexto local adquiere gran relevancia. PROBLEMA Los procesos de formación y capacitación a pequeños agricultores no son pertinentes a las condiciones socioculturales de las comunidades de la zona rural andina de Colombia, aspectos que limitan la apropiación de la Agroforestería como una alternativa de producción sustentable; debido a que las herramientas didácticas empleadas en los procesos de formación son escasas y se limitan a medios impresos y algunas prácticas de campo; aspectos que restringen una eficiencia en el aprendizaje significativo para la adopción de tecnología pertinente. El material didáctico constituye una de las alternativas para la aplicación de los principios de la sustentabilidad, en tanto, conlleven a facilitar los procesos de aprendizaje para la adoptabilidad y adopción de sistemas agroforestales. OBJETIVO GENERAL Crear herramientas didácticas para la adopción de sistemas agroforestales, en la zona andina del departamento de Nariño, Colombia. OBJETIVOS ESPECÍFICOS -Identificar las características socioambientales más relevantes para el desarrollo de material didáctico agroforestal. -Diseñar un material didáctico versátil que faciliten la comprensión de la Agroforestería, aplicable en procesos de capacitación de comunidades rurales. METODOLOGÍA El estudio se adelantó en el departamento de Nariño, municipio de Tangua, vereda Marqueza Alta con la participación de 18 familias de la comunidad rural. 252 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Se adelantó una Investigación Acción Participativa fundamentada en la interacción ser humano – naturaleza, considerando la lúdica como eje estratégico de acción para la construcción de conocimiento, con el apoyo de un equipo interdisciplinario conformado por: Ingenieros agrónomos, forestales, agroforestales, diseñadores gráficos, licenciados en informática y publicistas. En este sentido con el enfoque participativo se desarrollaron las siguientes fases: Fase 1: Identificación de características socioambientales relevantes para el desarrollo de material didáctico Mediante reuniones con la comunidad de la vereda la Marqueza Alta y el grupo de investigación (Técnicos, extensionistas, profesionales, líderes y estudiantes) se socializó el proyecto y consolidó el compromiso de participación, para lo cual se estableció un cronograma de actividades considerando el rol de cada uno de los participantes. A partir de los intereses y necesidades socioambientales de los usuarios, de sus destrezas, habilidades y aptitudes se adelantó una revisión documental de fuentes secundarias (sociales, culturales y medioambientales) con su comprobación y validación en campo. Mediante talleres, empleando distintas técnicas y herramientas participativas como la lluvia de ideas, encuestas semiestructuradas, diálogos, recorridos de campo, el uso de ilustraciones y la aplicación de materiales didácticos, se evaluaron 9 variables (edad, sexo, ocupación del tiempo libre, conocimiento y preferencia de juegos de mesa, conocimiento de juegos tradicionales, preferencia de color, habilidad, preferencia del sistema de trabajo (Grupal o individual) y aspectos a considerar para recuperar la armonía en el grupo. Posteriormente, se categorizó las respuestas encontradas en las variables y se realizó el análisis de correspondencia múltiple, a través del método el agrupamiento de variables (Clúster), empleando el software estadístico SPAD v 5.6 Fase 2: Diseño de material didáctico para agroforestería A partir de las características identificadas en la fase 1 se planteó la identidad del material didáctico que fue perfeccionada con los aportes de un panel de expertos (equipo interdisciplinario). De igual manera con un equipo de expertos agroforestales se priorizó las temáticas agroforestales fundamentales e indispensables a ser empleadas en el proceso de capacitación de adultos para la adopción agroforestal. Una vez aprobada la identidad se diseñaron dos tipos de materiales: el material didáctico manual y una multimedia; en relación al primero se elaboraron los distintos bocetos de juegos, de los cuales se analizó su viabilidad con base en la aplicación de las temáticas agroforestales priorizadas, tras varias pruebas de juegos se logró seleccionar los de mayor pertinencia, los cuales se sometieron a un análisis de funcionalidad preliminar teniendo en cuenta las siguientes variables: 1. Diversidad de material: material creativo e innovador que en su conjunto atrae al usuario y facilita procesos de capacitación de los adultos. Fue necesario diseñar diversos materiales para los distintos temas determinados prioritarios para abordar la agroforestería. 2. Claridad en el contenido: material con mensajes claros, ideas concretas sobre los temas específicos que despertasen motivación e interés del usuario hacia la temática en cuestión. 253 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 3. Calidad del material: la calidad en términos de durabilidad, resistencia, diseño atractivo con ilustraciones claras, integrando imágenes y textos cortos con tipos de letra, tamaño y color de fácil comprensión. 4. Ergonomía: Se analizó el tipo de material con el cual se elaboró los juegos y sus empaques, dado que los extensionistas deben desplazarse a zona rural, lugar donde en su mayor porcentaje es difícil el acceso y los medios de transporte son escasos. Respecto al segundo tipo de material, la multimedia, se diseñó especialmente para jóvenes agricultores considerando que la utilización de la Tecnología de la Información y Comunicación - TIC refuerza las prácticas pedagógicas. Los prototipos finales fueron probados con los grupos de comunidades rurales mediante el desarrollo de talleres y se realizaron pruebas y ensayos de funcionalidad y acogida con los diferentes participantes, para finalmente realizar ajustes y contar con materiales de calidad. Todo el proceso fue registrado en forma escrita y audiovisual con el apoyo de los profesionales y estudiantes investigadores quienes evaluaron periódicamente cada fase para interpretar, ordenar y tabular datos a fin de sistematizar la experiencia. RESULTADOS Se logró una reconstrucción paisajística animada a partir del reconocimiento de los principales aspectos que despiertan interés y motivación hacia el medio de vida rural; considerando el aporte de la comunidad, de gran relevancia, tal como lo plantea Ausbel (1983), quien afirma que una exploración por los conocimientos previos de los habitantes de la zona de estudio permite analizar diversidad de aspectos que determinan su significancia en la concepción de materiales didácticos que facilitan a su vez la transferencia de desarrollos tecnológicos y el “aprender” nuevas relaciones socioculturales con sistemas productivos alternativos. El análisis de agrupamiento, permitió inferir de los 18 factores característicos de las personas encuestadas, que hay un corte principal conformado por 4 clusters y determinado por las variables: conocimiento y preferencia de juegos de mesa. (Figura 1). Cluster 1: Conformado por aspectos característicos y similares entre los individuos 1, 4, 17, 10 y 6; representados por la variable Conocimiento de juegos de mesa: “No sabe/ No responde” y la variable Preferencia de juegos de Mesa: “No conoce/ No le gusta”, con un porcentaje del 80%, (Figura 1). Cluster 2: Conformado por aspectos característicos y similares entre los individuos 14, 7 13, 16 y 3; los cuales afrman que el parqués, es su juego preferido de mesa como un factor determinante y un porcentaje representativo del 71,43%, (Figura 1). Cluster 3: Conformado por aspectos característicos y similares entre los individuos 12, 18, 8 y 9; este cluster esta representado por los individuos que afirman conocer el juego de cartas, sobre otro tipo de juegos de mesa, en su totalidad, (Figura 1). Cluster 4: Conformado por aspectos característicos y similares entre los individuos 2, 11 15 y 5; los que no tienen preferencia sobre un solo juego de mesa, sino también sobre otro tipo de juegos alternativos, (Figura 1). 254 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figura 1. Análisis de agrupamiento del conocimiento y preferencia de juegos de mesa de los 18 factores característicos de las personas encuestadas. 1 2 3 4 Considerando que tanto las fuentes primarias como secundarias funcionan como verificadoras que soportan la veracidad de la información, se tuvo en cuenta el análisis de contenido, el visual, la confiabilidad y validez; tal como lo plantea Galeano, (2007), identificando que las diferentes prácticas productivas cotidianas, son viables de redimensionar con innovaciones tecnológicas pertinentes, una vez se ilustran para visualizar opciones productivas; en este caso, desde una planificación agroforestal, aspectos que condujeron a propuestas de diseño gráfico preliminares. Se resaltó en la zona el quehacer cotidiano del hombre y mujer rural, identificando las características relacionadas con: topografía, recursos naturales, tipo de viviendas, rasgos faciales y vestimenta de los habitantes, como las de mayor relevancia para la construcción de identidad del material didáctico. De esta manera la identidad del material didáctico fue denominada agroforestería material didáctico, representada por don Braulio y doña Josefina una pareja de campesinos con su vestimenta característica y en un entorno rural. Aspectos de gran importancia para elevar autoestima de la comunidad y generar mayor apropiación del proceso, (Luna, et.al 2009). Por consiguiente los materiales didácticos que se diseñaron corresponden a dos tipos de medios de comunicación: uno el material didáctico manual que contiene cinco juegos y otro informático, que es una multimedia; los cuales permiten a los participantes interactuar y crear un ambiente de aprendizaje significativo, donde cada individuo es libre de expresar sus ideas, conocimientos y saberes, que son valorados por el grupo a fin de colectivamente construir nuevos conocimientos, que se perfeccionan al relacionarlos con las distintas situaciones reales de cada zona donde se desarrolla el taller, procurando enfatizar en el reconocimiento de las potencialidades de la región, para que los participantes descubran opciones de fácil emprendimiento, ( Berdegue, 1988) 255 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 “AGROFORESTERÍA - MATERIAL DIDÁCTICO”, es un conjunto de cinco juegos didácticos diseñados estratégicamente para la enseñanza - aprendizaje de la agroforesteria. Está dirigido a los agricultores y jóvenes tecnólogos y/o universitarios que incursionan en esta área del conocimiento. Es adecuado para trabajar con grupos de 20 personas aproximadamente. Siendo importante considerar que el extensionista – facilitador del proceso debe tener en cuenta los conceptos básicos agroforestales, conocer aspectos relacionados con andragogía, participación comunitaria y uso de la lúdica, a fin de orientar adecuadamente su uso y lograr el resultado deseado. El material didáctico contempla cinco temáticas que representan los aspectos generales para emprender acciones de desarrollo agroforestal en las fincas; compuesto por: una maleta viajera con su logotipo de identidad que contiene: guía del facilitador y cinco morrales con cada juego. GUIA DEL FACILITADOR: Orienta a quienes utilicen el material didáctico agroforestal para una mayor eficiencia en el trabajo de campo. MORRALES: Diseñados ergonómicamente para su transporte a campo, son livianos, de fácil transporte y elaborados en lona impermeable, resistente y de alta durabilidad. Cada morral contiene un juego con su respectiva ficha de instrucción; que permite orientar a los participantes en la comprensión de cada una de las temáticas agroforestales. JUEGOS: 1.Juego: SENDERO DEL CONOCIMIENTO: Considerando que son muchos los sucesos que han marcado la historia agroforestal, se ha diseñado un contexto natural a manera de un gran territorio que ilustra los principales hitos del desarrollo agroforestal en relación a su definición y función. 2. Juego: RIQUEZA AMBIENTAL: Mediante un rompecabezas con un paisaje típico de la zona andina, se resalta los beneficios del árbol como especie multipropósito, que brinda bienes y servicios ecológicos y socioeconómicos, destacando las ventajas y desventajas de su uso y manejo en los distintos sistemas de producción. 3. Juego: ClasiSAF: Es una ruleta que gráficamente muestra los componentes agroforestales y sus interacciones que conllevan a su clasificación. 4. Juego: ARREGLOTON: Este juego representado en un árbol y su interacción con el suelo, en un área determinada, facilita el diseño de diferentes arreglos agroforestales. Los procesos de degradación de suelos andinos, han contribuido a que los arreglos agroforestales se conviertan en un gran tesoro para el desarrollo sustentable y en el repensar tanto de académicos como agricultores para la implementación de alternativas de uso y manejo espaciotemporal adecuadas a las condiciones socioambientales actuales. 5. Juego: PLANIFICANDO ANDO: El juego consiste en el uso de maquetas que representan terrenos con distintas condiciones topográficas, suelos y agua; planificadas de acuerdo a los recursos económicos con que se cuenta. Este juego permite conocer los lineamientos requeridos para la planificación de fincas, que a su vez, se encuentra articulado a los principios del manejo de cuencas hidrográficas. De esta manera con “Agroforestería – material didáctico” se logró estructurar una herramienta de trabajo que es auto educativa, auto correctiva y auto reforzadora; considerada como auxiliar del extensionista en la impartición de sus capacitaciones y tienen como característica que proporciona al usuario una experiencia directa o indirecta de la realidad. Como auxiliares, los recursos están supeditados a la acción de quien los maneja y serán tan efectivos en tanto que la persona que hace uso de ellos sepa cómo aprovecharlos al máximo para conseguir los objetivos que se plantean en la planeación de cada sesión de trabajo. 256 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 La multimedia “MUNDO AGROFORESTAL” es un medio interactivo, con alto grado de versatilidad, fue diseñada especialmente para jóvenes rurales y pretende guiar y motivar al usuario hacia la comprensión de la dimensión agroforestal. Contempla tres interfaces que de manera organizada y lógica conllevan a un aprendizaje significativo en la medida en que es gradual y secuenciado, y que estimula el descubrimiento de soluciones agroforestales a problemáticas socioambientales, se caracteriza por tener un nivel de dificultad adecuado al nivel de conocimientos y habilidades del usuario, contener estrategias de instrucción que son ayudas pedagógicas que facilitan la comprensión del texto y finalmente por estar contextualizada en la realidad de Nariño. Los materiales didácticos diseñados consideraran los postulados de Freire, 2005 quien plantea “El extensionista-educador que no conoce el mundo del campesino no puede tratar de cambiar la actitud de este último. La intención de Freire era destacar los principios y fundamentos de una educación que fomente la práctica de la libertad. Esta práctica no debe reducirse a un simple apoyo técnico, sino incluir el esfuerzo personal de descifrarse a sí mismo y a los demás”. En este tipo de trabajo se utilizaban técnicas como grupos de estudio, grupos de acción, mesas redondas, debates y tarjetas temáticas. De esta forma, Freire y sus colaboradores empezaron a hablar de un “sistema” de técnicas de educación, del “sistema Paulo Freire” que podía aplicarse a todos los niveles de la enseñanza formal y no formal, tal como se desarrolló en esta zona de trabajo del departamento de Nariño. La interacción de la perspectiva de Feverstein y la teoría de la modificabilidad estructural cognitiva con el análisis del contexto socioambiental particular y la implementación de material didáctico diseñado participativamente, permitieron lograr aprendizajes significativos; lo que conllevó a que la comunidad rural se motivara, empoderara y apropiara del conocimiento; lo que a su vez facilitó la adoptabilidad de sistemas agroforestales. BIBLIOGRAFÍA AUSUBEL, N. 1983.Psicología Educativa: Un punto de vista cognoscitivo. 2° Ed. Trillas México. BERDEGUE, J; LARRAIN, B. 1988.Cómo trabajan los campesinos. Ed. Celater. Colombia. FREIRE, P. 2005. Pedagogía del oprimido. Trad. por Jorge Mellado. 2a. ed. México, Siglo XXI, 246 p. GALEANO, M. 2007. Estrategias de investigación social cualitativa. La Carreta Editores E.U. Medellín. Co. 240 p. LUNA, C; MOLINA, A; TORRES, F. 2009. Maíz y Frijol, Cultivos De Tradición, Ed. Editorial Universitaria Universidad de Nariño. Co. 100 p. 257 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 ESTABLECIMIENTO DE ESPECIES ARBÓREAS DE USO AGROFORESTAL MEDIANTE TÉCNICAS CONSERVACIONISTAS EN CONDICIONES DE SECANO Ingrid Martínez G., Carlos Ovalle, Hamil Uribe, Christian Prat, Alejandro Del Pozo, Natalia Valderrama, Germán Ruíz Instituto de Investigaciones Agropecuarias. Av. Vicente Méndez 515. Chillan, Chile [email protected] Introducción En Chile, las zonas de clima mediterráneo se caracterizan por suelos altamente erosionados. En estas condiciones la disponibilidad hídrica para especies leñosas es un factor clave en la sobrevivencia, crecimiento y establecimiento exitoso de las especies arbóreas. El uso de sistemas conservacionistas que mitiguen la erosión hídrica permitiría establecer un balance hídrico más favorable, incrementando la infiltración del agua en el suelo y su disponibilidad para el desarrollo de especies agroforestales. El objetivo de este estudio fue evaluar el establecimiento de especies arbóreas mediante diferentes métodos de manejo conservacionista de suelo, determinando el efecto sobre el contenido de humedad del suelo y el crecimiento de las especies. Procedimiento experimental El ensayo fue establecido en otoño del 2007 y ejecutado hasta el 2011, la dimensión de cada parcela fue de 30 x 50 m, en una zona con pendiente de 30%. Las técnicas conservacionistas seleccionadas para el estudio fueron: i) Subsolado con camellón (SC), que consistió en realizar un subsolado a 40cm profundidad y elaborar un camellón donde fueron plantados los árboles; ii) Zanjas de infiltración (ZI), se elaboró una zanja a 20cm profundidad, el ancho de la parte superior fue de 40cm, la tierra que se excavó fuera de la zanja fue acomodada en el borde inferior a 10 cm de la zanja y sobre ella fueron plantados los árboles; y un iii) Testigo sin manejo de suelo (TS), que presentó la plantación de los árboles. Las especies leñosas fueron Alcornoque (Quercus suber), Quillay (Quillaja saponaria) y Tagasaste (Chamaecytisys proliferus). Se evaluó la altura de árboles y diámetro de tronco, así como el contenido de agua en el suelo (0 a 100 cm profundidad). El contenido volumétrico del agua en el perfil se monitoreó con una frecuencia de una a tres semanas las que comenzaron al término de las lluvias de cada temporada hasta el período seco de la temporada 2007, 2008 y 2009. Para este estudio se consideró el contenido volumétrico total desde los 0.2 m hasta 1 m de profundidad. Las mediciones de humedad se realizaron con una sonda de neutrones (TROXLER, 4300 USA), en tubos de acceso de aluminio con un diámetro de 50mm y 1.2 m de largo. Estos tubos fueron instalados en forma vertical a una profundidad de 1.1 m al establecimiento del ensayo. Cuatro tubos fueron instalados en cada réplica, ubicados a una distancia de 0.5 y 1.0 m hacia arriba y hacia abajo de la estructura conservacionista, por lo que en cada tratamiento había 12 tubos de acceso. Los árboles fueron plantados al establecimiento del ensayo, en otoño del 2007. Después de cumplir el primer año, se evaluó en el 2008 y luego en el 2011 la altura y diámetro del tronco. 258 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Las variables en estudio fueron analizadas estadísticamente con un ANOVA (p<0,05). La separación de medias entre tratamientos fue realizada mediante prueba de Duncan, usando el SAS System for Windows v8 (SAS Institute, 2002-2003). Resultados y discusión Las precipitaciones anuales para los años 2007, 2008 y 2009 fueron de 372, 768 y 539 mm, respectivamente, las que se concentraron en los meses de mayo y agosto. El contenido de agua en el suelo en el perfil presentó diferencias significativas dependiendo de la técnica de manejo del suelo, siendo SC superior a ZI y éste a su vez superior a TS. En el año más seco (2007), SC fue superior en un 19% a ZI y un 26% a TS, mientras que en el año más húmedo (2008) SC fue superior en un 16% a ZI y TS. Los resultados además, revelaron diferencias significativas (p>0,05) entre los tratamientos, las especies, así como en su interacción en la altura de árboles y diámetro de tronco. Las evaluaciones al segundo y quinto año de establecimiento de las tres especies agroforestales mostraron un comportamiento distinto en las variables de crecimiento dependiendo del sistema conservacionista utilizado. La especie que demostró un mayor crecimiento fue C. Proliferus, sin embargo, los resultados sugieren que para el establecimiento de esta especie fue más limitante la alta compactación que presentan estos suelos que el contenido de agua en el perfil, al mostrar un mayor crecimiento en el diámetro del tronco en IT>SB>CO (p<0,05). En cambio Q. Suber demostró ser más sensible al contenido de agua en el suelo, al presentar un mayor crecimiento en el tratamiento SB>IT>CO (Fig. 1, Tabla 1). Cada temporada se caracterizó por un periodo de acumulación de agua en el suelo y un periodo de reducción de agua en el perfil debido a la disminución de las precipitaciones. Las técnicas conservacionistas generaron una mayor eficiencia en la captura de las aguas lluvias durante el período invernal permitiendo una mayor disponibilidad del contenido de agua en el suelo durante la estación seca (Castaldi et al., 2012; Makurira et al., 2011; Nichols et al., 2012). Sin embargo, a diferencia de lo esperado, el subsolado con camellón (SB) fue mucho más efectivo que las zanjas de infiltración (IT) en mantener un mayor contenido de agua en el suelo. Esto indicaría que el subsolado promueve una mayor infiltración del agua hacia los horizontes inferiores y una mayor capacidad de conservarla en el perfil durante la estación seca (Martínez et al., 2012; Makurira et al., 2011). Las estructuras de conservación de agua y suelo demostraron un gran potencial para estas condiciones al favorecer un mayor contenido de agua en el perfil en relación al tratamiento testigo (TS), lo que indicaría un menor escurrimiento superficial, siendo este un factor de gran interés para esta zona que se caracteriza por suelos severamente degradados (Martínez et al., 2011; Cao et al., 2007). El tratamiento SC al presentar el mayor contenido de agua en el suelo, permitió una mejor adaptación de los árboles en condiciones de suelo inadecuadas. La ZI es una estructura conservacionista ampliamente utilizada, sin embargo, no es la que más favoreció la infiltración del agua en el suelo a pesar de que el movimiento del suelo al elaborar la estructura favoreció el establecimiento de los árboles. 259 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 70 SC Temporada 2007 ZI 60 TS mm 50 40 30 20 16/10/07 29/10/07 13/11/07 27/11/07 01/01/08 25/02/08 11/10/07 22/10/07 05/11/07 19/11/07 03/12/07 18/01/08 Fecha 70 Temporada 2008 60 mm 50 40 30 20 15/09/08 04/09/08 02/10/08 23/09/08 22/10/08 07/10/08 18/12/08 20/11/08 26/01/09 Fecha 70 Temporada 2009 60 mm 50 40 30 20 16/10/09 29/09/09 10/11/09 29/10/09 07/12/09 24/11/09 Fecha Figura 1. Contenido de agua en el suelo en el perfil (0.2-1 m de profundidad) para las temporadas 2007, 2008 y 2009 (p<0,05). 260 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 SC: Subsolado con camellón; ZI: Zanja de infiltración; TS: Testigo. Tabla 1. Altura y diámetro del tronco para C. Proliferus, Q. Suber y Q. saponaria en las temporadas 2008 y 2011. Especies Tratamientos Altura de árboles C. proliferus Q. suber Q. saponaria 2008 Q. suber Q. saponaria 2011 SC 136 b 184 b ZI 174 a 219 a TS 133 b 150 c SC 56 a 109 a ZI 56 a 91 b TS 54 a 70 c SC 75 a 105 ba ZI 78 a 118 a TS 70 a 92 b Diámetro de tronco C. proliferus Año 2008 2011 SC 171 a 144 b ZI 182 a 223 a TS 111 b 150 b SC 67 a 119 a ZI 57 b 91 b TS 58 b 72 c SC 62 a 109 a ZI 60 a 105 a TS 42 b 83 b Valores seguidos de letras distintas en cada columna indican diferencias significativas, según prueba de Duncan (p<0,05). SC: Subsolado con camellón; ZI: Zanja de infiltración; TS: Testigo. Conclusiones Esta investigación ha confirmado que en suelos severamente degradados y de alta pendiente, el uso de estructuras conservacionista favorece un mayor contenido de agua en el suelo y un mejor establecimiento de las especies arbóreas. Nuestros resultados demuestran que el tagasaste mostró una mejor adaptación 261 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 y un rápido crecimiento en las zanjas de infiltración, en cambio el alcornoque mostró un mayor crecimiento en el subsolado con camellón. El establecimiento y crecimiento del quillay fue más favorable en las estructuras conservacionistas. Finalmente, los resultados demuestran que el subsolado con camellón fue la técnica conservacionista más eficiente en retener agua en el perfil, por ello, debiera ser visto como una técnica conservacionista innovadora en el planeamiento de sistemas agroforestales para zonas degradadas. Bibliografía Cao, S., Chen, L., Xu, C., Liu, Z. 2007. Impact of three soil types on afforestation in China’s Loess Plateau: Growth and survival of six tree species and their effects on soil properties. Landscape and Urban Planning 83: 208–217 Castaldi, F., Chiocchini, U. 2012. Effects of land use changes on badland erosion in clayey drainage basins, Radicofani, Central Italy. Geomorphology, v. 169-170: 98-108. Makurira, H., Savenije, H.H.G., Uhlenbrook, S., Rockström, J., Senzanje, A. 2011. The effect of system innovations on water productivity in subsistence rainfed agricultural systems in semi-arid Tanzania. Agricultural Water Management 98, 1696-1703. Martínez, I., Ovalle, C, Del Pozo, A., Uribe, H., Valderrama, N., Prat, C., Sandoval, M., Fernández, F., Zagal, E. 2011. Influence of conservation tillage and soil water content on crop yield in dryland compacterd Alfisol of central Chile. Chilean Journal of Agricultural Research 71, 615-622. Martínez, I., Prat, C., Ovalle, C. Del Pozo, A., Stolpe, N., Zagal, E. Subsoiling improves conservation tillage in cereal production of severely degraded Alfisols under Mediterranean climate. Geoderma 189190, 10-17. Nichols, M.H., McReynlds, K., Reed, C. 2012. Short-term soil moisture response to low-tech erosion control structures in a semiarid rangeland. Catena 98: 104-109 SAS Institute, 2002–2003. Software. 9.1.3 Service Pack. SAS, Institute, Cary, North Carolina, USA. 262 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 ENLAZANDO CONOCIMIENTO LOCAL Y RASGOS FUNCIONALES DEL COMPONENTE ARBÓREO, BANANOS Y PALMAS EN SISTEMAS AGROFORESTALES CON CACAO Y CAFÉ EN COSTA RICA J.C. Ordonez1, O. Deheuvels2, 3, E. Castro3 Centro Internacional de Investigación en Agroforestería (ICRAF) - América Latina; Centro de Cooperación Internacional en Investigación Agronómica para el Desarrollo (CIRAD) 3 Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). Agroforestería, CATIE 7170, Turrialba 30501, Cartago-Costa Rica [email protected] 1 2 Enunciado del problema Existen varias coincidencias o patrones en el conocimiento local de los agricultores sobre las funciones de los árboles en sistemas agroforestales y los atributos deseables relacionados a estas funciones (Cerdan, 2012; Sinclair and Joshi, 2000). Dado que la información existente tiene un carácter cualitativo, es difícil comparar percepciones y conocimiento de los agricultores en regiones con diferentes condiciones ambientales, diferentes especies arbóreas y sistemas productivos (Heinze, 2013). Métodos basados en la ecología funcional podrían hacer esta comparación posible, utilizando rasgos funcionales. Se ha demostrado que varios rasgos funcionales de las plantas pueden ser utilizados para entender como las plantas responden a factores ambientales y como pueden influenciar procesos ecológicos (Garnier et al., 2004; Lavorel and Garnier, 2002; Violle et al., 2007) independientemente de la identidad botánica de las especies, bioma o tipo de vegetación. Combinar el conocimiento local de los agricultores con herramientas derivadas de ecología funcional puede facilitar los análisis cuantitativos sobre la contribución de los árboles a varios procesos ecológicos así como facilitar la comparación del conocimiento derivado de diferentes personas, sistemas y regiones. Objetivos generales Los objetivos generales del presente trabajo son: Evaluar la correspondencia del conocimiento local de los agricultores sobre los efectos de especies de sombra en la provisión de nutrientes en el suelo, con los valores de rasgos funcionales medidos en campo para las mismas especies arbóreas en sistemas agroforestales con café y cacao Identificar cuáles son los rasgos funcionales de las especies de sombra que mejor reflejan el conocimiento local sobre las funciones ecológicas de regulación de ciclos de nutrientes en sistemas agroforestales de café y cacao Materiales y Métodos Sitios de estudio Las fincas con sistemas agroforestales de café que fueron incluidas en este estudio están ubicadas en los alrededores de Turrialba (09° 47' 14" latitud norte y 83º 34' 03" longitud oeste), en las comunidades de Santa Teresa, Pejibaye, Tres Equis, San Juan Sur, las Colonias de Tuis y la Suiza. Su elevación promedio es de 1000 metros sobre el nivel del mar. Las fincas con sistemas agroforestales de cacao se encuentran en la zona de Talamanca (09°25'40" latitud norte y 83°01'20" longitud oeste) en 3 comunidades: Amubri, Watsi y Namuwoki. Su elevación es de 32 msnm. 263 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Selección de especies Para la selección de especies se hizo una entrevista con los agricultores para conocer cuáles eran las especies más abundantes en sus parcelas de café y cacao. Con la información colectada se elaboró un listado de especies y se escogieron las 20 especies más comunes para cada sistema. Las listas generadas se compararon con información disponible sobre los sistemas de cada zona (Cerdan, 2012; Deheuvels, 2011) para verificar la selección de las especies. Tabla 1. Especies seleccionadas para el estudio Especie Familia Nombre científico SAF café SAF cacao Aguacate Lauraceae Persea americana x x Banano Musaceae Musa sp. x x Cacao Malvaceae Theobroma cacao x Cas Myrtaceae Psidium friedrichsthalianum x Casha Fabaceae Abarema idiopoda Cedro Meliaceae Cedrela odorata Cola de pava Sapindaceae Cupania cinerea Guaba Fabaceae Inga sp. 1 Guabacaite Fabaceae Inga sp. 2 x Guabilla Fabaceae Inga sp. 3 x Guanabana Annonaceae Annona muricata x Guarumo Cecropiaceae Cecropia sp. x Guayaba Myrtaceae Psidium guajava x Guitite Solanaceae Acnistus arborescens x Higuito Moraceae Ficus pertusa x Itabo Agavaceaes Yucca sp. x Jabillo Euphorbiaceae Hura crepitans x Jobos Anacardiaceae Spondias mombin x Laurel Boraginaceae Cordia alliodora x Limon Rutaceae Citrus sp. x Madero negro Fabaceae Gliricidia sepium x Mamon chino Sapindaceae Nephelium lappaceum Manzana de Agua Myrtaceae Syzygium malaccense 264 x x x x x x x x x x x PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Naranja Rutaceae Citrus sp. x Pejibaye Arecaceae Bactris gasipaes x Pilon Phyllanthaceae Hieronyma alchorneoides x Poro Fabaceae Erythrina sp. x Poro Fabaceae Erythrina poeppigiana x Quizarra Lauraceae Ocotea floribunda x Sapote Sapotaceae Pouteria sapota x x Priorización y Entrevistas semi-estructurada En este estudio se caracterizó el conocimiento local de 100 productores de café y cacao con respecto a 3 funciones (regulación de la fertilidad de suelos, control erosión, regulación de microclima), a través de entrevistas semi-estructuradas y un método de priorización (ranking). En este artículo nos concentramos en los resultados sobre fertilidad de suelos. Para el método de priorización se presentó a cada productor, tarjetas con el nombre común y una foto de las 20 especies de sombra más comunes presentes en los cafetales y cacaotales (según el caso). Cada agricultor debería seleccionar al azar 10 especies de las que conociera de la muestra indicada. Se pidió al agricultor que ordenara estas 10 especies de la “mejor” a la “peor” para la regulación de la fertilidad de suelos (producción de abono), control erosión (protección del suelo contra deslaves), y regulación de microclima (que tan fresco es bajo estas especies). Para cada función se registró el orden de las especies y los comentarios del agricultor sobre las razones para el orden dado y cualquier información sobre rasgos funcionales que se mencionara durante la priorización. Mediciones de rasgos funcionales Se midieron 3 rasgos funcionales por su facilidad de medición en campo y el potencial impacto en la regulación de ciclos de nutrientes y producción de biomasa: área específica foliar (g m-2), resistencia de la hoja a la ruptura (N mm-1) y altura de planta (m). Los rasgos se midieron de acuerdo a metodologías establecidas (Cornelissen et al., 2003) Análisis estadístico de resultados Para la priorización se hizo un análisis de comparaciones en pares con el modelo Bradley Terry, que se usa comúnmente en análisis de evaluaciones subjetivas de varios individuos, para obtener respuestas comparativas cualitativas. Este es un modelo linear que estima las posiciones de varias opciones basado en una escala arbitraria, con una opción (especie) de referencia contra la que las demás opciones son comparadas. La distancia entre sus posiciones permite calcular la probabilidad de una opción priorizada sobre otra. Los datos fueron analizados con un paquete de R “RankAnalysis” elaborado por el Centro De Servicios Estadísticos De La Universidad De Reading y el Centro Internacional para la Investigación Agroforestal (ICRAF). La correspondencia de los resultados del análisis de priorización con las medidas de rasgos funcionales se hizo con análisis de correlación de Pearson. Resultados Conocimiento local y priorización de especies por sus efectos en la fertilidad del suelo 265 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 En sistemas de café 43 de los 50 agricultores entrevistados indicaron un total de 14 características de los árboles que determinan efectos positivos sobre la fertilidad del suelo, siendo las más mencionadas la producción de muchas hojas (n=36) y la rápida descomposición de las hojas (n=33), y solo unos pocos mencionaron descomposición lenta (n=4), hojas suaves (n=3) y copa extensa (n=3). En los sistemas de cacao 45 de los 50 agricultores entrevistados indicaron un total de 15 criterios que incluían características de los árboles en si (n=13), e impactos que los agricultores observan en el suelo bajo los árboles (n=2). Las características de árboles más mencionadas fueron la producción de muchas hojas (n=31), la rápida descomposición de las hojas (n=13) y el aporte de la descomposición de frutas (n=7); mientras que en cuanto a impactos en el suelo se mencionó como muy importante el observar tierra de color negro (n=16) y suelo húmedo (n=11). El ejercicio de priorización de especies por sus efectos en la fertilidad del suelo en sistemas de café, muestran al Poro (Erythrina poeppigiana) como el mejor árbol para la fertilidad, seguido especies de banano (Musa sp.); mientras que el pejibaye (Bactris gasipaes) y el itabo (Yucca sp.) como los que menos contribuyen a la fertilidad del suelo. El resto de árboles están entre estos extremos y fueron ordenados pero las diferencias son menos claras. En el caso del cacao la priorización de árboles fue menos definida en general (mayores errores estándar, indicando más variabilidad en las respuestas). Se identificó como la mejor especie para la fertilidad del suelo al poro (Erythrina sp.), seguida de guaba (Inga sp. 1), guabilla (Inga sp. 3) y madero negro (Gliricidia sepium). La única definición clara fue para el pejibaye (Bactris gasipaes) que se colocó como la especie que menos contribuye a la fertilidad del suelo (figura 1). Rasgos funcionales de árboles, bananos y palmas en sistemas agroforestales de café y cacao Los rasgos funcionales fueron escogidos por su potencial impacto en la regulación de ciclos de nutrientes y producción de biomasa: área específica foliar (2.45 - 45.8 g m-2), resistencia de la hoja a la ruptura (0.5 – 6.5 N mm-1) y altura de planta (2.3 – 18.5 m). Los diferentes rasgos mostraron un rango de variabilidad dentro de los limites esperados para estos rasgos funcionales(Cornelissen et al., 2003). A) B) Figura 1. Priorización de especies de sombra con respecto a sus impactos en la fertilidad del suelo en sistemas agroforestales de café (A) y cacao (B). 266 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Correspondencia de priorización de especies por los agricultores y rasgos funcionales medidos en campo La relación entre la priorización de los agricultores y los rasgos funcionales fue diferente para cada sistema agroforestal y para cada rasgo funcional. En general altura no estuvo relacionada a la priorización de especies en ningún sistema. El área específica foliar estaba positivamente relacionada a la priorización de especies en sistemas agroforestales con café (r=0.74, P<0.001), pero no tenía ninguna relación a la priorización de especies en sistemas agroforestales con cacao (r=0.06, P=0.81). El quiebre mostro una relación negativa a la priorización de especies en los dos sistemas agroforestales (café: r= 0.55, P= 0.017; cacao: r= -0.44, P= 0.052) Discusión Conocimiento local y priorización de especies Los principales criterios que los agricultores utilizan para evaluar especies de sombra por sus efectos en la fertilidad del suelo son muy similares en sistemas de café y cacao, siendo estos principalmente la alta producción de hojarasca de rápida descomposición. Lo cual indica que los agricultores prefieren plantas que favorecen un ciclo rápido de nutrientes en grandes cantidades (el extremo rápido del espectro de la económica foliar)(Westoby et al., 2002; Wright et al., 2004). Sin embargo las respuestas dadas también muestran algunas diferencias interesantes, como por ejemplo que los agricultores en Talamanca que producen cacao, también mencionaron muy frecuentemente los impactos de las especies en el suelo (tierra negra, tierra húmeda) antes que los atributos de las especies en sí, y fueron los únicos que mencionaron a las frutas y flores como componente importante de biomasa para abonar el suelo. Esto podría explicarse por diferencias culturales y de manejo entre cacao y café. Las comunidades indígenas de Talamanca tienen bajos niveles de intensificación y tecnificación, muchos de sus cacaotales mantienen varias especies arbóreas, bananos y palmas en plena producción, mientras que los sistemas de café de Turrialba se maneja mucho más la sombra con podas por lo que la producción de fruta puede ser mucho menor o en algunos casos inexistente. Muy pocos agricultores mencionaron atributos que directamente reflejan los rasgos funcionales que fueron medidos en campo, esto es lógico ya que es más probable que los agricultores evalúen las especies por características más integrales (producción de biomasa). a) sistemas agroforestales con cacao b) Sistemas agroforestales con café Figura 2. Diagrama bi-variado de correspondencia de priorización de especies por los agricultores y rasgos funcionales medidos en campo 267 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 En cuanto al ejercicio de priorización, fue posible ordenar las diferentes especies de árboles y mostrar la diferencia que tienen unas especies con otras de acuerdo a la percepción de los agricultores. El patrón común que emerge de este análisis es que los agricultores pueden distinguir claramente unas pocas especies en los extremos (las mejores y las peores), pero no es posible tener una clara distinción para las especies entre los extremos. En el caso de Talamanca se muestra una menor división de las especies en general, lo que se refleja en errores estándar mayores. Este patrón refleja que los agricultores tienen un conocimiento o una percepción más concreta acerca de los impactos de unas pocas especies con efectos fuera de lo normal y un conocimiento bajo de otras especies. En el caso de Talamanca, los resultados parecen reflejar el menor manejo que se da a la sombra (y por ende una menor experiencia con las especies en campo). Es importante considerar que el ejercicio de priorización da resultados relativos y por depender mucho de la persona que responde y de las especies que entraron en el ejercicio de priorización, por ejemplo una especie puede ser puesta en un lugar muy bajo en su contribución a la fertilidad en comparación con otras especies que fueron priorizadas, pero puede ser que los agricultores todavía consideran a esta una buena especie. Este tipo de problemas pueden ser aminorados al incrementar el tamaño de la muestra y colectando más información específica a las especies durante las entrevistas de priorización y entrevistas sobre conocimiento local. Conocimiento local vs. Rasgos funcionales La comparación de las estimaciones obtenidas con la priorización y los rasgos funcionales, en el caso de café muestran que existe correspondencia entre las preferencias de los agricultores (que buscan una alta producción de biomasa de rápida descomposición) y la teoría de la economía foliar. Esto es las especies que los agricultores evaluaron como las mejores contribuyentes a la fertilidad del suelo son aquellas que tienen una alta área específica foliar y baja resistencia a la ruptura. Estos resultados van de acuerdo a la teoría de la economía foliar que muestra la existencia de un eje de variación de rasgos funcionales foliares en las plantas. En un extremo se encuentran plantas que tienen hojas delgadas, delicadas, con altos contenidos de nutrientes y un rápido crecimiento pero por cortos plazos; mientras que en el otro extremo se encuentran plantas con una estrategia conservadora de características opuestas y lento crecimiento (Wright et al., 2004). Las plantas con la estrategia de rápido crecimiento crecen mejor en sitios de alta fertilidad (Ordoñez et al., 2009) pero también producen hojarasca con altos contenidos de nutrientes con una rápida descomposición, dando una retroalimentación positiva al suelo para mantenerlo con una alta fertilidad (Aerts and Chapin, 2000; Berendse, 1994). Entonces es claro que la preferencia de los agricultores refleja el objetivo de mantener un ecosistema con un rápido ciclo de nutrientes en el suelo en grandes cantidades, necesario para mantener la producción agrícola. Aunque esto no limita la presencia de otras especies como el pejibaye que seguramente se mantiene por sus beneficios alimenticios. En el caso del cacao los resultados solo concuerdan con la teoría para el caso la resistencia a la ruptura mientras que no hay relación con área específica foliar. En los sistemas de cacao se encontraron especies con áreas específicas foliares sumamente altas (más allá de los valores que se encontró en café). Esto puede deberse a tres posibilidades 1) que existieron errores de medición 2) que estas especies con alta área específica foliar tienen otras características que pueden reducir su descomposición (por ejemplo alto contenido de compuestos secundarios como fenoles) o 3) que los agricultores no aprecian a estas especies como las mejores para la fertilidad de los suelos porque no las conocen suficiente. Dada la relativa buena correspondencia con los datos de quiebre, parece ser que la primera opción es posible. En el caso de Talamanca debido a su aislamiento y falta de infraestructura, no fue posible realizar el secado de las hojas inmediatamente después de la colección, esto puede tener 268 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 impactos en el peso de las hojas debido a pérdidas de exudados. Sin embargo no se debe descartar que existen otras características importantes, que no han sido medidas, por ejemplo también en Talamanca la guabilla es una de las especies de mayor preferencia pero no tiene una baja resistencia al quiebre, cabe mencionar que los productores la prefieren debido a que especie produce un alta cantidad de hojas y es fijadora de Nitrógeno. Con respecto a la tercera posibilidad, cabe mencionar que los agricultores no dieron referencias a él porque algunas de estas especies con alta área específica foliar no fueron priorizadas como las mejores. En general parece que la resistencia al quiebre puede ser un rasgo funcional que podría utilizarse para comparar especies de diferentes sistemas con respecto a su contribución a la fertilidad del suelo, esto por la relativa correspondencia que se obtuvo con las apreciaciones de los agricultores, la base de conocimiento ecológico y su facilidad para ser medido en campo. Sin embrago es necesario explorar tanto con el quiebre como con área específica foliar cuales son las fuentes de las inconsistencias encontradas, para refinar los métodos de conocimiento local así como para identificar si es necesario evaluar otros rasgos funcionales en campo. Referencias Aerts R., Chapin F.S. (2000) The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Advances in Ecological Research 30:1-67. Berendse F. (1994) Litter decomposability - a neglected component of plant fitness. The Journal of Ecology 82:187-190. Cerdan C.R. (2012) Local knowledge of impacts of tree cover on ecosystem services in smallholder coffee production systems. Agricultural Systems 110:119-130. Cornelissen J.H.C., Lavorel S., Garnier E., Diaz S., Buchmann N., Gurvich D.E., Reich P.B., ter Steege H., Morgan H.D., van der Heijden M.G.A., Pausas J.G., Poorter H. (2003) A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany 51:335-380. Deheuvels O. (2011) Compromis entre productivité et biodiversité sur un gradient d’intensité de gestion de systèmes agroforestiers à base de cacaoyers de Talamanca, Costa Rica, Ecole Doctorale Systèmes Intégrés en Biologie, Agronomie, Géosciences, Hydrosciences, Environnement, SUPAGRO Montpellier, Montpellier. Garnier E., Cortez J., Billes B., Navas M.L., Roumet C., Debussche M., Laurent G., A B., D A., A B., Neill C., Toussaint J.P. (2004) Plant functional markers capture ecosystem properties during secondary succession. Ecology 85:2630-2637. Heinze A. (2013) Farmer's local ecological knowledge on trees: coffee agroforestry systems in two Central American sites, School of Environment, Natural Resources and Geography, Bangor University. Lavorel S., Garnier E. (2002) Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Functional Ecology 16:545-556. Ordoñez J.C., van Bodegom P.M., Witte J.P.M., Wright I.J., Reich P.B., Aerts R. (2009) A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Global Ecology and Biogeography 18:137-149. 269 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Sinclair F.L., Joshi L. (2000) Taking local knowledge about trees seriously, in: A. Lawrence (Ed.), Forestry Forest Users and Research: New ways of learning, European Tropical Research Network (ETFRN), Wageningen Netherlands. pp. 45-58. Violle C., Navas M.L., Vile D., Kazakou E., Fortunel C., Hummel I., Garnier E. (2007) Let the concept of trait be functional! Oikos 116:882-892. DOI: 10.1111/j.2007.0030-1299.15559.x. Westoby M., Falster D.S., Moles A.T., Vesk P.A., Wright I.J. (2002) Plant ecological strategies: Some leading dimensions of variation between species. Annual Review of Ecology and Systematics 33:125-159. Wright I.J., Reich P.B., Westoby M., Ackerly D.D., Baruch Z., Bongers F., Cavender-Bares J., Chapin T., Cornelissen J.H.C., Diemer J., Flexas J., Garnier E., Groom P.K., Gulias J., Hikosaka K., Lamont B.B., Lee T., Lee W., Lusk C.H., Midgley J.J., Navas M.L., Niinemets U., Oleksyn J., Osada N., Poorter H., Poot P., Prior L., Pyankov V.I., Roumet C., Thomas S.C., Tjoelker M.G., Veneklaas E.J., Villar R. (2004) The worldwide leaf economics spectrum. Nature 428:821-827. 270 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 RUNA GUAYUSA – DESARROLLO DE UN SISTEMA DE CULTIVO AGROFORESTAL DE Ilex guayusa Loes. Juan F. Dueñas-Serrano1, Eliot Logan-Hines1, Maureen Stimola1, Florencia Montagnini2, Alexandra Humanate1, Nigel Melican3. 1 Grupo Runa (Fundación Runa y Runatarpuna Exportadora S.A.), Archidona, Ecuador. de Ciencias Forestales y Medio Ambientales de la Universidad de Yale, Northford, CT, Estados Unidos de América. 3Teacraft Technical Services, Bedford, Reino Unido. [email protected] 2Escuela Resumen El grupo Runa trabaja para crear una nueva cadena agroforestal de valor enfocada en la guayusa (Ilex guayusa Loes). La guayusa es un árbol nativo de la Amazonía cuyas hojas contienen altas concentraciones de cafeína y es apto para cultivo en asociación a otras especies. La utilidad de la guayusa ha sido reconocida por varios grupos de nativos amazónicos, en particular por la nacionalidad Kichwa de la alta Amazonía ecuatoriana. Aunque el uso ancestral ha permitido conocer algunas de las características biológicas y agronómicas de ésta especie, el conocimiento disponible es insuficiente para asistir en la transición de la guayusa hacia un cultivo de exportación. Debido a ello, Runa está desarrollando un sistema de trabajo con 2500 pequeños productores que busca re-valorizar el conocimiento ancestral, mientras se genera nuevo conocimiento. En el 2013 Runa, a través del apoyo del Programa Manejo Forestal Sostenible en la Región Andina del Instituto Interamericano de Cooperación para la Agricultura (IICA) y el Ministerio de Asuntos Exteriores de Finlandia, ha diseñado un amplio programa de investigación científica y participativa. Ésta presentación resumirá el alcance y enfoque de este programa de investigación y los avances obtenidos en lo que va de este año. Específicamente se tocarán estos aspectos: (1) compilación y revalorización del conocimiento ancestral sobre el cultivo de Ilex guayusa Loes; (2) revisión de los ejes del programa de investigaciones y avances; y, 3) recomendaciones para el futuro. Palabras Clave: Ilex, sistemas agroforestales, investigación participativa, conocimiento ancestral Introducción Durante miles de años la Amazonía ha sido habitada por pueblos indígenas ancestrales que han desarrollado sistemas agrícolas complejos como medio de subsistencia. Sin embargo, entre académicos y la sociedad en general existe una concepción de la Amazonía que considera a ésta como un ecosistema prístino capaz de sostener solamente a pequeños grupos humanos de cazadores y recolectores nómadas. Por el contrario, en este artículo sostenemos que la Amazonía posee un historial extenso en cuanto al manejo agrícola y que en la actualidad podemos aprovechar este rico legado. Numerosas especies amazónicas han sido utilizadas por las poblaciones indígenas durante milenios, como parte de sus sistemas de subsistencia. Entre estas especies la guayusa, Ilex guayusa Loes., una planta nativa del noroeste de la amazonía, ha sido tradicionalmente cultivada y usada por diferentes nacionalidades indígenas para preparar una bebida ritual, que actúa como estimulante y emético (Lewis et al., 1991; Schultes, 1972). Las 271 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 propiedades medicinales y estimulantes de la guayusa la están haciendo una bebida popular en mercados de USA y Canadá. Debido a ello el grupo Runa busca promover la intensificación del cultivo en el marco de un sistema agroforestal multi-estrato, basado en un programa de investigación científica que cubre aspectos del conocimiento ancestral de la especie, así como su agronomía, genética, ecología y fito-química. Este artículo busca hacer un breve recuento de lo que sabemos sobre la guayusa, y una revisión del ámbito y avances obtenidos hasta el momento en el programa científico que RUNA está llevando a cabo. Esperamos que esto nos ayude a entablar un diálogo con instituciones de investigación públicas y privadas, para atraer su atención hacia este cultivo promisorio. Así mismo, esperamos que este diálogo nos ayude a elaborar una agenda de prioridades de investigación para esta especie en el futuro. ¿Qué es la guayusa? – aspectos botánicos y ecológicos Ilex es el único género extante de la familia Aquifoliaceae, una familia con un pasado filogenético oscuro (Judd et al., 1999). El género Ilex es cosmopolita y se compone de alrededor de 600 especies, las cuales típicamente se distribuyen en sitios con altos índices de precipitación y humedad (Loizeau et al., 2005). Esto hace que sean especialmente abundantes en las áreas montañosas de los trópicos. En el neotrópico se encuentran alrededor de 300 especies (Loizeau and Barreira, 2007) y en Ecuador están reconocidas alrededor de 32 especies (Loizeau and Barriera, 1999). La guayusa es una especie arbórea que se distribuye en el extremo nor-occidental de la Amazonía Sud Americana, desde el norte Peruano hasta el sur de Colombia entre los 200 a 2500 msnm (Rios et al., 2007). En el Ecuador se ha registrado en las provincias de Sucumbíos, Napo, Orellana, Pastaza, Morona Santiago, Zamora Chinchipe, Pichincha, Loja y Tungurahua. Los orígenes evolutivos de la guayusa son igualmente oscuros, de hecho Loesener describió la especie en ausencia del espécimen tipo (Loizeau and Barriera, 1999). La morfología floral y la anatomía de la madera están bien conservadas entre especies del género Ilex, mientras que la variación intra-especie en cuanto a la morfología de las hojas es alta. Estas observaciones indican una alta propensión a la hibridización (Brizicky, 1964; Manen, 2004). La evidencia morfológica está apoyada por la evidencia genética, ya que se ha observado inconsistencias entre las filogenias inferidas a partir de DNA proveniente de plástidos versus las producidas a partir de DNA del núcleo celular (Manen et al., 2010). Esto ha conducido a la conclusión de que una amplia revisión taxonómica del género muy probablemente reduciría el número de especies actualmente reconocidas (Loizeau et al., 2005). Aunque la guayusa fue descrita formalmente como especie en 1901, se ha publicado poco sobre su historia natural y ecología. Sus flores fueron descritas casi 80 años después de la descripción de la especie (Shemluck, 1979). La misma publicación de Shemluck (1979) pone de manifiesto la dificultad de encontrar material fértil. Quizá debido a ello, las plantas de guayusa se cultivan exclusivamente mediante estacas leñosas o semi-leñosas de tamaños variables tomados de una planta madura, es decir, aproximadamente mayor a 4 años. Su fruto es una drupa, verde o roja, y crece hasta alcanzar un centímetro de diámetro. Las plantas son dioicas, la flor de guayusa posee una corola blanco-verdosa con pétalos obtusos de igual número de estambres y pétalos (4), anteras alargadas, y un ovario sub-sésil global que contiene de 4 a 6 cavidades (García Barriga, 1992). Los árboles de guayusa pueden alcanzar una altura media de 10 metros y un diámetro promedio a la altura del pecho de 50 a 80 centímetros. Típicamente, las plantas cultivadas de guayusa están 272 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 constituidas por una multitud de troncos delgados y flexibles que se nacen desde su base. Dicho esto, se ha documentado que ocasionalmente los árboles de guayusa pueden alcanzar alturas de más de 25 metros (Figura 1). Figura 1. Imagen de un árbol antiguo de guayusa junto a una planta frecuentemente cultivada y una juvenil en la provincia de Napo, Ecuador. A la infusión de guayusa se le atribuye varias funciones medicinales como tónico, diurético, anti-gripal, analgésico y promotor del líbido (de la Torre et al., 2008) Se ha detectado que las hojas de guayusa contienen altas concentraciones de alcaloides (cafeína y teobromina), fenoles, taninos, azúcares reductores, esteroides, terpenos, compuestos lactónicos, flavonoides y quinonas (Radice and Vidari, 2005). La presencia de cafeína (8,13 mg por ml) y otros químicos promotores de salud no es extraño dentro del género, ya que existen al menos tres especies más con estas características, de las cuales la más conocida es la Yerba Mate (Ilex paraguariensis) (Hao et al., 2013). Es por ello que la guayusa se encuentra listada como una de las especies más promisorias de la Amazonía (Russo, 1992). Conocimiento ancestral de la guayusa – lecciones del pasado Se ha reportado el uso de guayusa como estimulante y medicamento en al menos seis grupos indígenas del oriente ecuatoriano: Kichwa, Cofán, Secoya, Záparo, Shuar y Achuar (de la Torre et al., 2008; Kvist and Holm-Nielsen, 1987) y al menos tres grupos de otras regiones: Kichwa de la sierra, Mestizo y Tsa’chi (de la Torre et al., 2008) No está claro desde cuándo los grupos indígenas de la amazonía empezaron la domesticación de guayusa, pero está claro que se conoce desde al menos 1500 años. Existen crónicas que hacen referencia al uso de la guayusa por parte de los nativos Amazónicos que datan del siglo XVII. Muchas de estas curiosas referencias fueron documentadas por misioneros y científicos europeos (Patiño, 1968; Schultes, 1979). Por ejemplo, se encontró un atado de hojas de guayusa en una tumba tiahunacoide en el altiplano Boliviano que data del año 500 DC (Schultes, 1972). La guayusa ha sido reconocida por sus propiedades terapéuticas y simbólicas, especialmente entre los Kichwas (Innerhofer and Bernhardt, 2011) y los Achuar (Lewis et al., 1991). La gente Kichwa cosecha hojas frescas de guayusa para hervirlas por horas a la mañana siguiente. Entre los Kichwa, beber guayusa en las mañanas ayuda a mejorar la convivencia y relaciones afectivas dentro del grupo familiar. Se aprovecha el ritual para aconsejar a los miembros jóvenes de la familia, planear el día o simplemente 273 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 intercambiar experiencias. También se considera que la guayusa “proteje” al cazador durante su jornada nocturna y repele ciertos peligros de la selva como serpientes y mosquitos. Entre los Achuar, es una bebida purificadora que debe tomarse en la mañana para limpiar el cuerpo de impurezas mediante el vómito. El programa científico de RUNA – un nuevo contexto A partir de febrero de 2010, la Fundación Runa y su socio exportador, la empresa Runatarpuna Exportadora, S.A. empezaron a reproducir guayusa a través de viveros comunitarios. Entre 2010 y 2012 alrededor de 150.000 plantas de guayusa fueron sembradas entre aproximadamente 750 familias agricultoras en la provincia de Napo. La propagación de la planta fue enteramente vegetativa. Por lo general la guayusa fue plantada en hileras de 3 x 3 m con 200 plantas asignadas a cada agricultor para un área de una hectárea en forma de parcela. Esta etapa de siembra intensificada dio el inicio al segundo periodo de domesticación de guayusa. La segunda etapa de domesticación posee un nuevo conjunto de condiciones basadas en el contexto de la comercialización de la planta en lugar de su uso doméstico. Por ejemplo, el nuevo modelo de comercialización exige la siembra de guayusa a mayores densidades (i.e. entre 50 a 1000 plantas ha-1). Esto puede afectar negativamente la diversidad de los agro-ecosistemas representados en las chacras tradicionales provocando el estancamiento de la productividad a largo plazo, incrementando la vulnerabilidad a enfermedades y plagas y comprometiendo la provisión de servicios del ecosistema (Tscharntke et al., 2011). La domesticación de un cultivo va más allá de la esfera ambiental ya que se relaciona con el patrimonio cultural, estilo de vida y estructura social de los agricultores. Por esta razón el enfoque de las investigaciones de Runa da importancia a la dinámica social, cultural y económica del cultivo; sin descuidar la necesidad de información sobre las propiedades genéticas, fitoquímicas, biológicas, agronómicas y ecológicas de la especie. 2013 un arduo año de trabajo – avances del programa científico Variabilidad Genética: Desde Agosto del 2012, en conjunto con el laboratorio de biotecnología vegetal de la USFQ13, se colectaron 150 muestras de guayusa en seis provincias de la Amazonía y una de la Sierra sur (Anexo 1). Se ha extraído ADN de todas las muestras, el cual está siendo amplificando utilizando 12 marcadores moleculares que flanquean microsatelites (inter simple sequence repeat) previamente caracterizadas para el género Ilex. Hasta el momento uno de los locus ha mostrado un polimorfismo moderado exhibiendo seis alelos (Figura 2). Fito-química y procesamiento: Muestras de hojas maduras de las mismas 150 plantas colectadas para el estudio genético fueron enviadas al laboratorio de química analítica de la USFQ. Extractos acuosos de Guayusa están siendo sometidos a HPLC14 para determinar el contenido de cafeína, teobromina, ácido cafeínico, ácido glutámico y L-teanina. También se está midiendo la capacidad antioxidante de los extractos mediante titulación con el método de radicales libres DPPH15. El grupo Runa recientemente equipó un laboratorio de control de calidad e innovación en el procesamiento. Este laboratorio está investigando la mejor manera de producir guayusa para conservar sus propiedades medicinales 13 Universidad San Francisco de Quito Cromatografía líquida de alta presión 15 2,2 difenil -1-picrylhydrazyl 14 274 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figura 2. Gel de poliacrilamida al 1.75% mostrando el polimorfismo alelico del marcador p824 Reserva genética: Estacas de cada espécimen colectado para estos análisis se retuvieron en el invernadero de RUNA, con miras a establecer un jardín clonal, una vez que los resultados de estos estudios sean conocidos. Parcelas experimental multifactorial: A finales del 2012 y durante el 2013, el grupo Runa ha implementado varias parcelas multifactoriales en la finca de su propiedad en Archidona. Para ello se ha contado con la asesoría internacional de Teacraft Technical Services. En un área de 7500 m2 se establecieron tres bloques (replicas) de 25 parcelas experimentales (10 m2). Cada parcela contiene 49 plantas de guayusa a una densidad de 1,25 x 1,25 m (n = 3675 plantas). Estas parcelas tienen el objetivo principal de estudiar los efectos del estado nutricional del suelo, técnicas de poda formativa, cosecha y frecuencia de las dos anteriores en el crecimiento y estado nutricional de la planta y hojas de guayusa. A partir de Junio de 2013 se empezó a aplicar un tratamiento de poda formativa de fuste principal en tres niveles de severidad: cada 5 cm, cada 10 cm y cada 20 cm. Aprovechando estos ensayos se está midiendo el crecimiento y diámetro del tallo principal. Parcelas experimentales agroforestales (SAF): Runa desea promover el aumento de siembra de guayusa mediante el diseño de sistemas agroforestales multi-estrato altamente productivos. El establecimiento de estas parcelas empezó en el mes de agosto de 2013. La guayusa va a ser sembrada en hileras de 1,8 m x 2 m. Para el control de malezas se va a sembrar yerba luisa (Cymbopogon citratus) cada 6 hileras de guayusa. Además para el estrato herbáceo se utilizará maní forrajero (Arachis pintoi) como cobertura en todas las secciones entre parcelas. Para el estrato arbóreo se piensa colocar, según la disponibilidad de plantines en viveros locales, especies maderables tales como Chuncho (Cedrelinga cataeniformis), Batea (Cabralea canjerana), Pachaco (Schizolobium amazonicum), y especies fijadoras de nitrógeno como la Guaba (Inga spp.). Agro Ecología de guayusa en la chacra Kichwa: Se han realizado 19 visitas a chacras tradicionales Kichwa. Las zonas visitadas están estratificadas por altura (0-600m, 600-1200 m) y tipo de suelo (aluvial o no aluvial). En cada chacra se ha muestreado el suelo adyacente a una planta madura de guayusa (> 6 años de edad estimada) cultivada de la forma tradicional. El objetivo es relacionar las características biofísicas del suelo con la productividad de árboles maduros de guayusa. La productividad estará 275 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 determinada por el peso el promedio de la cosecha de ese individuo en ese año. Quince muestras de suelo han sido analizadas en el laboratorio de Agrocalidad en Quito. Un vistazo preliminar a estos datos indica que el contenido de nutrientes es constante para la mayoría de los sitios visitados (Tabla 1). El pH promedio, incluyendo todas alturas y todos tipos de suelos, es 5,4 (ácido). Las chacras cercanas a ríos tienen un nivel de fosforo medio (~11 ppm) mientras que las demás tienen un nivel bajo (~7 ppm). Hemos encontrado que las plantas más frecuentemente asociadas a guayusa en un rango de 10 m son cacao, café, y plátano. Tabla 1. Resumen de parámetros químicos encontrados en el suelo de las chacras Kichwas. Solo se presentan resultados de macro-elementos. Las columnas resaltadas pertenecen a chacras cercanas a ríos. Sitio pH Materia Orgánica (%) Nitrógeno total (%) Fósforo (ppm) Potasio (cmol/Kg) Flor de Bosque 5.15 5.01 0.25 <3.5 0.28 Santo Domingo 5.71 4.9 0.24 3.9 0.39 9 de Junio 5.06 4.42 0.22 <3.5 0.2 Ahuano (1) 5.02 5.49 0.27 <3.5 0.32 Ahuano (2) 5.34 5.82 0.29 4.9 0.36 Dos Piedras 5.72 4.47 0.22 <3.5 0.21 Union Venecia 6.07 5.24 0.26 6.1 0.16 Alto Pusuno 5.27 5.44 0.27 5.6 0.16 Aguayaku 4.96 5.21 0.26 5.3 0.12 Santa Rita 01 5.25 3.05 0.15 <3.5 0.31 Santa Rita 02 5 1.95 0.1 <3.5 0.3 Pachakutik 5.22 2.61 0.13 <3.5 0.2 10 de Agosto 5.21 3.35 0.17 <3.5 0.26 Santa Rosa 5.42 0.99 0.05 14.4 0.16 San Cristobal 5.48 1.36 0.07 13.2 0.21 Propagación: En el vivero de producción perteneciente a RUNA se ha empezado a producir guayusa con un protocolo estandarizado de colección y siembra. La meta es alcanzar los 40.000 plantines al año. Investigaciones preliminares en estas instalaciones arrojaron indicios interesantes. En primer lugar, las estacas extraídas de plantas madre ubicadas en sitios de barbecho tienen mayor porcentaje de sobrevivencia que estacas extraídas de plantas bajo otros regímenes de uso de suelo. Por otro lado, estacas de diámetros < 1 cm tienen menor tasa de supervivencia en las condiciones impuestas por los viveros comunitarios (de Vlieger, 2011). Un segundo estudio determinó que aunque dos aditivos de enraizamiento comerciales, Raizplant 50016 y Rootmost17 aumentan el porcentaje de enraizamiento de estacas leñosas (73,19—83,33%, 20—25 cm de longitud y 1—3 cm de diámetro), ninguno de estos tratamientos motivó aumentos significativos en relación a los controles (Ríos-Granizo, 2011). Se está empezando a colectar y caracterizar morfológicamente frutos y semillas de guayusa, los cuales serán sometidos a ensayos de germinación. Finalmente, se está trabajando en el diseño de experimentos de propagación vegetativa que nos ayuden a reducir el tamaño del material requerido para propagación y entender las características optimas del ambiente de la planta madre como el de propagación. 16 17 Raíz Plant 500, N, P, K, S, B, ácido húmico, ácido fúlvico, fitohormonas no identificadas (500 ppm). Dosis: 300 ml/20 l de H2O Rootmost, Algas, N, P, K, citoquininas (80 ppm), gibelerinas (10 ppm), auxinas (1000 ppm). Dosis 400 ml/20 l de H2O 276 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Investigación participativa: Se ha empezado un programa de investigación participativa y transferencia de conocimientos mediante el involucramiento de agricultores Kichwa y los técnicos de campo del grupo Runa. Se han conducido dos talleres para definir temas de investigación y se ha realizado visitas de seguimiento a las propiedades de los agricultores involucrados. Gracias a esta apertura, hemos aprendido que la guayusa se reproduce mejor mediante estacas presentes en la parte basal de los árboles y que sus frutos son consumidos y dispersados por aves. Hemos aprendido que la reproducción y primeros años de crecimiento requieren de bajos niveles de irradiación y alta humedad. Junto con los agricultores estamos observando y reflexionando sobre los pasos adecuados para establecer una chacra de guayusa, conservar el suelo, la densidad óptima de siembra, el control de malezas, fertilización, métodos de poda; y sobre todo, los retos que impone el manejo orgánico de un cultivo perenne. Literatura citada: Brizicky, G.K., 1964. The genera of Celastrales in the southeastern United States. Journal of the Arnold Arboretum 45, 206–234. De la Torre, L., Navarrete, H., Muriel M., P., Macía, M.J., Balsev, H., 2008. Aquifoliaceae, in: de la Torre, L., Navarrete, H., Muriel M., P., Macía, M.J., Balsev, H. (Eds.), Enciclopedia de Las Plantas Útiles Del Ecuador. Herbario QCA & Herbario AUU, Quito & Aarhus, pp. 177–178. De Vlieger, R., 2011. Amazon guayusa : live the stimulating life. Van Hall Larenstein University of Applied Sciences. García Barriga, H., 1992. Flora Medicinal de Colombia, Tomo 2. ed. Editores Tercer Mundo, Bogotá. Hao, D., Gu, X., Xiao, P., Liang, Z., Xu, L., Peng, Y., 2013. Research progress in the phytochemistry and biology of Ilex pharmaceutical resources. Acta Pharmaceutica Sinica B 3, 8–19. Innerhofer, S., Bernhardt, K.-G., 2011. Ethnobotanic garden design in the Ecuadorian Amazon. Biodiversity and Conservation 20, 429–439. Judd, W.S., Campbell, C.S., Kellog, E.A., Stevens, P.F., 1999. Plant Systematics: A phylogenetic approach. Sinauer Associates Inc., Sunderland. Kvist, L.P., Holm-Nielsen, L.B., 1987. Ethnobotanical aspects of lowland Ecuador. Opera Botanica 92, 83– 107. Lewis, W., Kennelly, E., Bass, G., Wedner, H., Elvin-Lewis, M., Fast, W., 1991. Ritualistic Use of the Holly Ilex guayusa by Amazonían Jivaro indians. Journal of Ethnopharmacology 25–30. Loizeau, P.A., Barreira, G., 2007. Aquifoliaceae Neotropicales: Descriptions, Illustrations, Identification, et Recherche d’Information. [WWW Document]. 1st march 2007. URL http://www.villege.ch/cjb/bd/aqf/species_e/guayus_c.htm 277 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Loizeau, P.-A., Barreira, G., Manen, J.-F., Broennimann, O., 2005. Towards an understanding of the distribution of Ilex L . ( Aquifoliaceae ) on a World-wide scale. Biologiske Skrifter. Loizeau, P.-A., Barriera, G., 1999. Aquifoliaceae, in: Jørgensen, P.M., León-Yánez, S. (Eds.), Catalogue of the Vascular Plants of Ecuador, Monographs in Systematic Botany from the Missouri Botanical Garden. Missouri Botanical Garden, St. Louis, pp. 225–227. Manen, J.-F., 2004. Are both sympatric species Ilex perado and Ilex canariensis secretly hybridizing? Indication from nuclear markers collected in Tenerife. BMC evolutionary biology 4, 46. Manen, J.-F., Barriera, G., Loizeau, P.-A., Naciri, Y., 2010. The history of extant Ilex species (Aquifoliaceae): evidence of hybridization within a Miocene radiation. Molecular phylogenetics and evolution 57, 961–77. Patiño, V.M., 1968. Guayusa, a neglected stimulant from the eastern Andean foothills. Economic Botany 22, 310–316. Radice, M., Vidari, G., 2005. Caracterización fitoquímica de la especie Ilex guayusa Loes . y elaboración de un prototipo de fitofármaco de interés comercial. La Granja 3–11. Rios, M., Koziol, M.J., Borgtoft, H.P., Granda, G., 2007. Plantas útiles del Ecuador: Aplicaciones, retos y perspectivas, Primera. ed. Abya-Yala, Quito. Ríos-Granizo, M.A., 2011. Evaluación de la eficacia de cuatro enraizadores y tres tipos de estaca en la producción de plantas de guayusa (Ilex guayusa) a nivel de vivero en el cantón Archidona, provincia de Napo. Escuela Politécnica de Chimborazo. Russo, E.B., 1992. Headache treatments by native peoples of the Ecuadorian Amazon: A preliminary cross-disciplinary assessment. Journal of Ethnopharmacology 36, 193–206. Schultes, R.E., 1972. Ilex guayusa from 500 A.D. to the present. Etnologiska Studier 32. Schultes, R.E., 1979. Discovery of an ancient guayusa plantantion in Colombia. Botanical Museum leaflets 27, 5–6. Shemluck, M., 1979. The Flowers of Ilex guayusa. Botanical Museum leaflets 155–160. Tscharntke, T., Clough, Y., Bhagwat, S. a., Buchori, D., Faust, H., Hertel, D., Hölscher, D., Juhrbandt, J., Kessler, M., Perfecto, I., Scherber, C., Schroth, G., Veldkamp, E., Wanger, T.C., 2011. Multifunctional shade-tree management in tropical agroforestry landscapes - a review. Journal of Applied Ecology 48, 619–629. 278 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Anexo 1. Mapa de colección de muestras de guayusa en el Ecuador durante el 2013 279 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 EVALUACIÓN DE ALTERNATIVAS SILVOPASTORILES QUE PROMUEVAN LA INTENSIFICACIÓN Y RECUPERACIÓN DE PASTURAS DEGRADADAS Y CONTRIBUYAN A REDUCIR EL IMPACTO AMBIENTAL DE LA ACTIVIDAD GANADERA EN LA AMAZONÍA ECUATORIANA AL SEGUNDO AÑO DE ESTABLECIMIENTO. Nancy Criollo Rojas, Jorge Grijalva INIAP – EECA / Programa Nacional de Forestería [email protected] Introducción En la Región Amazónica Ecuatoriana (RAE), cerca del 53% de su territorio tiene potencial de uso para bosques, el 25.5%, es área que pertenece al Sistema Nacional de Áreas Protegidas (SNAP) y un 4% pertenece a otros usos; queda solo un 17.5%, o aproximadamente 1.1 millones de hectáreas, que tiene aptitudes de uso para actividades agropecuarias o afines. Por lo expuesto el Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP), a través del programa de Forestería, ha venido generando una serie de tecnologías aplicables en los diferentes pisos altitudinales del Ecuador, para que puedan ser difundidos y aplicados por el sector agropecuario de las diferentes zonas del país. Objetivos Evaluar el comportamiento agronómico de las especies que conforman las combinaciones silvopastoriles en estudio, en la Estación Experimental Central de la Amazonía del INIAP, al segundo año de establecimiento. Determinar el efecto de las alternativas silvopastoriles en estudio sobre las características físicas, químicas y biológicas del suelo, al segundo año de establecimiento Materiales y Métodos El ensayo estuvo constituido por los siguientes tratamientos: Tratamiento 1: Mulato II (Brachiaria hibrido Miles) + Guayaba (Psidum guajava) + Yuca Ratón (Gliricidia sepium Jacq); Tratamiento 2: Mulato II (B. hibrido Miles) + Guayaba (P. guajava) + Quiebra Barriga (Trichantera gigantea); Tratamiento 3: Mulato II (B. hibrido Miles) + Guayaba (P. guajava) + Flemingia (Flemingia macrophylla Wild); Tratamiento 4: Mulato II (B. hibrido Miles) + Guayaba (P. guajava) + Leucaena (Leucaena leucocephala Lam De Wit) ; Tratamiento 5: Mulato II (B. hibrido Miles) + Guayaba (P. guajava) + Botón de Oro (Tithonia diversifolia); Tratamiento 6: Dallis (B. decumbens Stapf) + Guayaba (P. guajava); testigo productor; Tratamiento 7: Mulato II (B. hibrido Miles) + Guayaba (P. guajava); testigo mejorado. Se evaluó variables: suelos, pastos, forrajes leñosos, ambiente y frutal, mediante muestreo sistemático, además se realizó el análisis de presupesto parcial. Resultados y Discusión Suelos 280 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Los resultados obtenidos con las evaluaciones de los indicadores de suelos, no tuvieron diferencias estadísticas significativas entre tratamientos (Cuadro 01), por lo tanto no existió influencia de los sistemas silvopastoriles en las características físicas, químicas ni biológicas. Cuadro 1. Resumen del comportamiento de los indicadores físicos y químicos del suelo en dos años consecutivos de evaluación. *Evaluación inicial – 2011 INDICADOR Densidad aparente Nitrógeno Fósforo Calcio Magnesio 0,734 0,794 0,821 Probabilidad 0,452a 0,687a 0,557a Promedio % 0,46 0,42 0,36 Probabilidad 0,5354a 0,4088a 0,3307a Interpretación contenido18 Muy alta Muy alta Muy alta Interpretación pH19 Optima Media Media Promedio ppm 17,95 14,25 8,25 Probabilidad 0,397a 0,3895a 0,5879a Interpretación contenido20 Normal Normal Baja Mediana Poca Poca Promedio meq/100ml 0,66 0,79 0,86 Probabilidad 0,4179ª 0,6526a 0,743a Interpretación RELACIÓN C/N pH2 contenido3 Normal Normal Normal Interpretación pH2 Optima Media Media Promedio meq/100ml 10,47 10,65 11,9 Probabilidad 0,7746a 0,3064a 0,7739ª Interpretación contenido3 Normal Normal Normal Interpretación pH2 Media Moderada Moderada Promedio meq/100ml 1,72 1,65 1,75 Probabilidad 0,3205a 0,9508a 0,8343ª Interpretación contenido3 Normal Normal Normal Media Moderada Moderada Interpretación MATERIA ORGÁNICA ***Evaluación final – 2012 Promedio gr/cm3 Interpretación Potasio **Evaluación final – 2011 pH2 Promedio % 21,19 20,93 17,04 Probabilidad 0,5309a 0,0702a 0,3419a Interpretación3 Muy alto Muy alto Muy alto Promedio 27,48 27,39 28,17 Probabilidad 0,369a 0,4662a 0,2107ª Interpretación21 Equilibrio entre mineralización e inmovilización Equilibrio entre mineralización e inmovilización Equilibrio entre mineralización e inmovilización Fuente: Laboratorio de suelos Santa Cantalina (2011); Laboratorio de suelos EECA (2012). Cuadro 2. Resumen del comportamiento de los indicadores biológicos del suelo en dos años consecutivos de evaluación. Lugo, (1986) y citado por Pavón, (2010) Diagrama de Troug 20 Rioja, A, (2002), citado por Pavón, (2010) 21 Plaster, E, (2000). 18 19 281 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 INDICADORES NÚMERO DE LOMBRICES PESO DE LOMBRICES Promedio (#/m2) Probabilidad Interpretación Promedio (gr/m2) Probabilidad *Máxima precipitación 2011 *Mínima - precipitación 2011 **Máxima - precipitación 2012 **Mínima - precipitación 2012 19 0,0405a Deficiente 8,03 0,0559a 21 0,2356a Deficiente 10,14 0,4995a 18 0,1142a Deficiente 4,27 0,0503a 63 0,7192a Medio 5,47 0,4085a – Fuente: * Caicedo, W. (2011); ** Criollo, N. (2012) En las especies forrajeras herbáceas Biomasa de la pastura. El promedio de producción de biomasa registrado durante 14 ciclos de pastoreo reflejó diferencias altamente significativas entre los tratamientos establecidos con pasto mejorado versus el testigo agricultor, es decir que la diferencia de producción está determinada por la especie del pasto y no por la influencia del factor en estudio, que viene a ser la forrajera leñosa. Grafico 2. Comportamiento de la producción promedio de M.S/a de las dos especies de pastos, durante dos años consecutivos (14 evaluaciones). La producción de biomasa del año dos disminuyó en aproximadamente el 50% en comparación con las evaluaciones del primer año. Relación Hojas tallo. La relación hojas – tallo, muestra diferencias altamente significativas entre especies de pastos; mientras Brachiaria híbrido, se encontró en rangos de 1.12 a 1.18, el promedio de relación hojas tallo de Brachiaria decumbens es de 0.8; esto obedece a la morfología de cada una de las especies. Valor nutritivo de la pastura. El valor nutritivo de la pastura no muestra diferencia estadísticas significativas en los indicadores de % de Proteína, % de Fibra, y % Fibra detergente neutra, durante los dos años consecutivos de evaluación, lo que supondría que no existió influencia de los tratamientos sobre estos factores en estudio. La diferencia en la FDN entre periodos fue de 29.29%, lo que significó una drástica disminución de la calidad del pasto al segundo año de evaluación. 282 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Grafico 4. Comportamiento de la proteína, fibra y FDN, en evaluaciones de dos años consecutivos, en un ensayo de sistemas silvopastoriles con 7 tratamientos (5 alternativas silvopastoriles + 2 testigos). En las especies leñosas forrajeras Biomasa. La Biomasa de la forrajera leñosa mostró diferencias significativas durante el primer año de evaluación, tal como fue reportado por Caicedo, W., (2012)., cabe resaltar que dichas evaluaciones se realizaron a los 30 días de consumo de los bovinos. Al respecto Caicedo W. (2012), reportó que en todo el periodo de estudio (6 pastoreos), los sistemas silvopastoriles que presentaron mayor rendimiento por efecto de su respectiva leñosa forrajera en materia seca fueron los sistemas con Leucaena (0,32 t/ha/año M.S.), Gliricidia (0,30 t/ha/año M.S.) y Flemingia (0,29 t/ha/año M.S.), mientras que el otro grupo lo conformaron los sistemas silvopastoriles con Quiebra Barriga (0,17 t/ha/año M.S.), y Porotillo (0,10 t/ha/año M.S.), con menor rendimiento. Al segundo año de evaluación la supervivencia de las especies Gliricidia sepium y Trichantera gigantea, se vio afectada por la interacción de los animales. En base a lo mencionado, las especies evaluadas fueron Leucaena leucocephala y Flemingia macrophylla, En dicha evaluación no existió diferencias estadísticas significativas, por lo tanto no existió influencia de los factores en estudio. La producción de forraje en Flemingia macrophylla evaluada cada 60 días reportó 368,72 kg/ha, mientras que Leucaena leucocephala reportó 300,72 kg/ha Valor nutritivo. Los valores de proteína cruda, fibra y fibra detergente neutra presente en las forrajeras leñosas al segundo año de evaluación, pastoreada cada 60 días, presenta diferencias estadísticas significativas entre las dos especies que se evaluaron. Durante el año 1, el contenido de proteína en Flemingia se encontró en 19,32%, mientras que en el año 2, estuvo en 18,14%; lo que significó una disminución de 1.18%; lo que supondría que esta especie es capaz de mantener el porcentaje de proteína con la madurez, tomando en cuenta que el primer año, las evaluaciones se realizaron cada 30 días y el segundo año fueron cada 60 días. En el caso de Leucaena, el contenido en el primer año de evaluación se encontró en 27,14%, mientras que en el año dos disminuyó a 22,63%, lo que significó un decremento de 4,51%. Podríamos atribuir que la edad de pastoreo (30 y 60 días), si causa efecto sobre este indicador, en esta especie forrajera. Con respecto al contenido de fibra, Flemingia reportó 30,94% en el primer año de evaluación y 31,24%, en el segundo año, por lo tanto reportó un aumento de 0,3% de variación entre los dos años, lo que indica que la edad de pastoreo 30 y 60 días, en esta especie, no es un factor modificatorio de este parámetro de calidad de forraje. Para Leucaena, el contenido de Fibra en el primer año se encontró en 17,27% y en el segundo año de evaluación fue de 17,97%, lo que significó un incremento de 0,7%; en este caso, las 283 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 edades de pastoreo, 30 y 60 días en el primer y segundo año respectivamente, no fueron un factor modificatorio de los contenidos. En la especie Animal Ganancia de peso diario. Las ganancias de peso se resumen en gráfico 10. Cabe indicar que al segundo año de evaluación se calculó la ganancia de peso con la digestibilidad del pasto registrada durante el primer año, con animales de 300 Kg de peso. Grafico 10. Comportamiento de la ganancia de peso en bovinos, en evaluaciones de dos años consecutivos, a dos pesos diferentes (478 Kg – año 1; 300Kg – año 2). Capacidad de carga animal. La capacidad de carga no mostró diferencias estadísticas significativas entre tratamientos, lo que indica que no existió influencia de los factores en estudio, sobre este indicador animal. La capacidad de carga animal promedio, calculada con la materia seca producida durante los dos años de evaluación, promedió entre 2.97 y 3.2 UBA’s (animales de 478 kg) por ha, en los tratamientos establecidos con pasto Mulato II, mientras que el pasto Dallis, reportó una capacidad de carga de 2.34 UBA’s por ha. Tomando en cuenta los datos de forraje seco producido al segundo año de evaluación, no existe diferencia entre tratamientos establecidos con los dos tipos de pastos, la capacidad de carga fluctúa entre 1.89 y 2.29 UBA’s (animales de 478 Kg) por ha. En los beneficios no tangibles Temperatura ambiental. La importancia de los árboles en sistemas silvopastoriles se resume en el CUADRO 03. El descenso de temperatura en los bovinos, según Cañas y Gasman, (1987), radica en la zona de termoneutralidad22. A una temperatura ambiente baja, la velocidad metabólica (producción de calor por unidad de tiempo) obedece a la necesidad de calor; cuando la temperatura ambiente aumenta la velocidad metabólica disminuye para mantener constante su temperatura corporal. Esta es una característica de los homeotermos23, actúan como termostato. Sin embargo, si la temperatura ambiental sube por encima de la temperatura crítica (entre 20 y 23°C para animales en ayuno), la producción de calor permanece constante con el aumento sucesivo de temperatura. Cuadro 3. Resumen de las temperaturas ambientales tomadas bajo las Guayabas, en sitios expuestos a la radiación solar y en el bosque secundario colindante con el sistema silvopastoril. La zona de termoneutralidad es una región entre la temperatura mínima crítica y el punto de aumento hipertérmico. Representa el rango de temperatura ambiental en el cual el animal no requiere gastar energía para mantener su temperatura (homeotermos) (Cañas y Gasman, (1987)). 23 Homeotermos.- Díc. de los animales que mantienen la temperatura interna constante e independiente de la del medio ambiente. Son homeotermos los mamíferos y las aves (http://es.thefreedictionary.com/homeotermos) 22 284 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 PROMEDIO ºC DIFERENCIA ºC 10H00 MÁXIMA PRECIPITACIÓN 14H00 SOMBRA DE GUAYABA 29,8 SITIOS EXPUESTOS 30,4 SOMBRA DE GUAYABA 31,6 SITIOS EXPUESTOS 32,7 SOMBRA DE GUAYABA 10H00 SITIOS EXPUESTOS SOMBRA DE GUAYABA 14H00 SITIOS EXPUESTOS SOMBRA DEL BOSQUE MÍNIMA PRECIPITACIÓN 10H00 33,2 33 34,4 1,1 1,2 1,4 26,8 DEFERENCIA ENTRE LA SOMBRA DEL BOSQUE Y LA SOMBRA DE LA GUAYABA 5,2 DEFERENCIA ENTRE LA SOMBRA DEL BOSQUE Y LOS SITIOS EXPUESTOS 6,4 SOMBRA DEL BOSQUE 14H00 32 0,6 27,8 DEFERENCIA ENTRE LA SOMBRA DEL BOSQUE Y LA SOMBRA DE LA GUAYABA 5,2 DEFERENCIA ENTRE LA SOMBRA DEL BOSQUE Y LOS SITIOS EXPUESTOS 6,6 Fuente: Datos primarios SSP – EECA, Tesis Criollo, N. (2012) Carbono total. El sistema que presentó la cantidad de carbono más elevada en el año 2012, fue la constituida por Quiebra barrigo + Guayaba + Pasto, con 168,48 t/ha, seguida por el sistema de Flemingia + Guayaba + pasto con 168.02 t/ha, luego el sistema integrado por Leucaena + Guayaba + Pasto con 167,61 t/ha; y el sistema que menos reportó fue el integrado por Gliricidia + Guayaba + pasto con 147,78 t/ha. Conclusiones Los resultados de la investigación al segundo año de evaluación dieron como resultados que las especies forrajeras leñosas, no tienen efecto significativo en las variables: agronómicas, ambientales ni económicas al segundo año de evaluación. La mejor alternativa silvopastoril en calidad y persistencia para la alimentación bovina es: Pastura + Leucaena + Guayaba, ya que la planta forrajera leñosa presenta alto contenido de proteína, alta digestibilidad y menor fibra detergente neutra, sin embargo la ganancia de peso de los bovinos, debido al aporte de materia seca de esta forrajera, no es significativa con respecto al aporte de materia seca del pasto. Otro aspecto positivo de los sistemas silvopastoriles con árboles de guayaba en los potreros es que reducen de 1 a 1.5°C con respecto a los lugares expuestos a la radiación solar directa. Los sistemas silvopastoriles bajo pastoreo rotativo, evitan la excesiva compactación y erosión de los suelos; y aumenta la capacidad de carga animal de los potreros en una UBA, con respecto a la media nacional. Referencias bibliográficas Caicedo, W. (2012). Evaluación de sistemas silvopastoriles como alternativa para la sostenibilidad de los recursos naturales, en la Estación Experimental Central de la Amazonía, del INIAP. Disponible en: dspace.espoch.edu.ec/bitstream/123456789/2274/1/17T1162.pdf Cañas y Gasman, (1987). Apuntes alimentación animal. VOL I. Pag 113. 285 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Delgado A, JC; Játiva S, P. (2010). Políticas Institucionales de Investigación, Transferencia de Innovaciones y Prestación de Servicios Tecnológicos. Quito Ecuador. Iniap, Dirección General, Dirección de Planificación y Economía Agrícola, 52p, (Publicación Miscelánea No 154) Nieto y Caicedo (2012). Análisis reflexivo sobre el desarrollo agropecuario sostenible en la Amazonía Ecuatoriana. Publicación miscelánea No. 405. Iniap – EECA. Joya de los Sachas – Ecuador. 10, 21pp Pavón, (2010). ANEJO III, Análisis de Suelos. Disponible en: www.ingenieriarural.com/Proyectos/AntonioPavon/05-AnejoIII Plaster, E, (2000). La ciencia del Suelo y su Manejo. Edit. Thompson Paraninfo. USDA, (1999). Guía para la Calidad y Salud http://soils.usda.gov/sqi/assessment/files/KitSpanish.pdf 286 del Suelo publicada. Disponible en: PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Ilex guayusa COMO MOTOR PARA EL DESARROLLO SOSTENIBLE EN LOS SISTEMAS AGROFORESTALES EN LA AMAZONÍA OCCIDENTAL Wain Collen1, Aliana Piñeiro2, Torsten Krause3, Eliot Logan-Hines2 1PlanJunto, Quito, Ecuador; 2Fundacion Runa, Quito, Ecuador; 3Lund University Centre for Sustainability Studies, Lund, Suecia [email protected] Resumen El Ilex guayusa (Loes), o guayusa, es un árbol nativo de la Amazonía occidental. Sus hojas contienen altas concentraciones de cafeína y antioxidantes, y tradicionalmente los pueblos indígenas de la región consumen la guayusa como té. Los árboles toleran sombra y son cultivos perennes que fácilmente adaptan a los sistemas agroforestales tradicionales. Comercialización a gran escala empezó en 2009 iniciado por RUNA, una empresa social con sede en Estados Unidos. Actualmente, RUNA compra hojas de guayusa de más de 2.300 pequeños productores indígenas en la Amazonía Ecuatoriana y vende las fundas de té y bebidas embotelladas en más de 4.500 tiendas y supermercados en los EEUU. Por ser certificados por comercio justo, productores tienen derecho a un premio social además de los ingresos directos por la venta de la guayusa por libra y pueden cosechar guayusa varios veces al año. Esto sugiere un ingreso más continuo comparado con otros cultivos agroforestales en la región. Sin embargo, las primeras fases de la comercialización de guayusa están generando cambios socioeconómicos y de uso del suelo que plantea preguntas sobre el futuro de guayusa como motor de desarrollo sostenible. Se investiga el proceso de comercialización inicial de guayusa en sistemas agroforestales en el contexto socioeconómico y socio-ambiental de los medios de vida de los indígenas en la Amazonía ecuatoriana. En el 2013, a través del apoyo del Programa Manejo Forestal Sostenible en la Región Andina del Instituto Interamericano de Cooperación para la Agricultura (IICA) y el Ministerio de Asuntos Exteriores de Finlandia, Runa ha apoyado el desarrollo de asociatividad entre los agricultores guayuseros y ha dado seguimiento al impacto socioeconómico del proyecto. El objetivo del estudio es identificar bajo qué condiciones la comercialización de guayusa puede contribuir a un desarrollo sostenible en la Amazonía. Los resultados se basan en datos preliminares obtenidos de entrevistas y encuestas con agricultores de guayusa indígenas, junto con datos secundarios. Se aplica el marco analítico de IRF2015 que enfoca en cuatro elementos necesarios para el desarrollo sostenible. Se concluye presentando las principales oportunidades y amenazas potenciales que determinarán el futuro de guayusa como factor que contribuye al desarrollo sostenible de la Amazonía ecuatoriana. Introducción Los sistemas agroforestales representan una relación íntima entre la Amazonía, sus bosques aluviales y tropicales y sus recursos, y las diversas etnias y tribus indígenas que habitan estas zonas. Estudios demuestran que una gran parte del ecosistema amazónico ha sido utilizado por generaciones de los pueblos indígenas (Clement and Junqueira, 2010). Aunque el grado de uso de los sistemas agroforestales depende de la región y la etnia, estos sistemas han sido una solución o adaptación agrícola al ecosistema del bosque tropical, que resiste la producción intensificada, o el monocultivo a largo plazo, por razones de degradación de suelos y plagas (Cochrane et al., 1985). Sin embargo, con la reforma agraria en Ecuador en 1964, que impulsó la migración de grandes números de colonizadores a 287 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 los bosques amazónicos, el conocimiento indígena sobre los sistemas agroforestales se han dejado a un lado. A cambio, en la frontera agrícola amazónica, se ha priorizada la maximización de ganancias económicas a corto plazo a través de la agricultura monocultivo y la ganadería extensiva (Denevan, 1973, Pichón, 1997). Esta tendencia ha contribuido a la deforestación tropical a gran escala (Nepstad et al., 2002), y su impacto positivo sobre el bienestar de los pueblos locales es debatible. Sin embargo, en los últimos años, el interés científico, político y local en sistemas agroforestales ha aumentado como potencial respuesta para enfrentar la destrucción de los bosques amazónicos y para generar beneficios e ingresos económicos más sostenibles (Follis and Nair, 1994). El Ilex guayusa (Loes), o guayusa, es un árbol nativo de la Amazonía occidental que podría ofrecer un ingreso estable para poblaciones indígenas a través de una comercialización que aprovecha de sistemas agroforestales tradicionales. Entre las varias especies medicinales cultivadas en la chacra, Ilex guayusa es la más comúnmente utilizada (Innerhofer and Bernhardt, 2011). En Ecuador, la planta de guayusa ha sido registrada en las provincias de Napo, Pastaza, Sucumbíos, Morona Santiago, Zamora Chinchipe, Pichincha, Loja y Tungurahua sin descartar la posibilidad de su existencia en otras zonas del país (Jørgensen, 1999). La “chacra” Kichwa comprende un área promedio de 0,5 ha (rango 0,25 a 1,5 ha), y por lo general se basa en un sistema agroforestal de subsistencia que combina especies comestibles, maderables y medicinales. Aunque en la chacra el cultivo predominante es la yuca (Manihot esculenta) y sus distintas variedades, las chacras pueden estar compuestas de hasta 48 especies comestibles (Perreault, 2005), y también co-existen plantaciones de orientación más comercial como el café (Coffea arabica), cacao (Theobroma cacao) y naranjilla (Solanum quitoensis). Nosotros investigamos el proceso inicial de comercialización de guayusa en sistemas agroforestales dentro del contexto socioeconómico y socio-ambiental del pueblo indígena Kichwa en la Amazonía ecuatoriana. El objetivo es identificar bajo cuales condiciones la comercialización de la guayusa realmente puede aportar a un desarrollo sostenible. Además, identificamos potenciales áreas, o preguntas (no respondidas en esta investigación) que podrían ser el enfoque de futuras investigaciones sobre la guayusa. Métodos Marco analítico Para responder a los objetivos de este estudio, este artículo aplica principios del marco analítico IRF2015 (IRF, 2013) para analizar la guayusa como impulsor de desarrollo sostenible. Los principios del IRF2015 fueron elaborados por el “Independant Research Forum” para establecer una agenda para el desarrollo sostenible, pos 2015. Estos principios provienen de los acuerdos de la “Agenda 21” (Robinson, 1993), y “la declaración y metas del milenio” (UN, 2000), que apuntaban a mejorar las vidas y reducir la vulnerabilidad de la gente más pobre del mundo para el año 2015. La propuesta de IRF2015 propone que el siguiente era de cooperación internacional (pos, 2015) debe enfocarse en acciones a nivel local, nacional y global, enfrentando los retos económicos, sociales y ambientales entretejidos. De acuerdo al IRF2015, para construir una plataforma para el desarrollo sostenible se requiere dar una balance igual de cuatro elementos interconectados (IRF2015, 2013), (Gráfico 1) 288 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Gráfico 1: Los principios de desarrollo sostenible (Adaptado de IRF2015) Progreso económico: Una economía dinámica forma la base para mejorar los medios de vida de la gente, y para proveer los bienes y servicios necesarios para el desarrollo. Sin embargo, cuando el desarrollo económico no está vinculado a los objetivos y políticas sociales y ambientales, el crecimiento económico podría debilitar a la reducción de la pobreza, aumentar la inequidad, y dañar el medio-ambiente. Ecosistemas productivos y saludables: Una gran parte del sistema económico proviene de los bienes y servicios eco sistémico. Lograr un desarrollo sostenible dependerá en encontrar vías de desarrollo que respeta los limites ecológicos de los ecosistemas (Daly, 1974), además de optimizar la contribución del medio ambiente al progreso económico. La participación de, y colaboración con, actores de interés: Una economía dinámica, un medio ambiente saludable, y bienestar universal va a depender de medidas de refuerzo mutuo, desde los hogares hasta el nivel global. Procesos de participación justo, la transparencia y rendición de cuentas son elementos críticos para construir la cooperación necesaria entre comunidades, gobiernos, el sector privado, y otros actores. Estos procesos van a requerir nuevas políticas, instituciones y maneras de cooperar. Prosperidad y oportunidad equitativa: Investigaciones recientes han demostrado que la inequidad debilita el crecimiento económico, desestabiliza la economía, y frena la reducción de la pobreza (IMF, Banco de Desarrollo Asiático en IRF2015). Además la inequidad es un factor importante que contribuye a la degradación ambiental. Recolección de información Para nuestro análisis, evaluamos la guayusa como un motor de desarrollo sostenible en la amazonía contra estos cuatro puntos. Se levantó información a dos niveles. Primero se hizo una revisión de literatura extensiva sobre temas relacionadas a los objetivos del artículo. Los temas principales para esta revisión incluyeron, guayusa, agroforestería en la amazonía, y desarrollo sostenible. Segundo, se hizo una revisión de información primaria relevante, incluyendo a) una línea base levantado en 2012, b) un estudio de monitoreo elaborado en junio 2013, c) datos de la última certificación orgánica en 2013, y d) datos de la cosecha de guayusa de 2011-2013. Resultados 289 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 En la siguiente sección, hacemos una comparación de la comercialización de guayusa como potencial motor de desarrollo sostenible, y los cuatro indicadores para desarrollo sostenible identificados por el IRF2015. Progreso económico: La comercialización de la guayusa ha generado dos nuevas fuentes de ingresos económicos para los productores de guayusa. El primero, y el principal, es el ingreso directo generado para el productor por la venta de su guayusa. El segundo adicional ingreso es el premio social de comercio justo, que equivale a 15% de todas las compras de guayusa. El precio pagado por la compra de la hoja fresca de guayusa es de $0.35 por libra. Por ser certificado comercio justo, esto es un precio mínimo; es decir que Runatarpuna Exportadora nunca bajará el precio pagado a los productores, no importa las condiciones del mercado internacional. Entre 2011 y 2012 el número de productores que vendió guayusa incrementó de 315 a 695 – un crecimiento de 121%. El ingreso promedio anual por productor incrementó de $52 a $107 o 106% (Ver Tabla 1). Tabla 1: Ventas de guayusa e ingresos al productor Año # Productores Vendedores # Libras de Guayusa Ingreso Total Ingreso Promedio por Productor 2011 315 46.953 $16.434 $52 2012 695 212.598 $74.409 $107 2013 1671* 457.307* $160.057* $96* Fuente: Datos de cosecha Runatarpuna Exportadora, S.A. (*hasta octubre) De acuerdo al estándar de Fair Trade USA (FTUSA), la empresa Runatarpuna Exportadora paga un premio social adicionalmente al pago a los productores por la compra de la guayusa. Este monto va en un fondo aparte y FTUSA requiere que los productores inviertan el premio social en el desarrollo de sus negocios y en el bien de sus comunidades (Glosario de los Estándares de FTUSA, p. 5). Los representantes de los productores toman las decisiones sobre la inversión del fondo y la asamblea general tiene que aprobar estos gastos. Hasta junio 2013, Runatarpuna había pagado un total de $24.259 al premio social y los productores habían gastado $2.924 en asuntos administrativos y $1.860 en pagar una abogada para tramitar el proceso de legalización de las cooperativas y asociaciones. Ecosistemas productivos y saludables: En esta sección, analizamos principalmente los cambios en el uso de suelo que ha promovido la comercialización de guayusa. Antes de iniciar la comercialización de la guayusa, la intensidad de siembra y atención requerida para el árbol era extremadamente baja, como era para uso personal. Con el nuevo modelo de comercialización, y creciente demanda en el mercado de los EE.UU, se exige la siembra de guayusa orgánica. Aunque esto no implica un cambio drástico de la forma tradicional de mantener a los árboles individuales de guayusa, es un nuevo requisito ecológico con que todos los productores tienen que cumplir. Cada año el número de productores de guayusa certificados orgánico aumenta y en mayo de 2013, 2.386 productores eran certificados orgánicos con un total de 265,27 hectáreas. 290 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Un cambio más notable está relacionado con la siembra de guayusa a mayores densidades por hectárea (i.e. entre 50 a 1000 plantas/ ha). Runa busca promover la intensificación del cultivo en el marco de un sistema agroforestal multi-estrato de características comerciales. Algunas de las detalles finales de dicho sistema multi-estrato no están finalizadas, y estarán basado en las conclusiones de un programa de investigación científica que cubre aspectos de la agronomía, genética, ecología, fitoquímica y conocimiento ancestral de la especie. Aunque la información está limitada en cuanto a los cambios de suelo que está generando la intensificación de la guayusa, cifras formales de la última certificación orgánica indica de qué 100% de la guayusa certificada orgánica está en sistemas agroforestales, mezclando la guayusa con otras especies (Ver Tabla 2). Tabla 2: Resumen de certificación orgánica 2013 % Asociado % Asociado # Productores Total Ha. # Parcelas # Plantas Perenes Ciclo Corto 2.386 265,27 3790 329.170 45% 28% Fuente: Certificación Orgánica, Base de Datos, Runa La participación de, y colaboración con actores de interés: % Asociado Maderables 27% En esta sección se enfoca en los procesos de participación en los espacios de toma de decisión y gestión financiera colectiva. En gran parte los lineamientos de participación están definidos por los estándares de comercio justo, que enfatizan una participación de los productores inclusiva y transparente, y RUNA prioriza el cumplimiento con los requisitos. Algunos de los más importantes requerimientos de FTUSA sobre la participación incluyen (FTUSA, 2012): Los representantes de los productores estén elegidos en elecciones libres, justas, transparentes. La asamblea de productores se reúne por lo menos una vez al año Todos los informes, presupuestos, cuentas, y planes de trabajo estén aprobadas por la asamblea, con la mayoría de productores en asistencia Los representantes de los productores cuentan con un proceso de comunicación para asegurar que todos estén informados sobre comercio justo y el manejo del premio social. Todas las actas y decisiones de la asamblea estén registradas En el caso específico de los productores de guayusa, y de acuerdo con los requisitos de comercio justo se eligió un “Comité Ejecutivo de Productores” (CEP) en 2011 para representar a los intereses de los productores. Asegurar el cumplimiento con comercio justo es una responsabilidad compartida entre la empresa y los productores, particularmente en los primeros años, cuando los productores podrían faltar capacidades e instituciones de organización para la producción agrícola. Hasta la fecha se ha mantenido 32 reuniones de cooperación entre RUNA y el CEP y varios talleres de capacitación sobre comercio justo y sobre la gestión financiera. Aunque la composición del CEP ha cambiado varias veces, presentando algunos retos para la continuidad, estas reuniones han sido una plataforma continua para el debate y discusión. A pesar de estos espacios de comunicación entre RUNA y el CEP, las oportunidades para involucrar a los productores mismos en los aspectos de comercio justo y el premio social hasta la fecha han sido limitadas. Además el conocimiento de los productores individuales sobre comercio justo está relativamente bajo. De una encuesta línea base llevado a cabo en Sept 2012 (n=150) – solo el 22% de los encuestados conocía que era el Comité Ejecutivo de Productores de Guayusa. Además, el 77% no sabía cuál era la responsabilidad de este comité. De una encuesta de monitoreo y seguimiento llevado a cabo 291 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 por RUNA en Junio 2013 - solamente el 40% de los encuestados podían identificar los dirigentes de las cooperativas, pero el 87.5% reportaban que participaron en la elección de los candidatos. No fue sorprendente entonces cuando la asamblea general de los productores de guayusa en junio, 2012 (un requisito de comercio justo) tomó la resolución de descentralizar la toma de decisión, y el manejo del premio social a través del establecimiento de siete cooperativas y dos asociaciones productivas. Tramitar la legalización de las cooperativas y asociaciones ha sido una de las principales responsabilidades del CEP. Se prevé que las cooperativas y asociaciones van a estar legalmente conformados a inicios de 2014. Esto significaría que cada cooperativa y asociación descentralizada va a tener que cumplir con los requisitos de comercio justo. Prosperidad y oportunidad equitativa: Históricamente, los pueblos indígenas amazónicos, incluyendo los Kichwa, han sido marginalizado política y económicamente. A través del establecimiento de un mercado internacional de guayusa que ofrece un precio mínimo a los pequeños productores Kichwa por una planta tradicional presenta una oportunidad para aumentar la prosperidad de estas familias rurales. La comercialización de guayusa ha generado un nuevo flujo de ingresos estables para casi 3000 pequeños productores. Además, el éxito de la guayusa está generando nuevas oportunidades para estos productores para acceder a créditos. El Banco Nacional de Fomento (BNF) está ofreciendo créditos – hasta $3000 por hectárea – a pequeños productores para la siembra de guayusa. Dado que una de las limitaciones principales para productores rurales es una falta de acceso a capital, esto presenta una oportunidad concreta para mejorar la posición socio-económica de productores de guayusa. Existen señas también de que la cadena de valor de guayusa está generando oportunidades para las mujeres Kichwa, fortaleciendo la equidad de género. En 2011, el 11% de los miembros del CEP se conformaba por mujeres. Al inicio de 2013, cuando los productores se decidieron organizarse en cooperativas y asociaciones esta cifra subió hasta 42%. Discusión Progreso económico En cuatro años la comercialización de guayusa por parte de Runatarpuna ha logrado resultados concretos en esta área. Por lo general, las poblaciones indígenas amazónicas enfrentan grandes retos para recibir un precio justo para sus productos agrícolas y otros recursos naturales (Mejia, 2013). En muchos casos ellos ocupan la posición más débil en la cadena de valor, dado su falta de experiencia con mercados, y por limitados recursos económicos (Mejia, 2013). Además, para productos en el mercado internacional, como el café y el cacao, los precios están sujetos a cambios constantes en la demanda y precios ofrecidos. Por lo tanto, poder garantizar un precio mínimo a los productores para la guayusa les libera de mucho riesgo proveniente con la siembra de un nuevo producto agrícola. Aunque los ingresos promedio por agricultor todavía no están muy altos, el hecho de que son ingresos confiables está impulsando a ellos elegir la siembra de guayusa encima de café o cacao. Esto podría ser una indicación de que lo que estos productores están buscando, no son solamente altos precios, sino un ingreso confiable y estable. El premio social de comercio justo también demuestra potencial para contribuir al progreso económico de estos productores. Este ingreso ya ha financiado todas las reuniones del CEP, las cuales han sido claves en solucionar problemas a tiempo. Además, el premio social ha financiado una gran parte de los costos de legalización hasta la fecha. Sin embargo, cabe destacar que a futuro el manejo efectivo del premio 292 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 social podría presentar retos para algunas de las cooperativas y asociaciones que no hayan tenido mucha experiencia con la gestión de dinero para fines comunes. Ecosistemas productivos y saludables En la provincia de Napo, la expansión agrícola está en gran parte dominado por la ganadería, y algunos monocultivos. Ambos usos de suelo tiene un impacto generalmente negativo sobre la biodiversidad y otros servicios ambientales. Comparado con estos, el modelo promovido por RUNA es mucho más sostenible. Todos los productores de RUNA tienen certificación orgánica promoviendo ecosistemas productivos más saludables. Además, todos los productores mantienen sistemas agroforestales. Ellos no tienen que dejar de sembrar sus cultivos tradicionales de autoconsumo, y el sistema agroforestal promueve mayores niveles de biodiversidad que el monocultivo. En la práctica, las parcelas de guayusa tienen potencial para actuar como zonas de amortiguamiento en las fronteras de zonas de conservación de bosques primarias. Por ejemplo, alrededor del Bosque Colonos, una área de conservación de 10.000 ha. en la Provincia de Napo, existen comunidades que venden guayusa y la Fundación Runa ha trabajado con ellos para elaborar mapas del uso de suelo para identificar los áreas de conservación y de producción. Dado que Ecuador está estableciendo un programa REDD+ que probablemente tendrá un enfoque en sistemas productivas (UN-REDD, 2013) que reducen la presión sobre el bosque, la guayusa podría ser un ejemplar para cumplir con este objetivo nacional. Aunque actualmente no parece que la siembra de guayusa está incentivando la degradación de bosque primario, sería importante seguir monitoreando este aspecto. Una posibilidad es establecer indicadores para seguir observando las prácticas de siembra, y el cambio en el uso de suelo que promueven. Si el mercado sigue creciendo, es posible que la intensificación de la siembra de guayusa pudiera afectar negativamente a la diversidad de los agro-ecosistemas representados en las chacras tradicionales. Esto podría provocar el estancamiento de la productividad a largo plazo, incrementando la vulnerabilidad a enfermedades y plagas y comprometiendo la provisión de servicios del ecosistema (Tscharntke et al., 2011). La participación de, y colaboración con actores de interés: En vez de definir sus propios procesos de participación, RUNA ha acudido al estándar de comercio justo (FTUSA) para establecer las normas de participación. Los criterios de comercio justo establecen principios ya probadas para fomentar la participación, la comunicación y la transparencia. Durante 20112013, hubo muy buena participación por parte de los representantes de los productores en la toma de decisión y gestión financiera. Sin embargo, el elemento de participación democrática y gestión financiera transparente es algo complejo, y un proceso paulatino. Bajos niveles de conocimiento sobre los reglamentos de comercio justo, y sobre el manejo del premio social entre los productores mismos indican que el proyecto de guayusa todavía debe seguir buscando los mecanismos para cumplir con los estándares de participación de comercio justo – que por lo general están basadas en resultados, no en cómo lograr estos. El establecimiento de cooperativas descentralizadas presenta oportunidades y retos. Los productores van a tener más cercanía a la toma de decisión y el manejo del premio social que podría fortalecer la participación y transparencia. Sin embargo, es probable que muchas de estas cooperativas falten experiencia con la gestión de cooperativas productivas e ingresos comunales. Es posible que algunas cooperativas enfrenten dificultades cumpliendo con los requisitos de comercio justo. Para RUNA existe la pregunta de cómo balancear la eficiencia – un enfoque de trabajo con las representativas, con la equidad – generando capacidades locales para la buena gobernanza. Prosperidad y oportunidad equitativa: 293 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Aunque medir cambios en la equidad es difícil, las indicaciones de este estudio son que la comercialización de guayusa a través de un enfoque en los pequeños productores está dando nuevas oportunidades a grupos históricamente desventajados. A un nivel, se genera un ingreso confiable trabajando con un producto tradicional. A otro nivel, las elecciones de los nuevos representantes para las nueve cooperativas indican que los productores están ubicando a mujeres en posiciones de toma de decisión. En la cultura amazónica, esto podría ser una tendencia importante, dado las dificultades que enfrentan las mujeres en una sociedad que favorece la participación de hombres en espacios de poder. Conclusiones El objetivo de este estudio era de analizar a la comercialización de la guayusa como un potencial motor para el desarrollo sostenible en la frontera agrícola amazónica ecuatoriana. Aplicando el marco analítico IRF2015 que evalúa cuatro áreas de importancia para el desarrollo sostenible, concluimos de que, en gran parte, sí puede ser un motor para el desarrollo sostenible. En solo cuatro años, la comercialización de guayusa ha generado un progreso económico tangible para casi 3000 productores. Sin embargo, la clave ha sido la manera de comercializarla. El modelo sostenible de RUNA esta fortalecido, primero por la aplicación de estándares de comercio justo (FTUSA). La implementación de estos estándares contribuye concretamente a cumplir con tres de las áreas del marco de IRF2015, incluyendo el progreso económico, la participación, y la prosperidad y oportunidades equitativas. En cuanto al área restante del marco IRF2015; ecosistemas productivos y saludables, la certificación orgánica asegura un mínimo impacto negativo para los ecosistemas tropicales. El segundo objetivo de este estudio era de identificar a potenciales áreas que podrían ser en enfoque de futuras investigaciones. Concluimos que todavía existen algunas preguntas sobre dos áreas principales. Primero; cúal impacto podría tener la guayusa en cuanto a cambio de suelo si la demanda para el producto sigue en aumento? Para asegurar que la guayusa no se convierte en un producto que impacta negativamente sobre los bosques tropicales, se recomienda de que Runa identifica e implementa nuevos indicadores que monitorean el cambio del uso de suelo que impulsa la guayusa. Por otra parte, la colaboración entre la empresa, la fundación y las nuevas cooperativas es un elemento que podría presentar varios cambios. Particularmente, si las nuevas cooperativas enfrentan dificultades con la gobernanza y con la gestión del premio social para cumplir con el estándar comercio justo. El trabajo directamente con los productores individuales ha sido una relación estrictamente económica, y más sencilla. Sin embargo, las cooperativas representan a grupos de productores, y manejan fondos para el bien común. Esto implica que el elemento de poder va a ser presente en las decisiones, y aportar con ayuda técnica sobre temas de gobernanza puede ser complejo. No hay respuestas claras todavía, pero esto será un proceso de aprendizaje. Recomendamos que Runa considere la posibilidad de establecer actividades piloto, con el objetivo de generar aprendizajes en conjunto, directamente con uno o dos de las cooperativas que manifiestan directamente que están buscando aliados para cumplir con los requisitos de comercio justo. Bibliografía Clement, C. R. & Junqueira, A. B. 2010. Between a pristine myth and an impoverished future. Biotropica, 42, 534-536. Cochrane, T. T., Sánchez, L., Azevedo, L. D., Porras, J. & Carver, C. 1985. Land in tropical America: a guide to climate, landscapes, and soils for agronomists in Amazonía, the Andean Piedmont, Central Brazil and Orinoco. Part 1. 294 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Daly, H. E. 1974. The economics of the steady state. The American Economic Review, 64, 15-21. Denevan, W. M. 1973. Development and the imminent demise of the Amazon Rain Forest*. The Professional Geographer, 25, 130-135. Follis, M. & Nair, P. 1994. Policy and institutional support for agroforestry: an analysis of two Ecuadorian case studies. Agroforestry systems, 27, 223-240. FTUSA 2012. Estandar para Pequenos Productores Independientes Version 1.0. Innerhofer, S. & Bernhardt, K.-G. 2011. Ethnobotanic garden design in the Ecuadorian Amazon. Biodiversity and Conservation, 20, 429-439. IRF 2013. Post 2015: Framing a new approach to sustainable development. Independant Reseach Forum. JØrgensen, P. M., S. Leon-Yánez (EDS) 1999. Catalogue of the Vascular Plants of Ecuador. Missouri Botanical Garden. Mejía, E. 2013. Aprovechamiento forestal informal en la Amazonía Ecuatoriana asociado a mercados domésticos y transfronterizos de madera. Third IUFRO Latin American Congress. San José. Nepstad, D., McGrath, D., Alencar, A., Barros, A., Carvalho, G., Santilli, M. & Vera Díaz, M. C. 2002. Frontier governance in Amazonia. Science(Washington), 295, 629-631. Perreault, T. 2005. Why chacras (swidden gardens) persist: agrobiodiversity, food security, and cultural identity in the Ecuadorian Amazon. Human Organization, 64, 327-339. Pichón, F. J. 1997. Colonist Land‐Allocation Decisions, Land Use, and Deforestation in the Ecuadorian Amazon Frontier. Economic Development and Cultural Change, 45, 707-744. Robinson, N. A. 1993. Agenda 21: Earth's action plan, Oceana Publications, Inc. Tscharntke, T., Clough, Y., Bhagwat, S. A., Buchori, D., Faust, H., Hertel, D., Holscher, D., Juhrbandt, J., Kessler, M. & Perfecto, I. 2011. Multifunctional shade‐tree management in tropical agroforestry landscapes–a review. Journal of Applied Ecology, 48, 619-629. UN 2000. United Nations Millennium Declaration. resolution adopted, 18. UN-REDD. 2013. Ecuador analiza a la ganadería sostenible como una potencial actividad REDD+ [Online]. Available: http://pnc-onureddecuador.org/comunicamos/noticias/100-ecuador-analiza-a-laganaderia-sostenible-como-una-potencial-actividad-redd.html [Accessed 05/11/2013. 295 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 MEJORAMIENTO DE CHAKRAS, UNA ALTERNATIVA DE SISTEMA INTEGRADO PARA LA GESTIÓN SOSTENIBLE DE BOSQUES EN COMUNIDADES NATIVAS DE LA AMAZONÍA ECUATORIANA Arevalo-Vizcaino, V., Vera-Vélez, R., Grijalva-Olmedo, J. 1Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP). Estación Experimental Santa Catalina. Programa Nacional de Forestería. Panamericana Sur, Km 1 sector Cutuglahua. Quito, Ecuador. [email protected] Introducción La deforestación es un problema complejo que afecta a la población mundial en vista de que el equilibrio climático del planeta depende en gran parte de los bosques. En los últimos años, los índices de deforestación tienden a retroceder en muchos países de Europa, América, Asia y Oceanía, sin embargo, en las mayores reservas de bosques naturales del mundo como en el caso de la Amazonía, sucede todo lo contrario (FAO, 2012). Por otra parte, los índices de pobreza aún son alarmantes, principalmente en África y América Latina, siendo más evidente en el área rural, lo que contribuye a una baja seguridad alimentaria (FAO, 2011). En Ecuador, los datos revelan una gran pérdida de bosques, aunque, su ubicación geográfica y características climáticas sugieren grandes ventajas para repuntar en el ámbito forestal (Grijalva et al, 2012), siendo que con una adecuada gestión, pudiere aportar a la economía del país, mejorar la seguridad alimentaria, y conservar los recursos naturales. En la cuenca del Río Napo, una gran cantidad de bosques se encuentra en posesión de comunidades nativas Kichwas y Colonas, las cuales ejercen presión sobre este recurso en respuesta a las necesidades de subsistencia de sus familias. La presión constante de los pobladores sobre el bosque hace a la zona más vulnerable y con tendencia a perderse en un horizonte relativamente corto. Por otra parte, un sistema tradicional de producción, conocido como chakra, es realizado por los pobladores, este sistema combina atributos de conservación y producción y se presenta como ideal para el manejo integrado de sus recursos. En ese sentido, la Asociación Agro Artesanal de Bienes Agrícolas, Pecuarios y Piscícolas de Napo (KALLARI), organización que agrupa a 21 comunidades que suman alrededor de 850 familias socias (95% indígenas Kichwas de la provincia de Napo), propone un modelo de desarrollo económico alternativo para sus comunidades basado en la venta del cacao nacional fino y de aroma proveniente de las chakras tradicionales, con lo cual han logrado establecer un nicho de mercado a nivel internacional principalmente en Europa. La creciente demanda en cacao y la necesidad de ingresos económicos de las familias, pudiere ser un factor determinante en la deforestación de los bosques, por lo que una gestión forestal encaminada a la conservación de los recursos naturales y la biocultura nativa, y una producción sostenible en beneficio de las familias y los bosques, es urgente. La chakra mejorada mediante un proceso participativo surge como una alternativa ideal que potenciando adecuadamente se ajusta a cubrir las necesidades ecológicas y económicas y alimentarias. Esta metodología puede ser aplicada a diversas localidades con situaciones similares. Objetivos a. Potenciar el sistema de uso tradicional de la chakra nativa, mediante la incorporación de elementos tecnológicos como el cacao nacional fino y de aroma (Theobroma cacao), semillas 296 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 de arroz (Oriza sativa) y maíz (Zea mays) y diversificar el componente arbóreo con especies de uso comercial. b. Contribuir a la seguridad alimentaria al fortalecer el uso de componentes tradicionales como la yuca (Manihot esculenta) y plátano (Musa sp). c. Proporcionar una alternativa de uso del suelo que genere ingresos económicos a las familias de la cuenca del Río Napo y que conjugue la conservación de los recursos del bosque. Metodología Mediante un enfoque transdisciplinario, el proyecto FLOAGRI fue ejecutado a nivel espacial y temporal. La mayor escala geográfica es el de cuenca hidrográfica y el de parcela (chakra) como de menor nivel. El área de trabajo en la Cuenca del Río Napo tiene características de bosque muy húmedo Pre Montano, con una altitud entre los 300 a 600 msnm y una temperatura media anual de 25º C, la precipitación media anual es de 4000 mm. El trabajo fue ejecutado en 4 comunidades que fueron seleccionadas de la asociación KALLARI, estas Comunidades de Referencia fueron: Campo Cocha, Sinchi Runa de Puní Bocana, Asociación Río Blanco, y la comunidad colona Colonia Bolívar, de estas comunidades, un total de 13 predios corresponden a los Predios de referencia. El diagnóstico rural participativo (DRP) consistió en caracterizar 22 chakras. Esto implicó la evaluación del componente arbóreo en riqueza y abundancia, medición de altura total, altura de copa, diámetro a la altura de pecho, y la georeferenciación de cada chakra; finalmente, se determinaron los factores socioeconómicos que influyen en el manejo y las percepciones de los productores hacia los componentes presentes en el sistema, principalmente hacia el componente forestal. Una propuesta tecnológica alternativa fue realizada de manera participativa con los miembros de las familias de cada comunidad para potenciar el sistema chakra, la cual consistió en un conjunto de acciones en predios de uso comunitario, familiar e individual en donde se integraron componentes tecnológicos como el cacao nacional fino y de aroma (Theobroma cacao), arroz (Oriza sativa), maíz (Zea mayz) y el componente forestal con: caoba (Swietenia macrophylla), laurel (Cordial alliodora) y cedro (Cedrela odorata) a los tradicionales componentes de este sistema. El seguimiento técnico y registro in situ de información cuantitativa de las actividades se realizó durante de cinco años, siendo Río Blanco, la última comunidad en incorporarse al proyecto, por lo que de esta solo se tiene información de 3 años. Para la presentación de los resultados en este informe, se lo divide en cuatro secciones, la primera donde se describe brevemente las características de las chakras tradicionales y su funcionamiento, adicionalmente se presenta un cuadro del sistema tecnológico propuesto y otro sobre su implementación; una segunda parte donde se relacionan los tiempos de trabajo, mano de obra y género; una tercera parte donde se analiza el rendimiento de sistema y finalmente una cuarta parte donde se analiza el flujo económico y el destino de la producción de los componentes de la chakra. Para el análisis de los datos se ha utilizado el programa estadístico R (R Core Team, 2012) y el paquete NLME (Pinheiro & Bates, 2011) para realizar el modelo de efectos mixtos. 297 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Resultados Diagnóstico, tecnología propuesta e implementación De acuerdo al Diagnóstico Rural Participativo (DRP), las chakras representan espacios productivos familiares desarrollados en claros del bosque o en realces y que mantienen una alta agrobiodiversidad representada por especies de uso múltiple que se ubican en diferentes estratos y categorías de uso, destacándose el de uso comestible como la yuca (Manihot esculenta), plátano (Musa sp.), maíz (Zea mays), arroz (Oriza sativa), fréjol (Phaseolus vulgaris), maní (Arachis hypogaea), entre otras, que contribuyen a la dieta familiar y se asocian a cultivos comerciales como el cacao (Theobroma cacao), café (Coffea robusta), diversos frutales y maderables para uso doméstico y venta como el cedro (Cedrela odorata), laurel (Cordia alliodora), canelo (Ocotea spp.), caoba (Swietenia macrophylla), chuncho (Cedrelinga catenaeformis), bálsamo (Myroxilum balsamun), tamburo (Vochysia spp), corcho (Apeiba membranacear), aguacate (Persea americana), naranja (Citrus sinensis) y otros cítricos, pambil (Iriartea deltoidea), morete (Mauritia flexuosa), avío (Pouteria spp), así como fibras y semillas para la elaboración de artesanías, cestería y techumbres, entre estas se cuentan la pita (Aechmea strobilacea), chambira (Astrocaryum chambira), paja toquilla (Cardulovica palmata), ungurahua (Oenocarpus bataua), anamora (Ormosa coccinea), San Pedro (Coix lacryma - jobi), caimito muyo (Micropholis chrysophyllum), achira (Canna indica), bulanti u ojo de venado (Mucuna spp) y matiri muyo (Clavija procera). A estas, se suman las plantan medicinales y rituales como ortiga (Urtica dioica), guayusa (Ilex guayusa), ayahuasca (Banisteria quitensis) y el barbasco (Poligonum sp), este último se utiliza como veneno para la pesca. El complejo multiestrato de las chakras comprende un sistema dinámico de uso de la tierra, dejando al suelo en descanso cada cierto periodo de tiempo. El tiempo de rotación depende del rubro de la chakra, estos pueden dividirse en: cacao, yuca, maíz y pastos, donde el primer y último rubro son perennes y los intermedios son temporales. Las chakras de cacao representan el 45% de los sistemas tradicionales, seguidos por los sistemas con maíz en un 27%, yuca con el 23% y finalmente los pastos representan únicamente el 5%. El componente forestal está presente en todas las chakras tradicionales, su densidad promedio es de 179, 22, 91 y 40 árboles/a en cacao, maíz, yuca y pastos respectivamente. La mayoría de los individuos están concentrados en diez especies que representaban el 64% del total. Los sistemas con cacao y maíz aportaban con el 45% y 39% de la riqueza arbórea, mientras que de manera general, las chakras nativas presentan un 62% de riqueza arbórea, frente al 38% de las fincas colonas. Cordia alliodora (Boraginaceae), Cedrela odorata (Meliaceae), Iriartea deltoidea (Arecaceae) y Terminalia oblonga (Combretaceae) eran las especies más numerosas y contaban con el 76% de la abundancia arbórea. A partir de la línea base generada del sistema tradicional de chakra, un esquema mejorado es discutido con las comunidades participantes, dando como resultado una propuesta de chakra mejorada cuya diferencia radicaba en el material genético de los componentes en todos los estratos del sistema y en su manejo agronómico con la finalidad de potenciar el rendimiento con el mínimo uso de insumos externos. Manteniendo la diversidad propia de las chakras tradicionales, el sistema propuesto fue implementado con cacao de material genético seleccionado, que garantiza una producción adecuada tanto en cantidad como en calidad. Una serie de componentes temporales fueron implementándose en torno a este cultivo, tales como el arroz, maíz, maní, fréjol, entre otros, que también provenían de materiales genéticos mejorados. Las comunidades aportaron otros componentes temporales con material genético local los cuales suministraban bienes y servicios propios de la cultura nativa kichwa, en el caso de las 298 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 comunidades colonas, mantenían un sistema acondicionado a sus propias costumbres. De manera simultánea, y como parte del estrato alto del sistema, fueron plantadas tres especies maderables: caoba (Swietenia macrophilla), cedro (Cedrela odorata) y laurel (Cordia alliodora) de procedencias conocidas. Bajo este nuevo esquema, se plantaron 1621 individuos arbóreos de valor económico y 98,1 hectáreas de chakra mejorada con la participación de 79 familias. El área del sistema mejorado implementado variaba desde 0,7 hasta 1,5 hectáreas en fincas colonas, mientras que para las familias kichwas el área implementada oscilaba entre 0,3 hasta un máximo de 1,1 hectáreas. En forma global, el 75% de los árboles fueron plantados en el sistema propuesto y el 25% restante en chakras locales; en cuanto a especies, el 70% de las caobas, el 75% de los cedros y el 82% de los laureles fueron plantados en el sistema propuesto. Las familias no necesariamente siguieron un patrón de siembra establecido (pej.: en línea o cuadrícula), más bien, buscaron cubrir espacios vacíos del sistema. Mantenimiento del sistema Más del 30% del tiempo de trabajo ha sido dirigido al cultivo de cacao (fig. 1), seguido por la yuca (23%), maíz (17%), arroz (14%), plátano (9%), frutales (3%) y un 2% del tiempo fue dedicado a la plantación de los árboles. En el caso del mantenimiento, el 90% del tiempo de trabajo se ha dirigido a la realización de deshierbes (fig. 1: sistema) en el total del área, y no hacia un componente en especial, solamente el cultivo de cacao ha recibido un 10% de tiempo en trabajo para su mantenimiento y el 1% de ese tiempo ha sido para la realización de podas en los árboles plantados. En labores de cosecha, más del 50% del tiempo de trabajo se realiza en el componente cacao y el resto en la yuca, plátano, frutales, arroz, maíz y árboles. Hay que destacar que en los componentes temporales del sistema, las familias han dedicado su tiempo únicamente para la implementación y la cosecha, no han realizado ninguna labor adicional. Los árboles demandan menos tiempo de trabajo por parte de las familias. En cuanto a género, tanto el hombre y la mujer cumplen con su papel en la chakra, según los datos observados, ambos dedican casi el mismo tiempo de trabajo a la implementación de su sistema de producción. El mantenimiento es una actividad que está más relacionada con el género masculino en un 60%, mientras que la cosecha es al contrario, un 60% del tiempo de trabajo está vinculado al género femenino, este último, es debido a que la mujer es la que cosecha los productos de la chakra para la elaboración de alimentos. Figura 1. Relación del tiempo de trabajo con cada componente del sistema Rendimiento de la chakra mejorada 299 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Siendo el cacao nacional fino y de aroma, uno de los componentes principales de la chakra mejorada, un análisis de la varianza en dos vías fue realizado para determinar si existen diferencias en el rendimiento entre los sistemas kichwa y colono con el área (ha) implementada, en vista de que tradicionalmente el sistema colono es ligeramente más intensivo que el kichwa y ocupan áreas más grandes de trabajo. El ANOVA muestra diferencias significativas entre el sistema de manejo kichwa y colono, mientras que no las hay para el área ni para su interacción. Una prueba de TukeyHSD fue realizada para sistema, donde nos indica que el rendimiento del cacao bajo el sistema de manejo de familias colonas es significativamente mayor que el sistema kichwa. Por otra parte no existen diferencias en el rendimiento del cacao para áreas comprendidas entre 0.3 y 1.5 has. La producción media de cacao al cuarto año de establecido el sistema fue de 140,95 kg/ha lo que representa un 55% más de lo estimado en el planteamiento inicial del proyecto y un 80% más que la producción local de 78 kg/ha/año (Arevalo, 2009). Por su parte, el arroz es un importante contribuyente a los rendimientos de la chakra, en vista de que los obtenidos con las variedades introducidas alcanzan los 1318 kg/ha, superando así los 454 kg/ha de variedades locales, esto corresponde a un 300% de incremento en el rendimiento; mientras que en maíz no existe mucha diferencia entre los rendimientos de los híbridos introducidos (2454 kg/ha), con las variedades locales (2272 kg/ha). La yuca produce entre 6 y 9 Tn/ha y el plátano entre 140 y 180 kg/ha. En cuanto al componente forestal, las tres especies plantadas superan en promedio el 70% de sobrevivencia hasta el cuarto año de establecimiento. Esto significa una importante contribución en riqueza arbórea y probablemente un medio de conservación in situ de componentes del bosque. Dado que las especies forestales fueron plantadas tanto en la chakra mejorada como en chakras locales, un ANOVA fue realizado para determinar diferencias en la sobrevivencia de estas, el resultado indica que la sobrevivencia no es afectada por las chakras tanto mejoradas o locales, sin embargo, la tendencia es a obtener una mayor sobrevivencia en las chakras mejoradas con un 81% frente al 67% de las locales; tampoco se halló evidencia de diferencias en sobrevivencia entre especies ni entre sistemas kichwas y colonos, sin embargo, sí existen diferencias en la interacción del sistema por la chakra. De la interacción se obtiene que cuando el componente forestal es instalado en chakras locales, la sobrevivencia está por debajo del 30% en fincas colonas, pero sube al 90% en chakras kichwas, por otra parte, cuando el componente forestal es instalado en chakra mejorada, la sobrevivencia está por sobre el 90% en fincas colonas, pero tiende a disminuir por debajo del 80% en chakras Kichwas. Por otra parte, cuando el ANOVA fue realizado para el crecimiento en altura, no evidenció diferencias estadísticas significativas entre las chakras, sin embargo, se determinó una interacción entre las especies y el sistema. En vista de que el crecimiento en altura es una variable con medidas repetitivas, un modelo lineal de efectos mixtos fue aplicado para explicar el crecimiento en altura en función de los factores: sistema y especie en el tiempo, lo cual resultó en una interacción altamente significativa entre estas. Un efecto del sistema es observado en el crecimiento en el tiempo para las tres especies (fig. 2). Se aprecia una mayor variabilidad a partir del segundo año de medición. El comportamiento en crecimiento parece ser más homogéneo para la chakra mejorada intra e inter especies y más heterogéneo en las chakras locales. El ritmo de crecimiento es rápido para todas las especies, alcanzando los 8 metros en promedio a los cinco años de establecidos, aunque en el caso del laurel (Cordia alliodora) parece alcanzar con mucha facilidad los 10 metros de altura entre los 4 y 5 años en chakras locales. 300 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA 8 6 4 2 0 altura (m) 10 12 14 Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 1 2 3.1.1 3 4 5.1.1 5 1 2.2.1 2 3 4.2.1 4 1.1.1 CAOBA 1 2 3.2.2 3 4 5.2.2 5 1 2.3.2 2 3 4.3.2 4 5 5 1.3.1 1 2 3.3.1 3 4 5.3.1 5 1 2.1.2 2 3 4.1.2 4 5 1.2.2 CEDRO CAOBA LAUREL CHAKRA LOCAL CEDRO LAUREL CHAKRA MEJORADA Figura 2. Crecimiento en altura para las tres especies plantadas en chakras locales y mejoradas en un horizonte temporal de 5 años Flujo económico y destino de la producción El cacao, plátano, la yuca y los árboles, son los principales componentes que contribuyen al ingreso económico de las familias, los primeros tres aportan con 2, 0.31 y 0.25 USD por cada kilo por hectárea, y 315 USD por cada árbol. Mientras que la mano de obra utilizada en las labores agronómicas constituye la principal fuente de egresos, su costo representa 7 USD por jornal por hectárea. Los sistemas kichwas y colonos marcan diferencias en el flujo económico anual de la chakra mejorada (fig. 8), en el caso de fincas de familias colonas, tienden a alcanzar un flujo económico positivo al segundo año de establecimiento, lo cual es bastante rápido si se compara con las chakras kichwas, las cuales no alcanzan valores positivos hasta mediados del tercer año, sin embargo, este sistema sobrepasa en valor económico a los colonos a partir del cuarto año, mientras que el flujo económico de las parcelas de colonos tiende a estabilizarse a partir de ese mismo año. Por otra parte, el aporte del sistema en la seguridad alimentaria viene dado en un 59% por parte del cultivo de yuca, un 27% por el arroz y un 14% por el plátano. Mientras que, del total de la producción del sistema, un 40% es destinado al consumo y un 60% a la venta, los rubros que principalmente aportan a la venta son el cacao y el café. Todos los productos del sistema son vendidos al intermediario común, mientras que en caso del cultivo de cacao, solamente un 10% se destina a la Asociación kichwa KALLARI. 301 PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Figura 8. Flujo económico anual para la chakra mejorada Discusión Las chakras tradicionales son áreas destinadas a la producción de cultivos principalmente para el autoconsumo, se establecen cerca de las casas y producen por dos a tres años. La intensidad de explotación y fertilidad de las chakras determina el tiempo en la cuales entran a una fase de descanso denominado realce y/o bosque secundario; por uno a tres años, el realce se desarrolla sin ser intervenido, se inicia la formación del bosque secundario en donde las especies pioneras, de rápido crecimiento aparecen y lo poblan. En otros casos, los periodos de descanso se han acortado para la intensificación de cultivos anuales, aumentando el retorno económico a corto plazo, lo que posiblemente ocasione que los patrones sostenibilidad vayan disminuyendo. Una interacción positiva resultó en la ejecución del proyecto FLOAGRI con la población local, que se ve reflejada en la implementación de las 98.1 has de chakra mejorada, esto significa una recuperación del componente arbóreo como lo es la plantación de tres especies forestales: caoba (Swietenia macrophylla), cedro (Cedrela odorata) y laurel (Cordia alliodora), pero mejor aún, el interés en estas especies forestales ha incidido en que se logre una sobrevivencia promedio del 77%, esto podría confirmar una estrecha relación entre la población nativa y el bosque y su interés por la conservación (Arevalo, 2009). Por otra parte, el éxito logrado en la implementación del cacao nacional fino y de aroma (Theobroma cacao), se podría deber a una fuerte influencia del creciente mercado internacional, el cual exige grandes cantidades de producto, lo que significa una oportunidad de ingresos adicionales a las familias y evidentemente satisfacer sus necesidades básicas insatisfechas, esto podría corresponder en que el 71% de la población de esta región con catalogados como pobres en función de sus necesidades básicas insatisfechas (INEC, 2012), y donde el proyecto FLOAGRI ha incidido en la utilización de material genético adecuado que garantice tanto calidad como cantidad. La relación género-chakra, se ve correspondido en el papel que cumple tanto el hombre como la mujer en las actividades; en el caso de la mujer, obedece a un estrecho vínculo con la chakra, pues se le considera como de carácter exclusivo de ésta (Arevalo, 2009), y los resultados reflejan precisamente ello, en vista de que es la PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 mujer la que cumple mayormente el papel en las cosechas, probablemente se deba a que ocupa una gran parte del tiempo en la cosecha de la yuca y el plátano para la elaboración de comidas tradicionales como la “chicha” o el “guarapo” para el hogar, por su parte el hombre colabora con el restante de actividades en la chakra como la implementación y el mantenimiento. El componente forestal parece favorecerse por las actividades agronómicas realizadas en la chakra, lo que se refleja en el crecimiento, sin embargo, la constancia que representa el manejo más intensivo de la chakra mejorada parece favorecer un crecimiento más homogéneo de la masa y que podría ser determinante a futuro en la calidad de la madera. Por otra parte, los rendimientos obtenidos tanto en el cacao como en el arroz son significativamente superiores a los obtenidos por el material genético local, esto significa una importante contribución en los ingresos económicos y de alimento para las familias, aunque era de esperarse, sobre todo en el cacao, teniendo en cuenta que es de origen natural Amazónico. Al parecer el sistema de manejo colono favorece un mayor rendimiento del cacao, esto podría obedecer a que estas comunidades dependen casi exclusivamente de este cultivo como fuente de ingreso para cubrir sus necesidades, por lo que su dedicación sería mayor en lo que respecta a la realización de labores agronómicas que incrementen el rendimiento, a diferencia de las comunidades nativas Kichwas que diversifican su fuente de ingresos tanto económicos como alimenticios. La chakra mejorada es un sistema que puede contribuir positivamente a los ingresos económicos familiares a partir del segundo año de establecimiento, sobre todo cuando las fuentes de ingresos son más diversificadas, aunque en promedio superan el flujo económico negativo a partir del tercer año que es cuando el cacao entra en producción, a partir de ahí la mayor dependencia económica está en función de este cultivo. El sistema propuesto también contribuye positivamente con la seguridad alimentaria con un 40% de la producción destinada a la alimentación de las familias, esto significaría un aporte de 237 Kcal/100g de yuca, 122 Kcal/100g de plátano y 354Kcal/100g de arroz para completar las 2752 Kcal requeridas por persona día y reducir la pobreza de consumo que está situada en un 60% en esta región (INEC, 2012). A pesar de que la Asociación KALLARI tiene un mercado internacional directo, las comunidades tienden a vender el 90% de la producción de cacao de sus chakras al intermediario común, probablemente se deba a que no hay una diferencia significativa entre el precio que paga el intermediario común y el que paga la Asociación KALLARI por cada kilo de cacao, por lo que al productor no le representaría una ganancia significativa. Este es un tema clave, en vista de que uno de los objetivos del fortalecimiento de la cadena de cacao es precisamente la reducción de la intermediación para que esas ganancias vayan en favor de los asociados, sin embargo, se puede asumir que los productores aún no sienten ese beneficio extra de ser miembros de la Asociación KALLARI en términos de ingresos económicos o un beneficio tangible para su comunidad. Conclusiones La chakra mejorada representa un modelo potenciado del tradicional que combina la conservación in situ de recursos boscosos como la producción diversificada con componentes que generan significativos ingresos económicos como alimentarios y su implementación en las diferentes localidades obedece a una participación activa de la población en la toma de decisiones sobre el uso de su territorio. PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 La seguridad alimentaria se ha visto fortalecida con este sistema de uso de la tierra, principalmente con los componentes temporales como el arroz, yuca y plátano, lo cual es interesante para reducir los altos índices de pobreza de consumo de esta región. El modelo desarrollado corresponde a una alternativa viable que puede ser extendida a los productores de la región de la cuenca hidrográfica del Río Napo. Bibliografía Arevalo, V. 2009. Chakras, bosques y ríos: El entramado de la Biocultura Amazónica. Publicación miscelánea No. 148, INIAP. 137. Quito. FAO. 2011. Los Bosques para una mejor nutrición y seguridad alimentaria. FAO. 11. Roma. FAO. 2012. El estado de los bosques del mundo. FAO. 52. Roma. Grijalva, J.; Checa, X.; Ramos, R.; Barrera, P.; Limongi, R. 2012. Situación de los Recursos Genéticos Forestales – Informe País Ecuador. INIAP-FAO. 95. Quito. INEC. 2012. Las condiciones de vida de los ecuatorianos: Pobreza y Desigualdad. INEC. 16. Quito. Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Development Core Team. 2013. nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-108. R Core Team. 2012. A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna. PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 RESÚMENES DE PONENCIAS Isabel Aulló, INIAP EVALUACIÓN DEL ESTADO DE CONSERVACIÓN DE DOS ESPECIES FORESTALES PRIORIZADAS EN LA SUBCUENCA DEL RÍO QUIJOS, NAPO, ECUADOR Mauricio Cáceres, Instituto Espacial Ecuatoriano ESTADO DEL BOSQUE PROTECTOR CHONGÓN Y COLONCHE SUBCUENCA LOMA ALTA Y AMPLIACIÓN, CORDILLERA CHONGÓN COLONCHE Y LA SUBCUENCA DEL RÍO CHONGÓN EN EL CANTÓN SANTA ELENA Pedro Ramírez Torres, GIZ-Programa GESOREN “INCORPORACIÓN DE ÁRBOLES MADERABLES DE ALTO VALOR EN PLANTACIONES NUEVAS DE CACAO FINO EN EL NORTE DE LA AMAZONÍA ECUATORIANA” Eduardo Cueva, Naturaleza y Cultura Internacional CRECIMIENTO Y FENOLOGÍA DE 13 ESPECIES ARBÓREAS DEL BOSQUE SECO TUMBESINO RESERVA NATURAL LAIPUNA – MACARÁ – LOJA – ECUADOR Paulo Barrera, INIAP PROSPECCIÓN DE SEMILLAS DE ESPECIES LEÑOSAS NATIVAS DE USO MÚLTIPLE (AUM) PRIORIZADAS POR LAS COMUNIDADES KICHWAS CON POTENCIAL AGROFORESTAL PARA EL MANEJO SOSTENIBLE DE LA TIERRA David Alejandro Romero, Jardín Botánico Milpe ORQUÍDEAS: PRODUCTO FORESTAL NO MADERABLE (PFNM) PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 José Ochoa, INIAP USO DE LA DIVERSIDAD GENÉTICA PARA LA CONSERVACIÓN DEL BOSQUE: CASO DEL CULTIVO DE LA NARANJILLA EN ECUADOR Carlos Nieto, PUCE EL GUARANGO (Caesalpinea spinosa Kuntze), UNA ESPECIE FORESTAL DE PROPÓSITO MÚLTIPLE PARA HACER FORESTACIÓN PRODUCTIVA Alvaro Cañadas, Universidad Técnica del Norte CONDICIONES DE SITIO Y CRECIMIENTO DE Centrolobium ochroxylum ESPECIE NATIVA CON POTENCIAL AGROSILVOPASTORIL DEL LITORAL ECUATORIANO? PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 EVALUACIÓN DEL ESTADO DE CONSERVACIÓN DE DOS ESPECIES FORESTALES PRIORIZADAS EN LA SUB-CUENCA DEL RÍO QUIJOS, NAPO, ECUADOR Aulló, Isabel; Jorge Grijalva; Raúl Ramos, Paulo Barrera Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP). Estación Experimental Santa Catalina. Programa Nacional de Forestería. Panamericana Sur, Km 1 sector Cutuglahua. Quito, Ecuador. [email protected] La sub-cuenca del valle del río Quijos conforma un entorno de inmensurable valor ecológico; sus bosques, sus ríos, sus parques naturales y su gran diversidad forman parte de la gran riqueza natural que posee el Ecuador. Estas riquezas conforman una herramienta indispensable en la lucha contra el cambio climático al constituir un sumidero de carbono y ser fuente de recursos genéticos forestales indispensables en futuras acciones de conservación. No obstante, esta riqueza se está viendo mermada y amenazada principalmente por la deforestación que desde hace más de cinco décadas está deteriorando el legado natural a favor de la expansión ganadera: degradando los recursos hídricos y forestales, empobreciendo la biodiversidad y disminuyendo la capacidad de respuesta frente al cambio climático. A esta realidad se suma el inconveniente de la falta de información sobre el estado real de los recursos del bosque. En este contexto, es necesaria la realización de una investigación que aporte información fidedigna de la situación de estos recursos y del alcance de esta pérdida natural, siendo imprescindible comenzar por el estudio de aquellas especies más valiosas para la población, determinando su estado de conservación mediantela aplicación de normas internacionales que informen del peligro que afronta cada taxón, aportando argumentos sólidos en los que sustentar futuros proyectos de defensa y protección de los mismos. Encuestas realizadas mediante Diagnóstico Rural Participativo a representantes de las parroquias pertenecientes a la sub-cuenca dieron como resultado la lista de las diez especies más valoradas, de entre las que se seleccionaron las dos primeras como objeto de la investigación, esto es: el Cedro, Cedrela montana Moritz ex. Turcz, y el Porotón, Erythrina edulisTriana ex Micheli. Con la colaboración del Herbario Nacional del Ecuador, la Pontificia Universidad Católica del Ecuador, la Universidad Nacional de Loja, la Escuela Superior Politécnica de Chimborazo, la Universidad Técnica del Norte y con los datos online del herbario de Azuay, se confeccionó una base de datos con la localización georreferenciada de muestras de estas dos especies. Utilizando estos datos como inputs para el programa MAXENT, además de los datos climáticos disponibles en www.worldclim.org, se elaboró un mapa que muestra en términos de probabilidad la distribución de las dos especies. Sobre dichos mapas y mediante utilización de herramientas SIG, se planificarán los muestreos a realizar durante los próximos meses. Durante estos muestreos, estratificados y por transectos, se georreferenciarán los individuos de las especies de interés y se obtendrán datos con los que se creará un nuevo mapa de probabilidad de ocurrencia. Finalmente, aplicando a este caso en particular los criterios de la Lista Roja de la UICN a nivel regional, y comparando los dos mapas de probabilidad de ocurrencia obtenidos, previos y posteriores al muestreo, se determinará el estado de conservación de las especies Cedrela montana Moritz ex. Turcz y Erythrina edulis Triana ex Micheli en la sub-cuenca del río Quijos. Palabras clave: Conservación, Recursos genéticos forestales, Deforestación, Cambio climático PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 ESTADO DEL BOSQUE PROTECTOR CHONGÓN Y COLONCHE SUBCUENCA LOMA ALTA Y AMPLIACIÓN, CORDILLERA CHONGÓN COLONCHE Y LA SUBCUENCA DEL RÍO CHONGÓN EN EL CANTÓN SANTA ELENA Mauricio Cáceres, Silvia Román, Misael Yánez Instituto Espacial Ecuatoriano [email protected] El presente trabajo se realizó dentro del marco del Proyecto Generación de Geoinformación para la Gestión del Territorio a Nivel Nacional, se caracterizó la cobertura ecosistémica y uso del suelo del cantón Santa Elena, y un segmento del Bosque Protector Chongón Colonche contenido dentro del mismo cantón, que forma parte del PANE declarada por el MAE, el cual nos permitirá delimitar las zonas con actividades antrópicas para implementar planes de conservación y refuerzo en las áreas afectadas y a la vez implementar PDOT dentro del cantón. Su desarrollo se basó en la utilización de herramientas SIG insumos cartográficos e interpretación de imágenes mediante el procesamiento digital de las ortofotos e imágenes satelitales, la misma que sirve para una interpretación interdependiente mediante la generación de polígonos que delimitaron las zonas agropecuarias y zonas de cobertura vegetal natural. La determinación de los bosques protectores se realizó mediante la superposición de las capas de la información generada por el MAE. La verificación y comprobación en tiempo real de unidades se realizaron en el campo en el año 2012. Dentro del cantón Santa Elena el Bosque Protector Chongón Colonche ocupa una área de 61 041,045 ha, que corresponden a: Loma Alta y Ampliación, Cordillera Chongón Colonche y la Subcuenca del Río Chongón. Las actividades agrícolas y pecuarias dentro de este Bosque ocupan un área de 3 762,68 ha que constituye el 6,21 % en los cuales se destaca el maíz, ciruelo, paja toquilla, y presencia de pastos cultivados para actividades pecuarias, entre los más preponderantes. La cobertura vegetal natural ocupa 57244,83 ha es decir el 93,78 % entre los que predomina el Bosque Húmedo en las zonas altas, también están el bosque seco, Matorral seco, matorral húmedo, vegetación herbácea seca y vegetación herbácea húmeda. La información del estudio realizado permitirá apoyar la toma de decisiones en el campo de la conservación y los proyectos de desarrollo. Los Gobiernos Locales y sus dependencias, deberían implementar políticas que den un enfoque de participación y coordinación entre las autoridades y la comunidad para la conservación de los ambientes naturales y el manejo adecuado del territorio a su cargo. Donde además es necesario reforzar el trabajo de las Áreas Protegidas para la conservación de los ecosistemas. Palabras claves: agrícola, antrópicas, geoinformación, pecuario, SIG PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 PROYECTO: LOS ÁRBOLES SON MI JUBILACIÓN “INCORPORACIÓN DE ÁRBOLES MADERABLES DE ALTO VALOR EN PLANTACIONES NUEVAS DE CACAO FINO EN EL NORTE DE LA AMAZONÍA ECUATORIANA” Pedro Ramirez Torres GIZ-Programa GESOREN [email protected] El objetivo principal de esta medida de desarrollo es promover la incorporación de especies maderables nativas de alto valor en cultivos nuevos de cacao fino buscando mejorar la sustentabilidad del sistema productivo en previsión al cambio climático y a largo plazo contribuir a la “jubilación” de hombres y mujeres del campo, es decir, a través de la venta de madera después de varios años, cuando los actuales productores se encuentren inactivos. Los resultados avanzados desde inicios del 2012 han sido la homologación de criterios técnicos con las instituciones contrapartes de la Mesa de Cacao de la Reserva de Biosfera Sumaco para la selección de especies, la forma de integrar a los sistemas agroforestales-SAF o sistema chakra, y la capacitación y el acompañamiento a las comunidades beneficiarias. Se ha adelantado la siembra de 248.000 árboles maderables especialmente de las especies Chuncho Cedrelinga catenaeformis y Balsamo Myroxylon balsamum, dentro de 320 ha de cacao y café, beneficiando a 496 familias de comunidades Kichwas, Waorani y Colonas de Napo y Orellana, donde las mujeres y jóvenes han sido los más interesados en perspectiva de un mejor futuro econòmico. La siembra se ha realizado bajo diseños específicos en hileras, en el contorno o en los espacios vacios, y la densidad ha variado de 48 a 168 árboles/ha, según las características encontradas en cada parcela, y en base a los criterios del productor beneficiario. Los avances de esta medida permite concluir que el cacao tipo Nacional al ser originario de la selva amazónica está habituado de manera ancestral a vivir bajo otros árboles lo que da oportunidad a sembrar especies de alto valor económico que fortalecen el sistema de cultivo; así mismo para las comunidades indígenas que manejan el sistema Chakra resulta menos controvertida la inclusión de árboles de sombra, sin embargo en estas condiciones resulta más difícil la ordenación de los SAF. Finalmente existen retos, por un lado en generar información técnica y científica de respaldo sobre las especies forestales nativas de interés y por otro en aumentar la disponibilidad de fuentes semilleras confiables para producir plantas y satisfacer la enorme expectativa y la motivación comunitaria existente en la Amazonía ecuatoriana. Palabras claves: árboles maderables, cacao y mi jubilación PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 CRECIMIENTO Y FENOLOGÍA DE 13 ESPECIES ARBÓREAS DEL BOSQUE SECO TUMBESINO. RESERVA NATURAL LAIPUNA – MACARÁ – LOJA ECUADOR Eduardo Cueva, José Acaro A Naturaleza y Cultura Internacional, Pío Jaramillo Avenue and Venezuela street 1101-332, Loja, EC [email protected] El bosque seco tropical mundialmente conocido como uno de los biomas más amenazados, posee un endemismo superior al 20% de flora y aun superior para otros grupos biológicos en el Sur de Ecuador y en el norte de Perú (Loja y Tumbes), estos bosques con seguridad tienen el mejor estado de conservación y se constituyen en el centro de la zona de “Endemismo Tumbesino”. En la Reserva Laipuna se viene investigando desde el año 2003 el crecimiento y fenología de 13 especies de árboles frecuentes distribuidos entre 600 y 1100 m s.n.m. Los resultados muestran claramente especies de rápido, mediano y lento crecimiento. Las de rápido crecimiento son las de madera suave y lento crecimiento las especies de mayor valor maderable. El crecimiento de los árboles se presenta en el periodo de lluvias (Febrero – Mayo) y los mayores crecimientos se dieron el año 2006 y 2012; y en los periodos más secos del año 2006, 2007 y 2011 se han encontrado especies que han sufrido un decrecimiento. Las especies que muestran la tasa de crecimiento mayor son: Ceibo Ceiba trichistandra 0.71 cm/año y Polo polo Cochlospermun vitifolium 0.67cm/año. En el grupo intermedio encontramos el porotillo Erythrina velutina con 0.41, pasallo Eriotheca ruizii con 0.25, gualtáco Loxopterigium huasango y faique Acacia macracantha ambos con una tasa de 0.35 cm/año. En el grupo de lento crecimiento tenemos: laurel Cordia alliodora con (0.20), sota (0.18), guarapo Terminalia valverdae (0.15), charán Caesalpinia glabrata (0,14) y guayacán Handroanthus chrysantha (0.10). Podemos concluir que los árboles crecen en el periodo de lluvias y en cuanto a la fenología en la floración existe una distribución de las especies durante todo el año, pero se observar ciertos picos en el segundo y tercer trimestre del año. Con la fructificación también se observa una distribución durante el año pero hay una mayor sincronización en el tercer trimestre principalmente en septiembre. Palabras clave: Fenología, bosque seco, especies nativas, restauración PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 PROSPECCIÓN DE SEMILLAS DE ESPECIES LEÑOSAS NATIVAS DE USO MÚLTIPLE (AUM) PRIORIZADAS POR LAS COMUNIDADES KICHWAS CON POTENCIAL AGROFORESTAL PARA EL MANEJO SOSTENIBLE DE LA TIERRA J. Grijalva, P. Barrera Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP). Estación Experimental Santa Catalina. Programa Nacional de Forestería. Panamericana Sur, Km 1 sector Cutuglahua. Quito, Ecuador. [email protected] En el Ecuador, la producción forestal ha estado circunscrita al aprovechamiento de las especies sin ninguna estrategia que minimicen su degradación, con la consiguiente erosión genética y el agotamiento de las poblaciones de las especies forestales. Esta investigación se desarrolla en el marco de la conservación de los individuos de interés para las comunidades Kichwas y la disminución de la presión sobre el bosque primario. Para este estudio participativo se priorizaron, en las comunidades kichwas de Chimborazo y Napo, 9 especies forestales nativas: Yagual (Polylepis sp), Quishuar Buddleja incana. Tilo Sambucus nigra. Chuncho Cedrelinga cateniformis. Balsamo Myroxylon balsamum. Ahuano Swietenia macrophylla. Cedro Cedrela montana. Porotón Erythrina edulis. Motión Hyeromina sp). Estas especies forestales son de importancia debido a sus usos múltiples (AUM), entre las que destacan: Leña, Asociación con cultivos, Medicinal, Artesanías, Alimento para animales, Tintes naturales, Protección de fuentes de agua, Madera para construcción. Posteriormente con ayuda de expertos locales se realizó acciones de prospección de especies forestales con búsquedas exhaustivas de individuos con el fin de evaluarlos, (fenotipo, fenológica y fisiológicamente), teniendo entre 10 y 15 procedencias para cada especie forestal, identificados los individuos se procede a georreferenciar (mapa de accesiones por individuo), todo este proceso con el fin de tener una gran diversidad genética y determinar el nivel de variación y la relación genética existente entre las poblaciones. Dentro del estudio se evaluó árbol élite que son los que se muestran fenotípicamente superior al resto de los individuos de su especie, el registro de la variación de las características fenológicas y el conocimiento de la calidad del material forestal de reproducción se reconoce de suma importancia no solo en la comprensión de la dinámica de las comunidades forestales, biodiversidad y productividad sino también es una herramienta fundamental para la consolidación de programas de conservación de los recursos forestales, gestión forestal sostenible que desarrolla el Ministerio del Ambiente (MAE), la información generada contribuye a la política nacional de forestación y reforestación productiva enfocada en el desarrollo social, económico y ambiental, a través de la producción de madera, punto de partida para programas de mejoramiento genético forestal de la subsecretaría Nacional forestal (MAGAP). Palabras clave: Prospección, semillas, priorización PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 ORQUÍDEAS: PRODUCTO FORESTAL NO MADERABLE (PFNM) David Alejandro Romero Castellanos JardínBotánico Milpe (JBM) [email protected] Dentro de un bosque nativo, los Productos Forestales No Maderables (PFNM); es todo aquello, que no sea madera y que tenga un origen orgánico. La orquídea es un PFNM y por ser una planta principalmente epifita (crecen sobre otras plantas sin parasitarlas); su hábitat son los bosques nativos del país. Hace 10 años se empezó en el Noroccidente de Pichincha un proyecto de conservación y aprovechamiento sostenible de un bosque nativo. El Jardín Botánico Milpe (JBM) es el resultado de dicho proceso. El JBM presenta el siguiente trabajo científico voluntario en el cual se realiza una descripción de diferentes especies de orquídeas las cuales como un producto del bosque deben ser conservadas y aprovechadas sosteniblemente. Entre los principales géneros de orquídeas que habitan los bosques nativos del Ecuador; tenemos a: Bosque Nublado: Dracula, Odontoglossum, Lepanthes y Platystele, Bosque Cálido-Húmedo: Huntleya, Gongora y Vanilla; y Bosque Seco: Cattleya. Dichas orquídeas por su potencial comercial y biológico son requeridas para la industria de flor de corte, aromas florales, fitomejoramiento y coleccionistas. La conservación de bosques nativos es una prioridad, si se quiere aprovechar plantas como las orquídeas; que representan más del 20% de la flora ecuatoriana y que aportan al fitoendemismo del país con 1707 especies. Palabras clave: Producto Forestal No Maderable (PFNM), epifita, orquídea, endémica PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 USO DE LA DIVERSIDAD GENÉTICA PARA LA CONSERVACIÓN DEL BOSQUE CASO DEL CULTIVO DE LA NARANJILLA EN ECUADOR J.B. Ochoa1; N. Amagua2; V. Barrera3, M. Ellis4 and J. Alwang5 1Departamento de Protección Vegetal, Est. Exp. Santa Catalina, Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP), Quito-Ecuador. 2Facultad de Ciencias Agrícolas, Universidad Central del Ecuador, Quito-Ecuador. 3Departamento de Ganadería, Est. Exp. Santa Catalina (INIAP), Quito-Ecuador. 4Plant Pathology Department, Ohio State University, Wooster-USA. 5Department of Economy, Virginia State University, Blackburg-USA [email protected] En Ecuador, el cultivo de la naranjilla (Solanum quitoense Lamark) ha sido exitoso casi exclusivamente en el bosque primario. Los agricultores afirman la imposibilidad de cultivos sucesivos de naranjilla. El cultivo de la naranjilla por lo tanto ha contribuido con la deforestación de las dos estribaciones de la cordillera de los Andes del país. Adicionalmente, la naranjilla domesticada ahora llamada “común” llego casi a su extinción a inicios de los 1980’s dando paso al cultivo de los híbridos Puyo, INIAP-Palora y Espinudo. Estudios biológicos y epidemiológicos de Fusarium oxysporum f. sp quitoense causante de la Fusariosis de la naranjilla han permitido establecer la causa del colapso de la naranjilla y la necesidad del agricultor de migrar al bosque primario. Para evitar el inóculo del patógeno, el agricultor ha movido el cultivo al bosque primario, pero paralelamente el patógeno se ha movido a través de la semilla y se ha establecido en el suelo por medio de clamidosporas (esporas de reposo), por lo que el agricultor ha tenido que continuar migrando al bosque primario. Una opción de manejo es usar semilla sana o desinfectada, pero el patógeno esta en muchos suelos agrícolas, por lo que la opción más práctica es el uso de la resistencia genética. En estudios sistemáticos y complementarios se identificó y caracterizó la resistencia genética a F. oxysporum f. sp quitoense en accesiones de la sección Lasiocarpa del banco de germoplasma del INIAP, mientras que todas las accesiones de S. quitoense fueron susceptibles. El proceso más rápido para usar la resistencia genética identificada es el uso de patrones resistentes. En estudios de evaluación de patrones, se seleccionó a la accesión ECU-6242 de Solanum hirtum como la más estable y permite el cultivo de la naranjilla común hasta en cuatro ciclos sucesivos, y por lo tanto el cultivo en suelos agrícolas, contribuyendo con la conservación de los bosques en las estribaciones de los Andes. La siguiente etapa para explotar la resistencia genética es la obtención de variedades resistentes. Al momento se han desarrollado poblaciones y líneas resistentes a F. oxysporum f. sp. quitoense en cruzas simples y retrocruzas entre S. quitoense con S. hiporodium, S. felinum, S. vestissimun y S. hirtum. El desarrollo de variedades resistentes a F. oxysporum f. sp. quitoense contribuirá significativamente al cultivo ecológico de la naranjilla en el Ecuador. PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 EL GUARANGO (Caesalpinea spinosa Kuntze), UNA ESPECIE FORESTAL DE PROPÓSITO MÚLTIPLE PARA HACER FORESTACIÓN PRODUCTIVA Carlos Nieto Cabrera Ph.D. Escuela de Ciencias Geográficas, Pontificia Universidad Católica del Ecuador, PUCE. Av. 12 de Octubre 10076 y Roca, Quito. [email protected] El Guarango, Campeche o Vainillo, conocido internacionalmente como Tara, es una especie Forestal de Propósito Múltiple, (FPM) típica, apta para programas de forestación productiva. Produce madera dura, apta para varios usos, es una planta melífera, forrajera, medicinal y produce frutos aptos para la industria, con dos derivados principales: Taninos, derivados de sus vainas, para uso en industrias como: curtiembre de pieles, pinturas, plásticos, barnices y otros y; Gomas, derivadas de sus semillas, para uso como espesante en la industria alimenticia. El carácter de FPM se complementa con la aptitud de la especie para reforestar áreas degradadas del ecosistema Bosque Seco Tropical, BST, que propicia la mejora del entorno ambiental y del paisaje y facilita la recuperación de la biodiversidad local. En esta ocasión, se presenta los resultados principales de varios proyectos de investigación y promoción de la especie, realizados durante el periodo del 2006 al 2012, con el apoyo de varias instituciones, entre ellas la Fundación Desde el Surco, la Secretaria Ambiental del Distrito Metropolitano de Quito, y la SENESCYT y cuyo objetivo principal fue: Investigar y poner en valor el potencial forestal, productivo e industrial de la especie. Los resultados más destacados son: i) La Colección Nacional de Germoplasma de guarango, con 42 muestras, cuyos duplicados están plantados en dos sitios de la Sierra: Tunshi, Chimborazo y Puellaro, Pichincha, en forma de Jardín Clonal y el duplicado de semillas está depositado en el Banco Nacional de Germoplasma, a cargo del INIAP; ii) El desarrollo de técnicas de manejo y silvicultura para la especie, que incluye gestión de vivero, y manejo de plantaciones, que se han consolidado en varias publicaciones, se resalta la más importante: (Nieto C., y G. Hidrobo, 2010). La cadena Agro productiva del guarango. (Caesalpinea spinosa Kuntze), elementos que resaltan su competitividad. Fundación Desde el Surco, SENESCYT. Quito, Ecuador, 45 P; iii) La formación de la Asociación Nacional de Plantadores de Guarango, ASOPROG, filial Pichincha, que se encuentra legalmente constituida como empresa asociativa y funcionando; iv) La realización de varios estudios de factibilidad para la gestión de la cadena agroindustrial y de mercado para derivados de la especie. Entre las principales conclusiones se resaltan las siguientes: El guarango es una especie FPM típica, ideal para programas de industria forestal. El guarango tiene potencial para programas de desarrollo rural, mediante la formación de emprendimientos comunitarios, que manejen la cadena agroindustrial de la especie y fomenten la participación social. El guarango es una planta ideal para programas de desarrollo agroforestal y para reforestar áreas degradadas de BST. Palabras clave: Taninos, Gomas, Forestación productiva, Industria forestal, Agroforestería, Participación social PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 CONDICIONES DE SITIO Y CRECIMIENTO DE Centrolobium ochroxylum ESPECIE NATIVA CON POTENCIAL AGROSILVOPASTORIL DEL LITORAL ECUATORIANO? Álvaro Cañadas1, Diana Rade1, Carlos Zambrano2, Carlos Molina2 y Luis Arce2 1 Universidad Técnica del Norte, Facultad de Ingeniería en Ciencias Agropecuarias y Ambientales, Av. 17 de Julio 5-21 y José Córdova, Ibarra, Ecuador Teléfono: 00593-6-2997800,www.utn.edu.ec 2 Instituto Autónomo de Investigaciones Agropecuarias (INIAP). Estación Experimental Tropical Pichilingue. Km 5 vía Quevedo - El Empalme, cantón Mocache, Provincia Los Ríos, Ecuador. [email protected] La selección de especies arbóreas nativas para el establecimiento de plantaciones agroforestales o con fines múltiples es un continuo desafío y oportunidad para la silvicultura tropical. No obstante de esto, existe una falta de información científica sobre las especies forestales nativas, sus requerimientos silviculturales y aptitudes ecológicas, la cuales son un gran limitante para el uso amplificado de las especies nativas en el Ecuador. La presente investigación muestran los resultados obtenidos para determinar las condiciones de crecimiento y producción del Amarillo Centrolobium ochroxylum para el litoral Ecuatoriano. Los datos provienen de parcelas temporales e intervalos de un total de 1300 parcelas establecidas de 9,348.00 sitios de reforestación, con toma de datos consecutivos entre el 2006-2008-2009 y las edades de los rodales fluctúan entre 1 a 30 años de edad. Ecuaciones de volumen, curvas de índice de sitio, modelos de producción fueron desarrolladas. Así como también, se determinó el incremento medio y corriente del Amarillo. Las curvas de índice de sitio fueron utilizadas para clasificar las condiciones de crecimiento en clases I, II, III y IV en orden decreciente de productividad. Con las funciones de producción desarrolladas para el Amarillo se pudo establecer el punto de corta de esta especie, con la finalidad de determinar el óptimo de rotación de esta especie. De tal manera, que se puedan determinar correctamente el cálculo del análisis de beneficio/costo y determinar la viabilidad económica de esta especie, para fines productivos o de conservación. Palabras Clave: Índice de Sitio, Incremento Medio Anual, Incremento Corriente, Modelos de Producción, Modelos no Lineales PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 LISTADO DE AUTORES DE LOS ARTÍCULOS PROVISTOS EN LA PRESENTE MEMORIA En orden alfabético: Autor Institución Correo electrónico Aliana Piñeiro Fundación Runa [email protected] Álvaro Cañadas Universidad Técnica del Norte [email protected] Andrea Pinos Pontificia Universidad Católica del Ecuador [email protected] Andrés Recalde Redrován Pontificia Universidad Católica del Ecuador [email protected] Angélica Cárdenas Instituto Nacional Autónomo de Investigaciones Agropecuarias, Programa Nacional de Forestería [email protected] Antonio Vera Instituto Nacional Autónomo de Investigaciones Agropecuarias, Programa Nacional de Forestería [email protected] Carlos Nieto Pontifica Universidad Católica del Ecuador [email protected] David Romero Jardín Botánico Milpe [email protected] Doris Chalampuente Centro Agronómico Tropical de Investigación y Enseñanza - CATIE [email protected] Eduardo Colina Universidad Técnica de Babahoyo [email protected] Eduardo Cueva Naturaleza y Cultura [email protected] PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Internacional Fabián Salas Gobierno Autónomo Descentralizado de Chimborazo [email protected] Felipe Bravo Oviedo Instituto Universitario de Investigación Gestión Forestal Sostenible, Universidad de Valladolid INIA [email protected] Franklin Cargua Universidad Nacional de Chimborazo [email protected] Franklin Sigcha Instituto Universitario de Investigación Gestión Forestal Sostenible, Universidad de Valladolid INIA [email protected] Galo Fernando Pillajo Ministerio de Agricultura, Ganadería, Acuacultura y Pesca, Subsecretaría de Ganadería [email protected] Gloria Cristina Luna Universidad de Nariño [email protected] Grabriela Jami Instituto Nacional Autónomo de Investigaciones Agropecuarias, Programa Nacional de Forestería [email protected] Ingrid Martínez Instituto de Investigaciones [email protected] Agropecuarias de Chile Isabel Aulló Instituto Nacional Autónomo de Investigaciones Agropecuarias, Programa Nacional de Forestería [email protected] PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Janice Jácome Pontificia Universidad Católica del Ecuador [email protected] Jenny Ordóñez Centro Internacional de Investigación en Agroforestería (ICRAF) [email protected] Jesús María Bastida Escuela Superior Politécnica de Chimborazo, PrometeoSENESCYT [email protected] Jimmy Cevallos Zambrano Universidad Laica Eloy Alfaro de Manabí [email protected] Jonathan Cornelius Centro Internacional de Investigación Agroforestal [email protected] Jorge Caranqui Herbario de la Escuela Superior Politécnica de Chimborazo [email protected] Jorge Luis Cué García Universidad Técnica del Norte [email protected] José Ochoa Instituto Nacional Autónomo de Investigaciones Agropecuarias, Departamento de Protección Vegetal [email protected] José Riofrío Instituto Universitario de Investigación en Gestión Forestal Sostenible, Universidad de ValladolidINIA [email protected] Juan Chucuri Instituto Nacional Autónomo de Investigaciones Agropecuarias, Departamento Nacional de [email protected] PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Recursos Fitogenéticos Juan Dueñas Grupo Runa [email protected] Leonardo Gallo Grupo de Genética Ecología y Mejoramiento Forestal, INTA Bariloche, Argentina [email protected] Lucrecia Maldonado Instituto Nacional Autónomo de Investigaciones Agropecuarias [email protected] María Mercedes Pavón Instituto Nacional Autónomo de Investigaciones Agropecuarias, Programa Nacional de Forestería [email protected] Mario Añazco Universidad Técnica del Norte [email protected] Mauricio Cáceres Instituto Espacial Ecuatoriano [email protected] Miren del Río Departamento de Silvicultura y Gestión de Sistemas Forestales. CIFOR-INIA [email protected] Nancy Criollo Instituto Nacional Autónomo de Investigaciones Agropecuarias, Programa Nacional de Forestería [email protected] Paulo Barrera Instituto Nacional Autónomo de Investigaciones Agropecuarias, Programa Nacional de Forestería [email protected] PRIMER ENCUENTRO DE BOSQUES, RECURSOS GENÉTICOS FORESTALES Y AGROFORESTERÍA Centro de Convenciones Eugenio Espejo, Quito: 11 al 23 de noviembre de 2013 Pedro Ramírez GIZ-Programa GESOREN [email protected] Rodrigo Sierra GEO-15 [email protected] Rommel Montúfar Pontificia Universidad Católica del Ecuador [email protected] Roy Vera Vélez Instituto Universitario de [email protected] Investigación Gestión Forestal Sostenible, Universidad de Valladolid – INIA. Programa Nacional de Forestería INIAP Sebastián Escobar Pontificia Universidad Católica del Ecuador [email protected] Tomislav Sapic Faculty of Natural Resources Management, Lakehead University, Canada. [email protected]
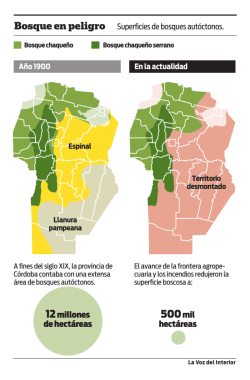

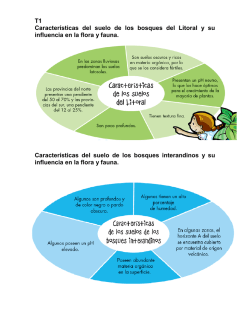
© Copyright 2026